一株茶轮斑病病原菌的分离鉴定及致病力
2021-06-09卢声洁赵兴丽罗林丽程宇豪张金峰周玉锋
卢声洁,赵兴丽,罗林丽,程宇豪,张金峰,李 帅,周玉锋*
1. 贵州大学 茶学院,贵州 贵阳 550025;2. 贵州省农业科学院 生物技术研究所,贵州 贵阳 550006;3. 贵州省农业科学院 茶叶研究所,贵州 贵阳 550006
茶树是我国传统经济作物,其经济价值主要集中于叶片。茶轮斑病(Tea grey blight)是茶树栽培中最为常见的一类叶部病害,可危害不同时期的茶树叶片;初期在叶片上形成灰色或褐色病斑,后期可导致叶片枯死脱落,严重时甚至引发整株茶树死亡,给茶叶生产带来较大经济损失[1-3]。前人研究发现,茶轮斑病主要由类拟盘多毛孢(Pesta lotiopsis-like)即原拟盘多毛孢属Pestalotiopsis Steyaert真菌侵染引起[4-5];目前,已报道的病原长刚毛拟盘多毛孢(Pestalotiopsis longiseta)、茶拟盘多毛孢(P. theae)、山茶拟盘多毛孢(P. camelliae)、棒形新拟盘多毛孢(Neopestalotiopsis clavispora)、山龙眼新拟盘多毛孢(N. protearum)、茶假拟 盘 多 毛 孢(Pseudopestalotiopsis camelliaesinensis)等16余种[6-9]。由于引起茶轮斑病的拟盘多毛孢属真菌种类较多, 且不同种间生物学特性存在差异,给茶轮斑病病害的防治带来困难。因此,明确该病致病菌种类和致病力是指导茶轮斑病防治的基础。
在早期茶树病害研究中,多以采用传统形态学特征结合致病性的方法对其病原进行鉴别和区分[10-11],但由于类拟盘多毛孢属真菌种间关系较为复杂,不同培养条件下菌落形态、分生孢子大小及颜色等均可能存在一定差异,给病原准确分类造成一定难度[12]。近年来,随着分子分类技术在物种划分中的广泛应用, 形态学特征结合致病性和多基因序列分析成为茶轮斑病病原鉴定的主要方法[13-15]。本研究为明确发生在贵州省湄潭县茶叶所资源圃中的茶轮斑病致病病原,采用单孢分离法对致病菌进行分离纯化,结合形态学观察、致病力测定以及分子生物学方法综合鉴定病原种类,以期为该地区病害防治提供理论依据。
1 材料与方法
1.1 材料
植物材料:健康茶树叶片采自贵州省农科院茶叶所资源圃,品种为福鼎大白茶。罹病叶片采自贵州省湄潭县茶叶所资源圃,品种为福鼎大白茶。
培养基:PDA和WA培养基的配制参照文献[16]。
1.2 方法
1.2.1 病原病菌分离纯化
采用单孢分离法对田间采回的罹病叶片进行病原分离。体视镜视野下找到病叶上的子实体,将注射器针头灼烧2 ~ 3次后,挑取子实体放入盛有少量灭菌水的陶瓷点滴板中充分破碎,制成分生孢子悬浮液,用灭菌胶头滴管将其吹打均匀后吸取一定量的悬浮液滴于皿底划有网格线并加有链霉素(50 µg/mL)的2%的水琼脂(WA)培养基平板上,室温静置培养12 h后挑取萌发的单个孢子接种到新鲜的PDA平板,于25℃条件下黑暗培养5 d。得到纯化的菌株后,转管保存。
1.2.2 病原致病性测试
采用针刺法对离体茶树叶片进行致病性测试。选取新鲜且大小一致无病斑无伤口的茶树叶片,经75%酒精表面消毒后置于铺有无菌滤纸的培养皿内,用无菌注射器分别对茶树叶片正面和背面进行针刺处理,无菌接种针挑取5 mm的病原菌菌饼贴于伤口处,以接种无菌PDA为对照,恒温保湿培养,每天观察并记录其发病情况,待病斑上产生子实体后再次挑取子实体分离致病菌。
1.2.3 病原鉴定
形态学观察:将分离所得菌株接种于PDA平板上培养 5 ~ 7 d,观察其培养性状;待产孢后挑取少量菌丝及分生孢子于镜下拍照观察,描述菌落形态、繁殖体孢子等形态特征。
分子鉴定:采用Ezup柱式真菌基因组DNA 抽提试剂盒提取病原菌基因组DNA,并以所提基因组DNA为模板,利用引物ITS5/ITS4(5′-GGAAGTAAAAGTCGTAACAAG-3′/5′-TCCTCCGCTTATTGATATGC-3′ )、Bt2a/Bt2b(5′-GGTAACCAAATCGGTGCTGCTTTC-3′/5′-ACCCTCAGTGTAGTGACCCTTGGC-3′)、EF1-526F/EF1-1567R(5′-GTCGTYGTYA TYGGHCAYGT-3′/5′-ACHGTRCCRATA CCACCRATCTT-3′)对菌株的基因组DNA进行PCR 扩增[17]。反应体系(25 μL):2×Taq PCR Master Mix 12.5 μL,dd H2O 9.5 ul,上游引物、下游引物和DNA模板各1 μL。ITS扩增程序为:95℃预变性3 min,95℃变性30 s,55℃退火45 s,72℃延伸60 s,35个循环,最后72℃延伸10 min;β-tubulin扩增程序为:95℃预变性4 min,94℃变性48 s,57℃退火50 s,72℃延伸60 s,35个循环,72℃延伸5 min;tef1扩增程序为:94℃预变性5 min,94℃变性18s, 54℃退火33 s,72℃延伸30 s,35个循环,最后72℃延伸7 min。PCR产物经1%琼脂糖凝胶电泳检测质量后,送生工生物工程 (上海) 有限公司测序。将测序结果与从 GenBank中下载公认菌株或模式菌株对应的基因序列用BioEdit进行比对和校正,利用 MAGA 6.0软件将三条基因序列按ITS、β-tubulin和tef1的顺序连接起来,比对拼接后的序列采用邻接法(N-J)构建系统进化树,确定该菌与同属菌株间的亲缘 关系。
2 结果与分析
2.1 病原菌分离与纯化结果
从具典型轮纹病斑的茶树病叶上分离获得一株茶轮斑病菌ZYP04-5,经3次续代培养后,得到纯菌落,斜面和水保存备用。
2.2 病原菌的致病性
对茶树叶片正反面进行针刺处理,接种ZYP04-5菌株4 d后可见叶片发生明显棕褐色病变,且有病菌菌丝穿透针刺部位附着在叶片上(图1B ~ 图1C)。叶片正面针刺处理发病程度快于背面针刺处理,而对照(图1A)和无刺伤处理(未体现)均无明显现象。当培养时间延长至9 d后,正面针刺处理病斑扩大至叶缘,而背面针刺处理病斑扩散不明显,此时病斑由初期的棕褐色转为中部黄褐、边缘黑褐色轮纹状并有少量棕褐色子实体排列成轮纹状附着在病斑上(图1D ~ 图1E)。挑取病斑子实体进行镜检和单孢分离培养,可获得与原接种菌株ZYP04-5形态特征基本一致的分生孢子和菌落。

图1 叶片正反面接种形成的病斑及显微特征Figure 1 Morphological characteristics of disease spots formed by inoculation on both sides of leaves
2.3 病原菌形态学鉴定结果
在PDA上,25 ℃条件下黑暗培养7 d后菌株ZYP04-5菌落可铺满直径为90 mm的培养皿,菌落白色至乳白色,背面白色,棉絮状,边缘整齐,具轮纹,轮纹由内向外呈放射状(图2A ~ 图2B);在菌落中央可形成半流动状黑色黏液团,接种针挑取一点制成水玻片于显微镜下观察,可发现产孢细胞在产孢初期时较长,直立状(图2C);中期分生孢子膨大,烧瓶状或啤酒瓶状(图2D ~ 图2F),淡色或无色;后期分生孢子纺锤形,大小约为(18.20 ~ 29.96)μm×(4.23 ~ 6.73)μm,具有 4隔膜 5细胞,分隔处向内稍微缢缩,中间横隔明显;分生孢子两端细胞呈锥形,无色;中间3细胞异色,长约为 12.13 ~ 17.63 μm,呈瓮状至近圆柱形;基部向上第二细胞浅褐色或橄榄绿色,三、四细胞深褐色或深橄榄绿色;顶端具2 ~ 3根附属丝,长约为 11.1 ~ 22.7 μm;基部附属丝 1 根,长约为 1.92 ~ 6.37 μm。根据形态学特征,初步将该菌鉴定为新拟盘多毛孢Neopestalotiopsis spp.属真菌。
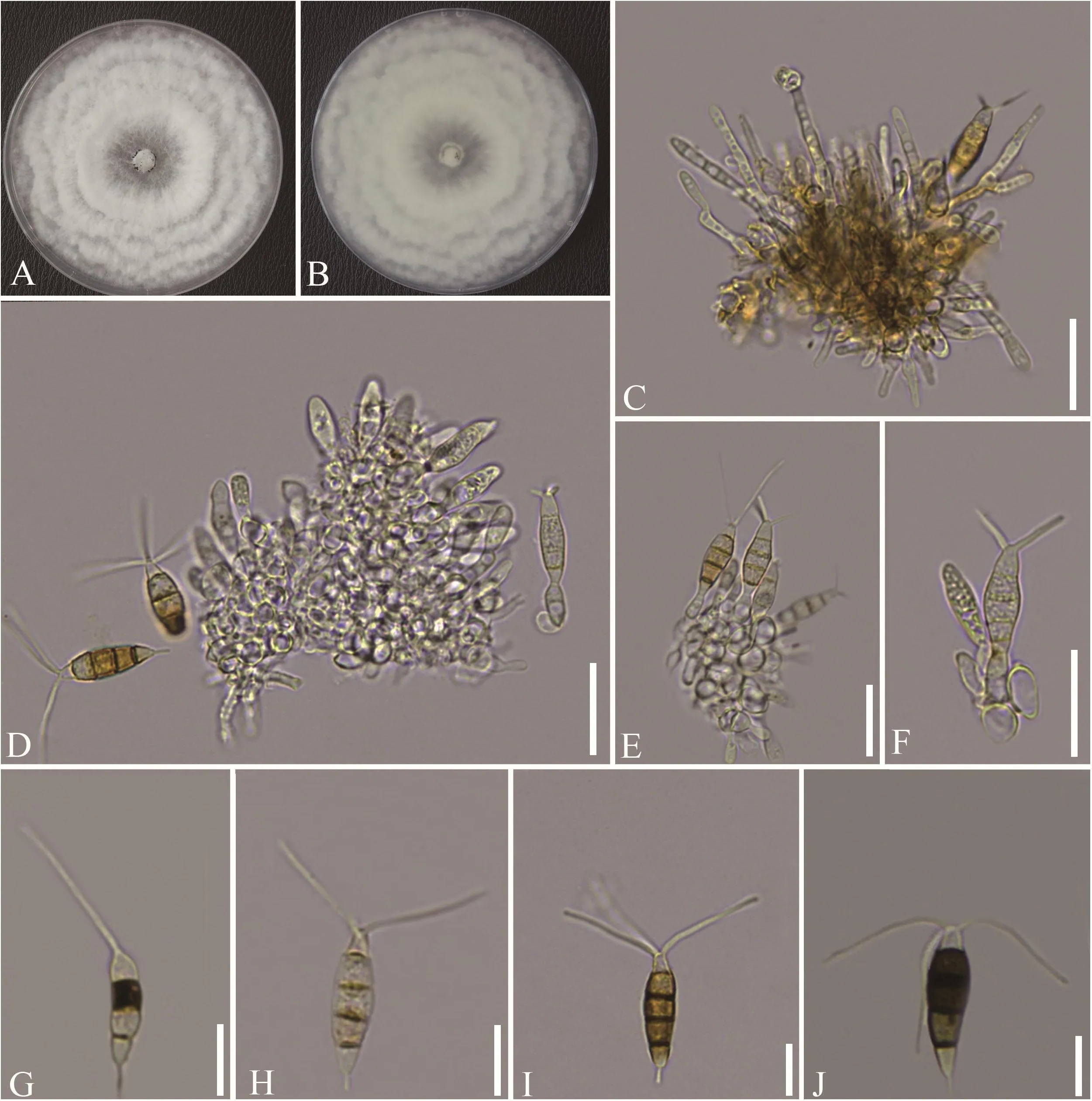
图2 菌株ZYP04-5的形态特征Figure 2 Morphological characteristics of strain ZYP04-5
2.4 分子鉴定结果
PCR分别扩增基因组DNA的ITS、β-tubulin和 tef1片段,大小分别为 580 bp、480 bp和1010 bp,经BLAST比对分析后选取并下载序列相似性高的菌株为内群,P. trachicarpicola为外群(表1);用BioEdit进行多位点序列比对并对其进行手动校正,MAGA 6.0软件拼接相应序列片段后构建ZYP04-5的邻接树(图3)。结果表明,菌株ZYP04-5以86%的支持率与N.ellipsospora聚为一枝,结合该菌株形态学特征分析,确定菌株ZYP04-5为卵圆新拟盘多毛孢(Neopestalotiopsis ellipsospora)。
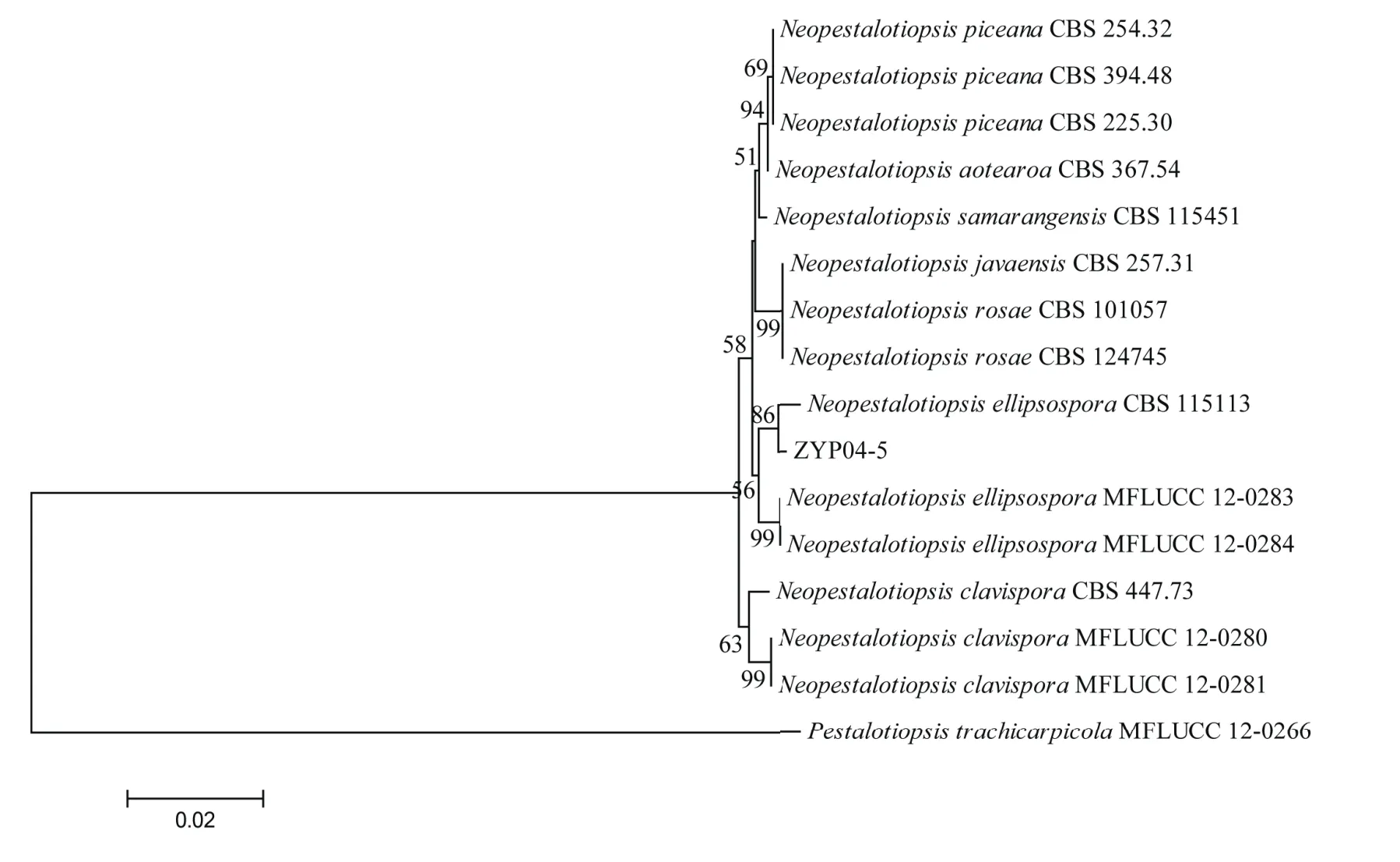
图3 基于ITS、β-tubulin和tef1构建邻接树Figure 3 Adjacency tree construction based on multi-locus sequences of ITS、β-tubulin 和 tef1
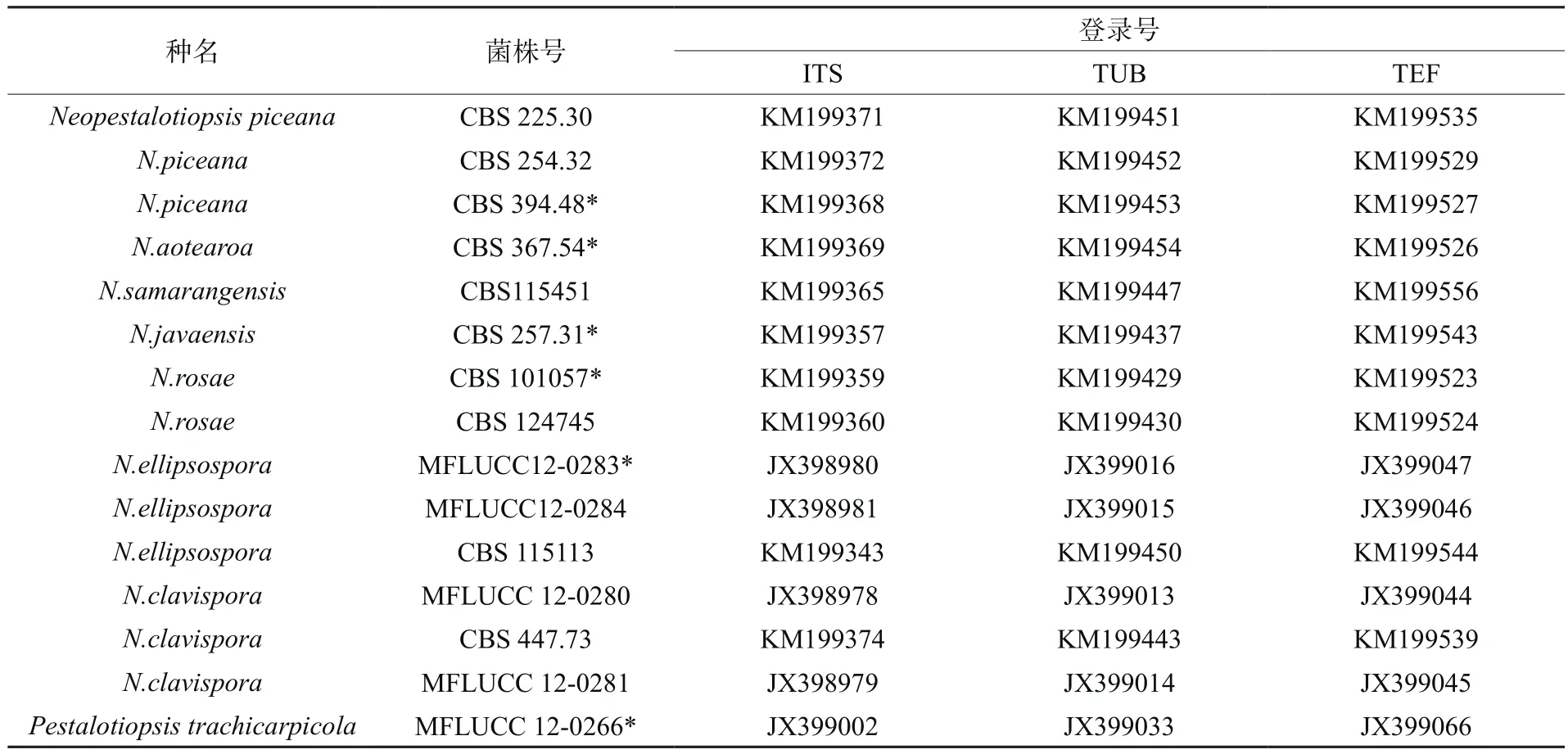
表1 所用的菌株情况Table 1 Strains used in this study
3 讨论
类拟盘多毛孢(Pestalotiopsis-like)属真菌为弱寄生性真菌,据报道该属真菌可侵染50多科植物发病[12]。传统上对该属真菌进行分类主要以依据分生孢子颜色和大小、附属丝长度和数量等来进行区分。2014年,Maharachchikumbura结合ITS、β-tubulin和tef三基因加合分析结果,将拟盘多毛孢属真菌按分生孢子颜色同色、同色且颜色较深和异色将其分成3个主要的进化枝,即Pestalotiopsis属、Pseudopestalotiopsis属和Neopestalotiopsis属[18]。本文报道的菌株分生孢子形态即属于第三个类群,形态学特征结合多基因联合分析将分离自贵州省遵义市湄潭县茶叶所资源圃中的茶轮斑病病原鉴定为卵圆新拟盘多毛孢(N.ellipsospora)。
国内外关于N.ellipsospora引起植物病害的报道较少, 仅在茶树、五加叶、莎叶兰和甘薯等一些经济作物上有相关报道[19-21],而关于N.ellipsospora在茶树上的研究,Wang等[7]从采自浙江和四川的茶轮斑病病叶中分离到6株N.ellipsospora,但未对该菌的致病性进行分析;Wang等[22]从采自安徽庐江的茶轮斑病病叶中分离到2株N.ellipsospora,经柯赫氏验证,该菌对10个茶树品种的叶片均不致病,因此推测其可能是茶树内生菌或腐生菌。本研究为确定从茶轮斑病病叶上分离的菌株是否能侵染茶树叶片,采用柯赫氏法则探讨了菌株ZYP04-5对茶树的侵染情况,结果表明该菌株能够侵染茶树引起茶轮斑病,研究结果可为茶轮斑病的防治提供一定的理论依据。
