基于宏基因组学技术不同地区蛋源表面微生物的比较
2021-06-04温佳奇王秀娟宋雨奇代伟长王玉华
李 侠,温佳奇,王秀娟,宋雨奇,代伟长,王玉华
(吉林农业大学食品科学与工程学院,吉林 春 130118)
我国是生产和消费禽蛋大国,禽蛋和蛋制品的质量与安全也备受消费者关注[1]。蛋源在母禽生殖道及生产环境中会受到污染,表面带有大量微生物,未经处理的禽蛋在打蛋过程中微生物会感染到蛋液[2-4];液蛋制品加工时,分离、过滤及均质过程中蛋液也会因加工器械等未消毒完全而被污染。虽然灌装后的液蛋制品会进行灭菌,但前期大量微生物的污染会导致灭菌难度大,灭菌效果差,影响液蛋制品的质量及保质期。因此,微生物污染源的控制非常重要[5],在可能产生污染的过程里,蛋源的微生物污染数量最多,污染最严重,对蛋源微生物来源的控制成为控制液蛋制品微生物污染源重中之重。蛋源表面的微生物种类繁多,常见的微生物有大肠杆菌属、假单胞菌属、不动杆菌属、变形杆菌属、产气单胞菌属、葡萄球菌属、沙门氏菌属、产碱杆菌属、副大肠杆菌属等[6-8]。据相关材料报道,输卵管带菌的家禽所产的蛋,70%是带菌蛋,所带细菌如鸡白痢沙门氏菌、鸡副伤寒沙门氏菌及金黄色葡萄球菌等[9-10]。禽蛋腐败的主要原因是微生物的存在,主要有非致病性细菌和霉菌。分解蛋白质的微生物主要是芽孢杆菌属、假单胞菌属、变形杆菌属、梭菌属、葡萄球菌属、无色杆菌属、产碱杆菌属、黄杆菌属和肠杆菌属等[11-12];分解糖的微生物有大肠杆菌属、枯草杆菌属和丁酸梭状芽孢杆菌属等[13];分解脂肪的微生物主要有假单胞菌属、沙门氏菌属等[14]。此外,还可能存在一些不可培养的微生物,特别是致病菌的存在,如霍乱弧菌和大肠埃希氏虽均具有生命活性,但不能培养[15-17],而这些未被培养的种类却代表了巨大的多样性,此类微生物可能存在潜在危害。因此,对蛋源表面微生物多样性的探究对后续消毒处理方法的选择具有重要意义,有助于液蛋制品产业的发展。
鸡蛋表面微生物种类繁多,利用传统的微生物培养法虽然能检测出常见食品致病菌的数量,但并不能将所有菌相完全高效、准确地检测出来,并且传统方法耗时耗力,将高通量测序宏基因组技术应用于蛋源表面微生物多样性的检测,从宏观角度准确高效地检测出微生物种类,为后期对蛋源前处理消毒方法的优化研究奠定基础,同时也拓宽了宏基因组学技术在食品领域的探索。本实验采用高通量测序的宏基因组学技术,分析吉林、安徽、北京和河北4 个不同地区蛋源表面微生物多样性,确定蛋源表面微生物菌相分布,并分析不同蛋源之间的差异性,为蛋源消毒前处理方法的优化提供一定的参考。
1 材料与方法
1.1 材料与试剂
当日产新鲜鸡蛋,分别来自吉林、北京、河北、安徽地区的大规模蛋鸡养殖企业(存栏量达300万 羽以上)。
营养琼脂培养基、结晶紫中性红胆盐琼脂培养基、缓冲蛋白胨水、亚硫酸铋琼脂培养基 北京奥博星生物技术有限公司;DNA试剂盒 北京全式金生物技术有限公司;其他试剂均为国产分析纯。
1.2 仪器与设备
AUM220电子分析天平 日本岛津公司;MCV-131超净工作台 上海博讯实业有限公司;AHWY-200B超低温冰箱 日本三洋公司;YX280立式压力蒸汽灭菌器、YX280立式压力蒸汽灭菌器 上海申安医疗有限公司;Infinile M200多功能酶标仪 瑞士TECAN公司;DHG-9146电热恒温培养箱 上海精宏实验设备有限公司。
1.3 方法
1.3.1 样品的采集
在同一月份,分别从吉林、北京、河北、安徽4 个地区的鸡场进行样品采集,随机采集每个鸡场当日产出的新鲜鸡蛋,用灭菌生理盐水棉拭子均匀擦拭蛋壳表面,每组样品50 枚,每个地区采6 组平行样品。取样后,-80 ℃冷冻保存,用于后续蛋源蛋壳表面微生物多样性分析。
1.3.2 细菌菌落总数测定
将擦拭蛋源表面的棉拭子充分溶于无菌生理盐水、混匀,获得样品菌液,然后进行稀释,分别取稀释倍数为10-3、10-4、10-5的样品,按照GB 4789.2—2016《食品微生物菌落总数的测定》进行蛋源表面菌落总数的测定。
1.3.3 DNA的提取
将擦拭蛋源表面的棉拭子充分溶于灭菌生理盐水,采用EasyPureGenomic DNA试剂盒进行DNA提取,严格按照试剂盒说明书进行操作。
1.3.4 微生物多样性分析
通过蛋源表面微生物16S rRNA基因的高度可变区V3和V4进行测序,基于测序结果进行生物多样性分析。实验流程为:提取的DNA→靶标片段PCR扩增→扩增产物回收和纯化→扩增产物荧光定量→测序文库制备→高通量测序,实验测序工作由上海派森诺生物科技股份有限公司完成。通过操作分类单元(operational taxonomic unit,OTU)对获得的序列进行分类,并且通常使用97%的序列相似度作为OTU分区阈值,其等于分类中的物种水平。根据不同样品中OTU的丰度分布,评价每个样品的多样性水平,使用稀疏曲线反映测序深度是否达到标准。分析不同分类水平的各地区微生物的具体组成,对吉林、安徽、北京和河北4 个地区的蛋源表面微生物菌群进行α多样性、群落结构和β多样性分析比较。
1.4 数据分析
实验数据利用Origin 2017、GraphPad Prism 7.0.软件和Excel 2007对数据进行整理、计算、统计与分析。P<0.05为差异显著、P<0.01为差异极显著、P<0.001为差异高度显著,具有统计学意义。
2 结果与分析
2.1 不同地区蛋源表面微生物OTU分布
Venn图可用于统计不同样本中共有和独有的OTU数目,本研究选用相似度为97%的OTU,结果如图1所示。吉林地区蛋源表面微生物OTU总的数目为2 341,其中独有的OTU数目为1 285,河北地区蛋源表面微生物OTU总的数目为1 216,其中独有的OTU数目为328,北京地区蛋源表面微生物OTU总的数目为1 708,其中独有的OTU数目为295,安徽地区蛋源表面微生物OTU总的数目为1 301,其中独有的OTU数目为135。4 个地区共有的OTU数目为218,吉林和河北地区共有的OTU数目为488,吉林和北京地区共有的OTU数目为765,吉林和安徽地区共有的OTU数目为574,河北和北京地区共有的OTU数目为675,河北和安徽地区共有的OTU数目为540,北京和安徽地区共有的OTU数目为957。由OTU数目分布Venn图可以初步得出,吉林地区微生物丰富度最高,北京和安徽地区微生物最相似。
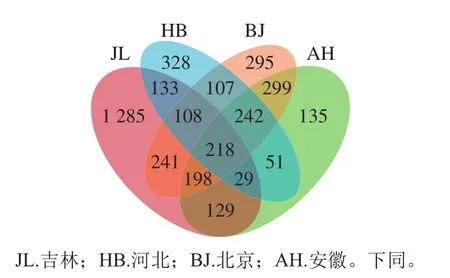
图1 不同地区蛋源表面微生物OTU的Venn图Fig.1 Venn diagram showing unique and shared microbial OTUs among eggs from different regions
2.2 不同地区蛋源表面微生物α多样性分析
α多样性即每个样本群落的多样性,可用Shannon指数曲线和α多样性指数等进行评价。
2.2.1 不同地区蛋源表面微生物Shannon指数曲线
Shannon指数曲线是反映样品中微生物多样性的指数,利用各样品的测序量在不同测序深度时的微生物多样性指数构建曲线,以此反映各样本在不同测序数量时的微生物多样性[18]。当曲线趋向平坦时,说明测序数据量足够大,可以反映样品中绝大多数的微生物信息。可以用来判断测序的有效性[19]。本实验选用Shannon指数曲线对测序有效性进行分析。
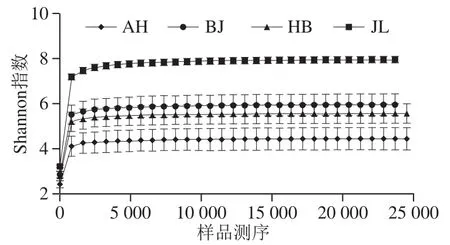
图2 不同地区蛋源表面微生物Shannon指数曲线Fig.2 Shannon index curves showing microbial community diversity on eggs from different regions
如图2所示,横坐标表示每个样品中随机抽取序列的总数;纵坐标表示微生物多样性的Shannon指数。曲线的长度反映了样品测序的数量。曲线的渐变程度反映了测序深度对观察样品多样性的影响,曲线趋于平坦,表明测序已趋于饱和,增加测序数据无法再找到更多的OTU。随着样品测序数量的不断增加,当其大于5 000时,曲线已趋于平缓,说明测序数据量足够大,足够反应样本中包含的微生物多样性。
2.2.2 不同地区蛋源表面微生物α多样性指数
对于微生物群落的α多样性可以通过Chao1指数、ACE指数、Shannon指数和Simpson指数反映。一般而言,Chao1指数或ACE指数越大,表明群落的物种种类越多、丰富度越高[20]。而Shannon多样性指数不仅反映群落的丰富度,还综合考虑了群落的均匀度。Shannon指数值越高,表明群落的多样性越高;Simpson指数与Shannon指数类似,是群落多样性的评价常用指数之一,Simpson指数值越高,说明群落的多样性越高[21]。
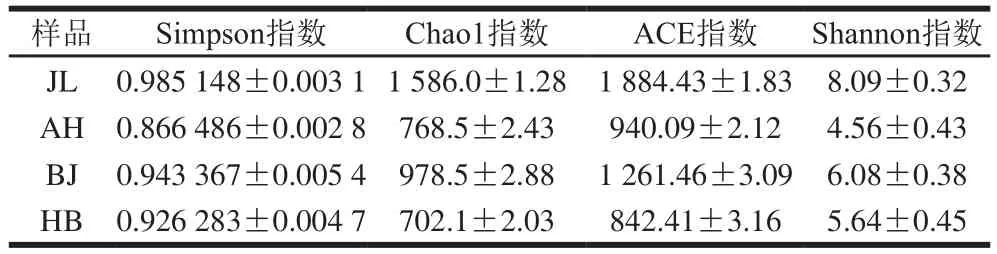
表1 不同地区蛋源表面菌群微生物多样性指数表Table 1 Microbial diversity indexes of surface microbial flora in eggs from different regions
由表1可知,4 个地区都具有较高的菌群多样性。其中Chao1指数和ACE指数表现为JL>BJ>AH>HE,说明在菌群丰富度上按照吉林、北京、安徽、河北的顺序依次降低;而Shannon指数和Simpson指数表现为JL>BJ>HE>AH,表明在群落多样性上按照吉林、北京、河北、安徽的顺序依次降低。由此可知,4 个地区相比,吉林地区蛋源表面微生物的群落丰富度及群落多样性均最高。
2.3 不同地区蛋源表面细菌群落结构分析
2.3.1 不同地区蛋源表面基于门水平的微生物分析
根据OTU划分和分类地位鉴定,获得了吉林、安徽、北京和河北在门、纲、目、科和属分类水平的具体微生物分布状况。在本研究中,选择门和属这2 个水平,对4 个地区的微生物进行分析。4 个地区蛋源基于门水平上的群落组成如图3所示。
吉林地区的蛋源样品细菌组成为放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和拟杆菌门(Bacteroidetes),相对丰度分别为49.12%、31.93%、8.41%及4.37%,还包括少量的异常球菌-栖热菌门(Deinococcus-Thermus)、蓝藻门(Cyanobacteria)以及绿弯菌门(Chloroflex);安徽地区的蛋源样品细菌组成为Proteobacteria、Firmicutes和Actinobacteria,相对丰度分别为77.03%、18.21%及2.90%,还包括少量的Cyanobacteria;北京地区的蛋源样品细菌组成为Proteobacteria、Firmicutes和Actinobacteria,相对丰度分别为57.93%、37.68%及2.93%,还包括少量的Cyanobacteria;河北地区的蛋源样品细菌组成为Proteobacteria、Firmicutes和Actinobacteria,相对丰度分别为84.28%、10.42%及3.12%,还包括少量的Cyanobacteria。安徽、北京和河北3 个地区蛋源菌群组成相似,优势菌门均为Proteobacteria,而在吉林地区Proteobacteria丰度仅为8.40%,变形菌门中包括很多种食品中常见的致病菌,如幽门螺杆菌、沙门氏菌和大肠杆菌等,吉林地区蛋源表面此类致病菌可能远低于其他3 个地区;而吉林地区的优势菌门Actinobacteria在其他3 个地区丰度较低, Chloroflex仅在吉林地区发现。因此,吉林地区与其他3 个地区在蛋源表面微生物菌群有很大差异,但由于其变形菌门丰度较低,含有常见致病菌的风险可能低于其他3 个地区,因此更适合作为蛋源生产产区。
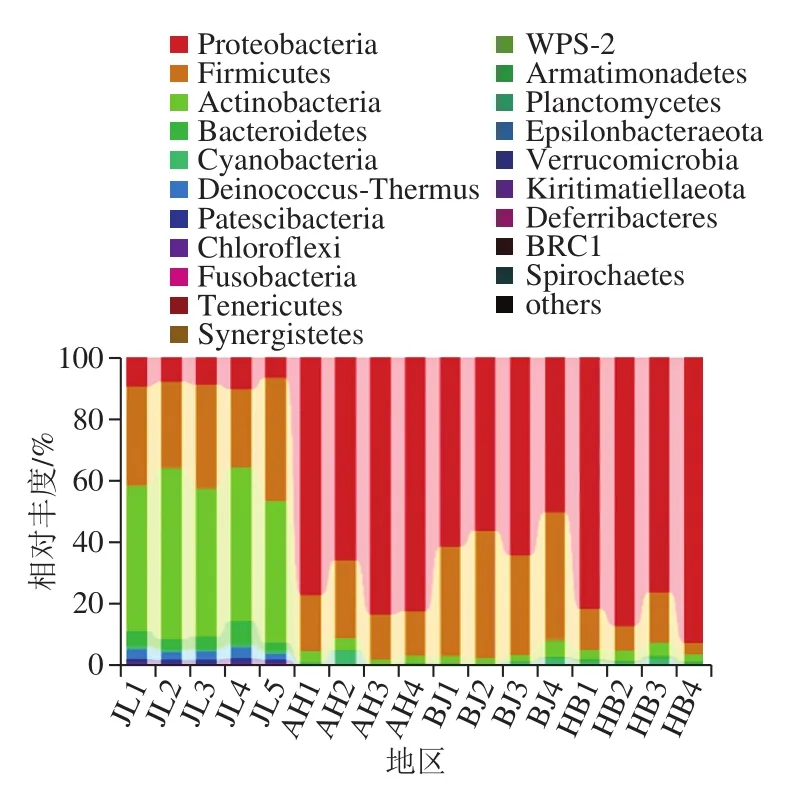
图3 基于门分类水平上的不同地区样品菌群分布图Fig.3 Distribution of microflora on eggs from different regions at the phylum level
2.3.2 不同地区蛋源表面基于属水平的微生物分析
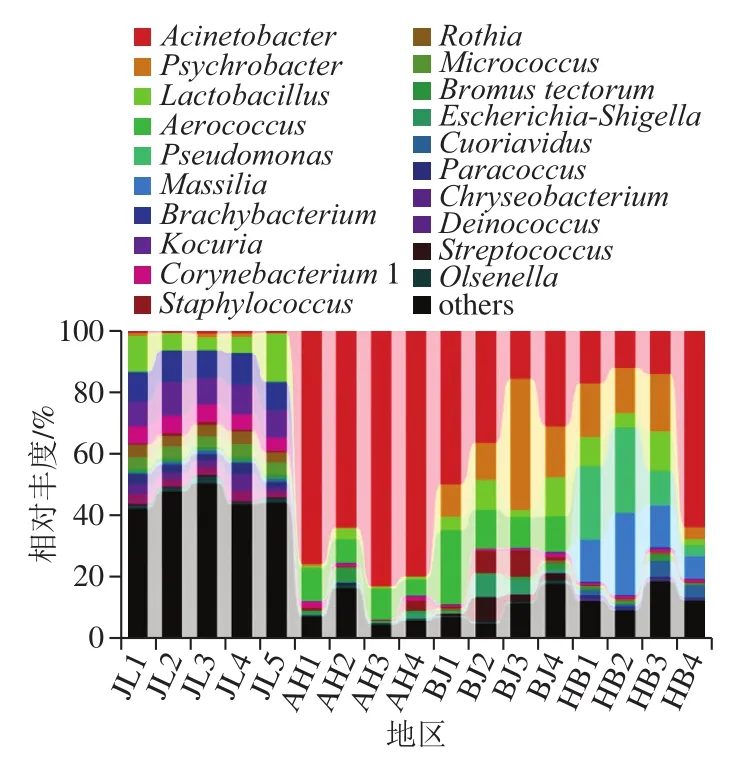
图4 基于属分类水平上的不同地区样品菌群分布图Fig.4 Distribution of microflora on eggs from different regions at the genus level
如图4所示,吉林地区的蛋源样品中丰度较高的菌属为考克氏菌属(Kocuria)占10.1%,短状杆菌属(Brachybacterium)占9.83%,榛状杆菌属(Corynebacterium1)占5.2%,uncultured占7.2%,微球菌属(Micrococcus)占3.95%,罗思氏菌属(Rothia)占3.80%,嗜冷杆菌属(Psychrobacter)占0.96%,不动杆菌属(Acinetobacter)占0.93%;安徽地区蛋源样品中丰度较高的菌属为Acinetobacter占75.52%,气球菌属(Aerococcus)占8.40%,乳酸菌属(Lactobacillus)占2.82%,Bromus tectorum占1.55%,葡萄球菌属(Staphylococcus)占1.20%;北京地区蛋源样品中丰度较高的菌属为Acinetobacter占26.42%,Psychrobacter占20.44%,Lactobacillus占15.12%,Aerococcus占14.36%,Staphylococcus占3.53%,uncultured占2.11%,埃希氏菌志贺氏菌属(Escherichia-Shigella)占3.15%;河北地区蛋源样品中丰度较高的菌属为Acinetobacter占26.70%,Psychrobacter占13.69%,Lactobacillus占7.32%,假单胞菌属(Pseudomonas)占16.28%,Massillia占15.39%。安徽、北京和河北3 个地区菌属分布较为相似,优势菌属均为Acinetobacter,而吉林地区Acinetobacter丰度仅为0.89%,Brachybacterium为吉林地区独有的丰度较高的菌属。由此表明安徽、北京和河北3 个地区菌属分布较为相似,优势菌属均为条件致病菌的Acinetobacter,而且存在其他致病菌属,即安徽地区蛋源样品Staphylococcus,北京地区蛋源样品Staphylococcus和Escherichia-Shigella,河北地区蛋源样品Pseudomonas。可见不同地区蛋源表面微生物属水平都存在一定差异性,相对丰度差异显著,北方地区与中部以南地区差异极显著(P<0.01),北方地区更适宜禽蛋生产。同时,为保障液蛋制品的安全性,蛋制品企业应该根据蛋源微生物的种类适当调整消毒处理方法。
2.3.3 不同地区蛋源表面微生物多样性聚类热图分析
根据不同样本的丰度分布或相似程度所组成的聚类分析图。其目的是通过聚类,可以区分高丰度和低丰度的分类单位,并且由颜色组成梯度反映样本之间的群落组成相似性。
如图5所示,4 个地区蛋源表面丰度前50 位的菌属聚类热图,红色代表相应样品中丰度较高的属,绿色代表丰度较低的属。在吉林地区丰度较高的29 个属在其他地区丰度较低;而河北地区丰度较高的属为Brevundimonas、Pseudomonas、Massillia、Alkanindiges、Cupriavidus、Sphingomonas和Ochrobactrum;安徽和北京地区蛋源属的分布最为相似,丰度较高的属为Acinetobacter、Jeoigalicoccus、Fackiamia、Lysinbacliius、Psychrobacter。
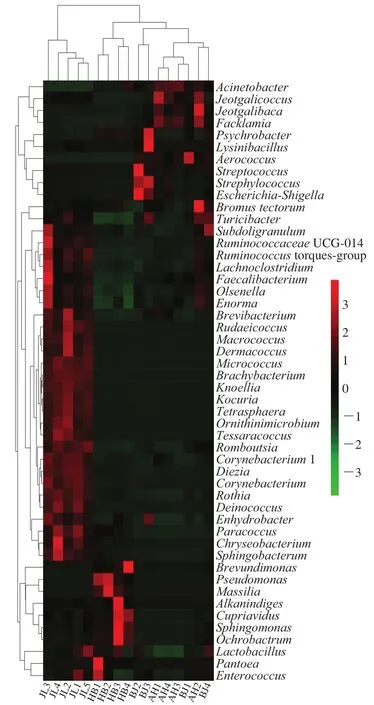
图5 不同地区蛋源表面微生物多样性聚类分析热图Fig.5 Heatmap of clustering analysis for microbial community diversity on eggs from different regions
2.4 不同地区蛋源表面微生物β多样性分析
β多样性分析的主要目的是评价不同样本之间群落结构的相似性。主要通过主成分分析(principal component analysis,PCA)、多维尺度分析(multidime-nsional scaling,MDS),对群落数据结构进行自然分解并通过对样本排序,从而观察样本之间的差异。
2.4.1 不同地区蛋源表面菌群PCA
菌群的PCA主要是通过线性变换将菌群OTU丰度矩阵组合,并通过线性变换将其投影到较低维空间坐标系中。从而达到降维的目的,简化数据结构,并显示样本的自然分布[22]。PCA可以从原始数据中提取样本之间最重要的差异,并根据这些差异在新的低维坐标系中对样本进行排序。使样本接近新坐标系可以最大程度地恢复样本之间的实际差异[23]。因此,通过分析PCA可以知道群落样本的主要分布特征,从而量化样本之间的差异和相似性。
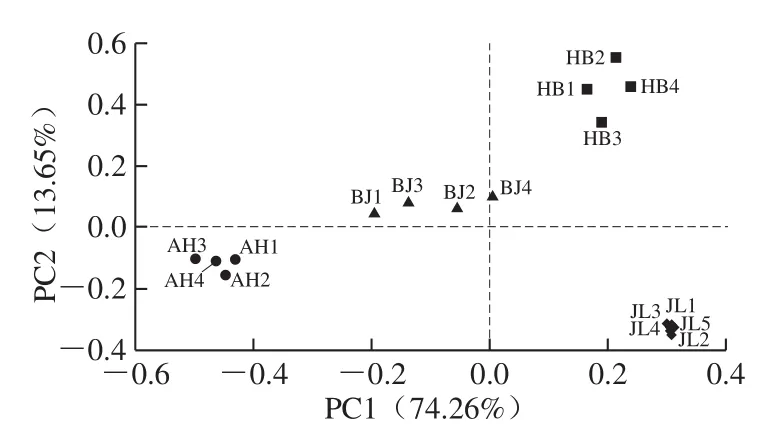
图6 不同地区蛋源表面菌群PCA二维排序图Fig.6 Two-dimensional sequencing diagram of PCA analysis for discrimination of microbial community composition on eggs from different regions
如图6所示,横坐标为PC1,其贡献率为74.26%。纵坐标为PC2,贡献率为13.65%。这2 个PC是样品微生物群落结构组成差异的主要因子。在PC1方向上,吉林和河北地区蛋源样品点与安徽地区样品点分布差异显著(P<0.05),样本点处于PC1水平的2 个极端;在PC2水平上,吉林地区样品点与河北地区样本点分布差异显著(P<0.05),吉林地区样本点分布于Y轴的下方,河北地区样本点分布于Y轴的上方。由此可知,吉林地区蛋源与河北地区蛋源菌群结构及微生物多样性差异极显著(P<0.01)。
2.4.2 基于UniFrac距离的NMDS非度量多维尺度分析
NMDS分析是基于样本距离矩阵的MDS分析方法。因此,在特定的距离尺度下描述了样本的分布特征。NMDS分析对样本距离进行排序,以使低维空间中的样本的顺序尽可能彼此接近。因此,NMDS分析不受样本距离值的影响,仅考虑彼此之间的大小关系[24]。
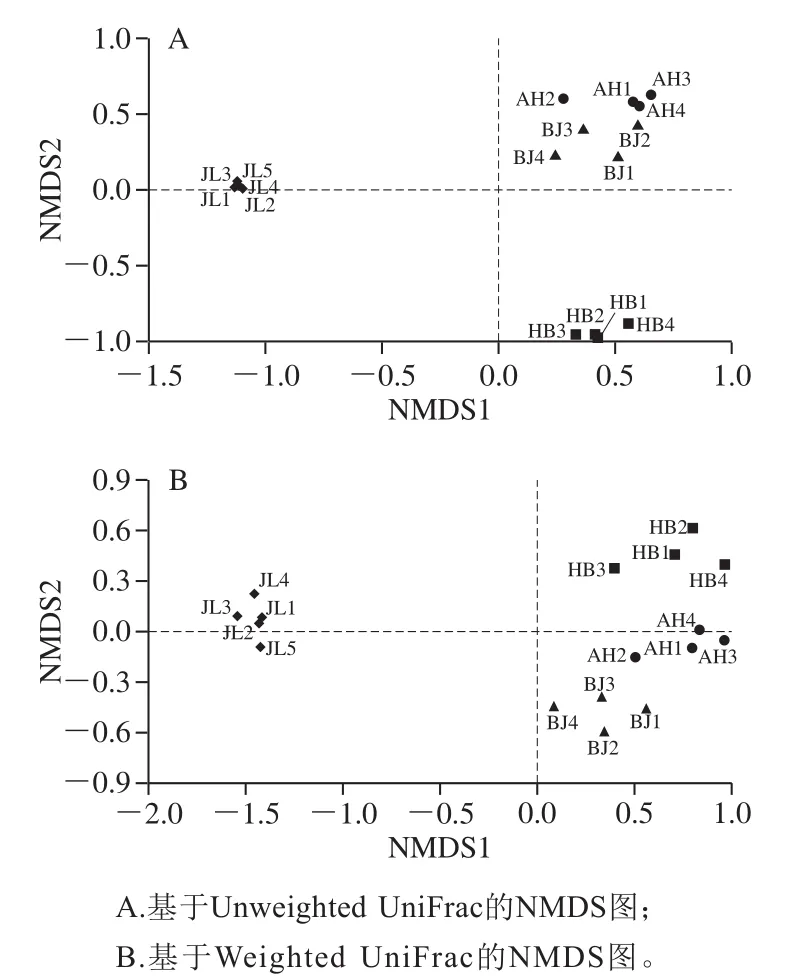
图7 不同地区蛋源表面菌群NMDS分析的二维排序图Fig.7 Two-dimensional sequencing diagram of NMDS analysis for discrimination of microbial community composition on eggs from different regions
由图7A可知,仅考虑OTU在样本中存在与否的情况下,安徽地区和北京地区样本之间的微生物群落结构相似度高,而吉林地区与河北地区样本差异性大,这与PCA中得到的结论吻合。而考虑由于群落丰度梯度导致差异的情况,如图7B可以看出,吉林地区样本与其他3 个样本之间有很大差异。因此,无论是否考虑OTU的丰度,吉林地区样本与其他地区差异都较大。
3 讨 论
蛋源微生物污染一直是蛋品领域值得研究的问题,采用高通量测序的宏基因组学技术能够一次并行对几十万到几百万条DNA分子进行序列测定,高效准确地测定出蛋源表面所有已知微生物菌群分布,它在分析微生物的群落结构时有着独特的优势,能够通过从环境样本中直接获取总DNA进行文库构建并测序[25-27],用16S rRNA基因的测序数据估计微生物群落的物种构成,更加真实地揭示原位环境中微生物群落的复杂性和多样性[28-29],对蛋源微生物有一个宏观的了解,可以详细地掌握蛋源表面所有基因库里已知菌属的情况;同时,本研究也发现蛋源表面微生物除了已知的菌属外,还存在一定量未知菌属,为今后更加深入研究蛋源表面微生物种类奠定良好基础,也为后续蛋源表面微生物多样性研究提供理论依据。本研究选择吉林、北京、河北、安徽4 个地区作为采样地区,是因为这4 个地区为国内主要蛋源产区,安徽以南温度过高,不适合大规模蛋鸡养殖,因此没有代表性。研究目的在于探究北部、中部和南部蛋源在菌群多样性上的差异性,从结果可以看出吉林地区的微生物多样性和丰富度与其他3 个地区差异性最大,优势菌门也不同,但值得关注的是,拥有较多食品致病菌的变形菌门在吉林地区丰度显著低于其他3 个地区,蛋液感染此类病源菌的风险也随之减少;而且不同地区样品微生物属水平差异更大,吉林地区的蛋源样品中丰度较高的菌属为Kocuria和Brachybacterium分别为10.1%和9.83%,目前还没有关于这2 个属存在致病菌的报道;安徽、北京和河北3 个地区菌属分布较为相似,优势菌属均为条件致病菌的Acinetobacter,另外还存在致病菌属。Acinetobacter是条件致病菌,当机体抵抗力降低时易引起机体感染,是引起医院内感染的重要机会致病菌之一。可引起呼吸道感染、败血症、脑膜炎、心内膜炎、伤口及皮肤感染、泌尿生殖道感染等。重症者可导致死亡。安徽地区蛋源样品致病菌属为Staphylococcus,北京地区蛋源样品致病菌属为Staphylococcus和Escherichia-Shigella,河北地区蛋源样品致病菌属为Pseudomonas。从相关文献报道可知,目前国内外对蛋源表面微生物的研究多采用传统的微生物培养法,集中在细菌的数量检测上,如Moyle等[30]对自由放养的2 个农场的鸡蛋表面细菌测定结果分别为3.84(lg(CFU/枚))和3.69(lg(CFU/枚)),分离培养得到肠杆菌科细菌平均水平为0.98(lg(CFU/枚))和0.68(lg(CFU/枚));薛艳蓉等[31]报道蛋壳表面细菌数一般在103CFU/枚。传统微生物研究采用实验室分离培养,只有少数的微生物能够被成功培养鉴定出来,而且用时长、筛选菌属也不够广泛,不能进一步分析微生物群落组成及其多样性。宏基因组学克服了这一限制,以基因组技术为基础通过对环境中全部DNA系统全面的研究,从宏观角度准确高效地对微生物种类进行测定。本研究中选取安徽、北京、河北、吉林地区作为取样点,涉及范围广泛,利用宏基因组学技术经过对蛋源表面菌群多样性分析,结果表明以吉林地区为代表的北方,更适合作为蛋源的生产区,蛋源表面微生物的菌群多样性分析结果为后续消毒处理方法的优化奠定了一定的理论基础。
4 结 论
采用高通量测序的宏基因组学技术对4 个地区蛋源表面菌群多样性系统分析,不同地区蛋源表面主要微生物门水平和属水平都存在一定差异性,相对丰度差异更显著,北方地区与中部以南地区差异明显,北方地区更适宜大规模蛋鸡养殖。为保障液蛋制品的安全性,蛋制品企业应该根据蛋源表面微生物组成情况适当调整消毒处理方法,对液蛋制品加工行业有着重要意义。
