先天性心脏病基础研究进展
2021-06-02艾珊珊何爱彬
艾珊珊,何爱彬
1南方医科大学基础医学院生理教研室, 广州 510515 2北京大学分子医学研究所, 北京 100871
先天性心脏病(congenital heart disease,CHD)是指胚胎期心脏和大血管发育异常所致的畸形,包括心脏壁、瓣膜及血管畸形。CHD作为最常见的人类出生畸形,其患病率约为0.8%~1.2%[1]。2019年,
一项系统综述分析了全球1970—2017年新生儿CHD的患病情况,其中以室间隔缺损(ventricular septal defect,VSD)和房间隔缺损(atrial septal defect,ASD)比率最高(图1)。由于成年哺乳动物的心脏缺乏再生能力,其受损后无法恢复正常,因此CHD具有极高的发病率和致死率[2]。近年来,虽然科学家对CHD的发病机制进行了深入研究和探讨,但由于其致病机制复杂,仅20%的CHD致病基因得以鉴定,大部分CHD的发病原因仍无从解释,这极大限制了CHD临床治疗的开展[3]。因此,加快CHD的基础研究迫在眉睫。
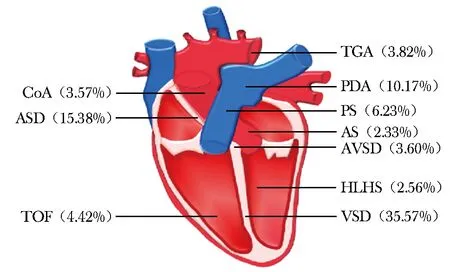
图 1 不同类型心脏畸形占先天性心脏病的比率[4]CoA:主动脉缩窄;ASD:房间隔缺损;TOF:法洛四联症;TGA:大动脉移位;PDA:动脉导管未闭;PS:肺动脉狭窄;AS:主动脉狭窄;AVSD:房室间隔缺损;HLHS:左心发育不全综合征;VSD:室间隔缺损
研究表明,导致CHD的突变基因多在心脏早期发育过程中发挥重要调控作用,包括心脏转录因子、心脏特异基因或某些信号通路分子[2]。然而,由于CHD的致病因素复杂,大部分CHD的病因无法仅通过遗传因素进行解释,如相同的CHD突变基因在不同患者的心肌组织中可引起不同类型的基因表达异常,相同的CHD致病突变可导致不同类型的心脏畸形。因此,对于CHD致病机制的深入了解,有助于寻找新的预防和治疗手段。本文将对CHD的致病因素及探究CHD致病机制的常用研究模型进行阐述,以期为临床诊疗提供理论依据。
1 遗传因素
自1949年Campbell首次报道CHD遗传因素以来[5],寻找CHD的遗传致病基因成为该领域的研究热点。根据基因突变类型,CHD的遗传致病因素可分为:点突变、染色体非整倍性、染色体拷贝数变异(copy number variation,CNV)和单核苷酸多态性(single nucleotide polymorphism,SNP)等[6]。
点突变通过改变靶基因的转录活性或改变蛋白质的功能而导致CHD,如Alagille综合征是由JAG1基因显性突变引起[7]。心脏的重要转录因子(如NKX2- 5、NKX2- 6、GATA4、GATA5、GATA6、IRX4、TBX20和ZIC3等)突变均可导致CHD[8]。关于CHD点突变的鉴定,最初主要通过家族性病例进行经典连锁分析,随着二代测序技术的快速发展,可直接通过高通量测序(包括外显子测序和全基因组测序)完成CHD点突变的检测。
除点突变外,CHD的发生常与某些形式染色体的异常相关,包括染色体结构和数目的异常。唐氏综合征(又称“21-三体综合征”)是一种最常见的染色体非整倍体形式,通常与ASD相关。染色体数目异常(非整倍性)同样可增加CHD的发病率。研究表明,约50%的唐氏综合征和特纳综合征患者可发展为CHD[9]。对于其他三体性疾病,如18-三体综合征和13-三体综合征,CHD的患病率高于50%[10]。此外,在法洛四联症、ASD、VSD、主动脉瓣狭窄和心室管腔狭窄的患者中均检测到大量结构异常的染色体[11]。队列研究表明,心脏特异基因在重复200 kb或缺失100 kb的情况下均可导致CHD,目前主要通过核型分析诊断染色体数目[11]。
一般而言,CHD患儿的CNV发生率较高。据报道,在某些法洛四联症[12]、左心室病变[13]及其他一些散发性CHD新生儿[14]中均存在CNV。因此,鉴定CHD的CNV有助于更好地认识其致病机理。Goldmuntz等[15]利用染色体微阵列(chromosomal micro-array analysis,CMA)技术鉴定了240种不同的CNV,这些基因在心脏发育过程中发挥重要调控作用,包括NRP1、NTRK3、MESP1、ADAM19和HAND1等。随着测序技术的迅猛发展,高通量二代测序技术正逐渐取代CMA用以鉴定染色体拷贝数的变化。
2 表观遗传因素
约20%的CHD由遗传因素导致,但多数情况下CHD的致病原因无法解释。越来越多的基础及临床研究表明CHD常伴随表观遗传图谱的改变。因此,深入了解CHD的表观遗传致病因素迫在眉睫。
表观遗传调控是指在不改变基因组序列的前提下,对基因表达进行调控的一种方式。除某些细胞(如配子细胞和免疫细胞)外,人体细胞都具有相同的基因型,表观遗传因素决定这些基因型相同的细胞表现不同的状态,发挥不同的功能。表观遗传调控包括DNA甲基化、组蛋白修饰、染色质重塑因子、长链非编码RNA(long noncoding RNA,lncRNA)、微小RNA(microRNA,miRNA)等。
2.1 DNA甲基化
DNA甲基化是一种重要的表观遗传调控方式,研究表明CHD患者具有特异的DNA甲基化图谱[16],DNA甲基化状态的改变与CHD的发生存在一定相关性。在DNA中,从5′→ 3′方向上连续的胞嘧啶-鸟嘌呤核苷酸称为CpG位点,在整个基因组中,CpG含量高的区域称为“CpG岛”。“CpG岛”是一段长约300~3000 bp,且不易被甲基化的区域。大约70%的基因启动子分布在“CpG岛”中,因此这些区域的甲基化常与基因表达“沉默”相关。除这种经典的调控方式外,“CpG岛”还可位于启动子之外,其甲基化在某些情况下可促进基因的表达[17]。
在心脏发育过程中,DNA甲基化控制基因的时空特异性表达。某些法洛四联症患儿心肌中NKX2.5和HAND2启动子区域甲基化水平升高,导致NKX2.5和HAND2表达下调[18]。比较CHD患儿(包括主动脉瓣狭窄、法洛四联症和VSD)与健康新生儿心肌的DNA甲基化图谱可发现,ASD/VSD新生儿心肌中存在52个基因的DNA甲基化异常,其中2个与冠状动脉疾病相关的基因APOA5和PCSK9出现高度甲基化[19]。
2.2 组蛋白修饰
染色体的基本结构单元为核小体,核小体由组蛋白八聚体组成。组蛋白可通过不同类型的共价修饰以改变染色体结构,影响蛋白质与DNA的结合,从而调控基因表达。组蛋白的共价修饰主要包括甲基化、乙酰化、泛素化、磷酸化和磺酰化等。组蛋白修饰可在不改变基因序列的情况下调控心脏转录因子的表达量及活性[20],因此组蛋白修饰紊乱是CHD的重要致病因素。组蛋白修饰的可逆性,使其成为治疗CHD的潜在靶点。
2.2.1 组蛋白去乙酰化酶
组蛋白乙酰化修饰通过减弱DNA与组蛋白的相互作用而促进基因表达,与之相反,组蛋白去乙酰化酶(histone deacetylase,HDAC)通过移除组蛋白乙酰化修饰而与基因表达“沉默”相关。研究发现,HDAC5和HDAC9双敲除的小鼠心肌细胞增殖与成熟受阻,可出现心室壁变薄及致死性VSD表型[21]。最新研究发现,单心室CHD患者的心肌HDAC活性异常增强,证实HDAC表达异常会导致CHD[22]。因此,HDAC抑制剂被用于治疗CHD。
2.2.2 多梳蛋白家族
组蛋白H3第27位赖氨酸的三甲基化修饰(H3K27me3)是一种广泛分布的抑制性组蛋白修饰,其主要催化蛋白为多梳蛋白家族的多梳抑制复合物2(polycomb repressive complex 2,PRC2)。PRC2蛋白的异常表达和H3K27me3修饰水平的紊乱均可导致CHD。心肌特异性EZH2(EZH2是PRC2的主要甲基转移酶)缺失的小鼠可出现VSD、ASD、心内膜垫增厚、上皮-间质转化受损及心肌细胞凋亡增加等表型[23]。研究发现,左心发育不良综合征患者的诱导性多能干细胞(induced pluri-potent stem cells,iPSCs)分化的心肌细胞中EZH2表达量下降,且一些重要的心脏发育基因TBX2、HEY2、NOTCH1、NKX2.5和HAND1的表达亦降低[24]。与PRC2作用相反,UTX和JMJD3是2个重要的H3K27me3去甲基化转移酶,与基因表达激活相关。UTX和JMJD3可上调GATA4的表达,同时UTX可在整个心脏发育过程中维持NKX2.5和TBX5的表达。UTX和JMJD3表达紊乱直接影响GATA4、NKX2.5和TBX5的表达[24- 25],因此UTX和JMJD3表达失调亦可导致CHD。
2.3 染色质重塑因子
染色质重塑复合物通过调控DNA结合蛋白的活性进而调节心脏的基因表达。BAF(BRG/BRM-associated factor)是一类重要的染色质重塑复合物。研究表明,BAF剂量的微小改变即可导致CHD,如BAF60c表达下调可严重影响心脏流出道的发育[26],BRG1表达紊乱可导致小鼠左心室发育严重缺陷[27]。另有研究发现,BAF可与心脏转录因子TBX1相互作用,调控右心室发育相关基因Wnt5a表达,TBX1和Wnt5a的表达下调均可导致右心室发育不良[28]。此外,BAF可与某些组织特异性蛋白相互作用,改变特定区域的组蛋白修饰状态,从而扰乱心肌细胞的基因表达[29]。
2.4 长链非编码RNA
lncRNA是由200多个核苷酸组成的无编码功能RNA。lncRNA通过参与组蛋白修饰等表观遗传调控过程调节基因表达,发挥其生物学作用。研究发现,某些VSD患者的心肌细胞可出现lncRNA表达失调,lncRNA与CHD的发生存在一定的相关性。据报道,孕有CHD胎儿的母体存在lncRNA表达异常,进一步证明了lncRNA参与CHD的发生[30]。
lncRNA uc.4过表达可抑制转化生长因子-β(transforming growth factor-β,TGF-β)信号通路,导致CHD的发生[31]。Anderson等[32]研究指出,抑制lncRNA Uph的表达可下调心脏特异性转录因子HAND2的表达,导致致死性右心室发育不全[32]。根据lncRNA与CHD的发生关系,lncRNA亦可作为潜在的生物标志物,用于CHD的产前检测。
2.5 微小RNA
miRNA是一类由内源基因编码的长度约22个核苷酸的非编码单链RNA分子,参与转录后基因表达调控。miRNA可与mRNA的3′UTR结合抑制RNA翻译或导致RNA降解,从而抑制基因表达。一种miRNA可与多种不同靶基因相互作用从而影响整个基因表达调控网络[33]。
miRNA在心脏发育过程中发挥重要的调控作用,参与调控心肌形态的发生、心肌细胞增殖和分化。因此,miRNA被视为CHD的重要治疗靶点,如miRNA- 499被证实可作为检测CHD的生物标志物[34]。研究表明,miRNA表达失调可导致右心室发育过程中重要转录因子TBX5、NOTCH1、HAND1和GATA3表达下降[35]。此外,Sucharov等[36]的研究也证明miRNA表达失调可导致严重的先天性心脏发育异常(如左心室发育不良)[36]。抑制miRNA- 184的表达可抑制心肌细胞增殖,并促使其凋亡,从而导致紫绀型CHD[37]。miRNA- 1通过促进HAND2(HAND2是心脏发育过程中的重要转录因子)的表达而促进心肌细胞增殖,并抑制caspase3剪切而降低心肌细胞的凋亡[38]。多项研究发现,法洛四联症患者的心肌组织中miRNA表达异常[39]。虽然miRNA与CHD的发生密切相关,但对于不同类型的CHD,特定的miRNA表达上调或下调及miRNA如何导致CHD的机制尚不清楚。
3 信号通路
心脏的发育和形成过程复杂,涉及心脏祖细胞的时空特异性谱系分化,以及心脏不同类型细胞的增殖、迁移和定位。因此,心脏的发育过程需协调不同信号通路以确保这些过程有条不紊进行。研究表明,在心脏的发育过程中,NOTCH、BMP和TGF-β等关键信号通路相互作用,共同促进心脏形态的建立[40]。
NOTCH信号通路在早期心脏发育过程中高度保守,参与细胞的发育、分化、增殖、凋亡、粘附和上皮-间质转化过程。NOTCH受体可与心内膜细胞相互作用而调控心脏的形态,包括心脏腔室和瓣膜形成[41]。NOTCH信号通路参与调控上皮-间质转化过程,调节心内膜-心肌相互作用,刺激肌小梁心肌细胞的产生。此外,NOTCH信号通路也参与调控心脏流出道的形成及肌小梁的致密化过程。因此,任何影响NOTCH信号通路的遗传或非遗传因素均可导致CHD的发生。动物实验证明,小鼠NOTCH受体和配体的突变可导致包括先天性心脏缺陷在内的多种先天性畸形[42]。研究证实,左心发育不良综合征患者亦存在NOTCH1突变[43]。
TGF-β信号通路在心脏发育过程中同样发挥必不可少的调控作用,β2-spectrin为蛋白Smads的配体,其在TGF-β信号通路中发挥重要作用。动物实验发现,小鼠β2-spectrin缺失可导致TGF-β信号通路失活,出现以左心室壁发育异常为主的心脏发育缺陷[44]。
4 营养素和代谢产物
近年来,关于母体孕前及孕早期营养素及代谢产物与CHD发生关系的研究成为热点。胚胎期,母体代谢产物对胎儿的生长具有直接而重要的影响。孕期营养不良、吸烟、紫外线辐射等外部环境因素均可导致表观遗传学的改变,引起基因表达紊乱,增加CHD的发病率。
DNA甲基化很大程度上取决于从食物中获取的甲基供体和辅因子的充足性,其是叶酸和蛋氨酸代谢的重要组成部分,因此孕期饮食可整体或在特定位点改变DNA的甲基化状态,从而改变表观遗传调控图谱。如母体长期营养不良,可能导致胎儿的印记基因胰岛素样生长因子2(insulin-like growth factor 2,IGF2)出现低甲基化,增加CHD患病风险。心脏发育过程中,叶酸及其衍生的同型半胱氨酸在心脏神经脊细胞迁移过程中发挥重要调控作用,围孕期母体叶酸缺乏可能导致CHD的发生。动物实验证实,围孕期缺乏维生素A的胚胎大鼠心肌中GATA4出现异常甲基化,导致胚胎出现CHD表型[45]。研究表明,孕期母体重度肥胖可增加主动脉分支缺损、大动脉移位、ASD和动脉导管未闭等CHD的患病率[46]。因此,孕期摄入健康、均衡的生物活性成分,有助于建立正确的表观遗传调控图谱,降低或预防CHD的发生。
5 研究模型
随着CHD基础研究的进步,成年CHD患者的存活率不断提高。然而,这些现状背后也引发了一些新的思考,如成年CHD幸存者的疾病复发风险是多少?是否存在预防CHD复发的相应措施?为研究这些问题,科学家们试图构建动物模型以重现特定的CHD表型,采用具有特定遗传学背景的动物或细胞模型开展CHD的致病机制研究,同时利用这些动物模型检测新的治疗策略。
在心脏发育和CHD研究领域,常用的动物模型包括果蝇、斑马鱼、青蛙、鸡、小型哺乳动物(小鼠)、大型哺乳动物(绵羊、狗和猪)以及体外干细胞分化系统[47]。不同的研究模型有其自身的优势和独特性。小鼠模型易于反向遗传操作,具有高度的序列保守性,具有与人类相同的心脏结构和心脏发育过程;其局限性在于制造转基因小鼠需花费较长时间和较昂贵的成本,且CHD小鼠通常在围产期死亡,从而限制了科学家对CHD小鼠的长期追踪研究。斑马鱼模型虽然易于进行基因操作,生产周期较短,但其心脏结构(仅有2个腔室)与人类不同[48]。
近年来,科学家们利用CHD患者的iPSCs作为体外模型研究CHD的致病机制[49]。虽然体外iPSCs系统无法重现CHD患者的心脏形态结构,但其具有与患者自身最一致的遗传学背景,可用来鉴定最相关的致病性分子调控机制。
6 小结
CHD的致病机制错综复杂,其发生是不同调控因素相互作用的结果,遗传因素的改变可直接或间接通过表观调控图谱改变基因表达;同时,表观遗传调控的紊乱亦可改变基因表达,反向调控染色质的状态,更深层次地影响基因表达。除遗传和表观遗传致病因素外,围孕期母体的营养素与代谢产物等环境因素也可改变表观遗传的调控状态,进而改变基因表达模式,导致心脏发育异常。正常生理状态下,遗传与非遗传因素相互影响、共同作用导致CHD的发生[50- 51](图2)。

图 2 遗传因素、表观遗传因素、环境因素相互作用及与先天性心脏病的关系[50- 51]
目前CHD领域尚存在诸多待解决的问题,为进一步了解CHD的分子发生机制,需对其致病机制进行综合分析。随着高通量测序技术的广泛应用,相信未来多学科交叉融合必将为人类揭示更精确、更丰富的CHD分子调控网络,为CHD患者带来新的曙光。
作者贡献:艾珊珊负责构思并撰写论文;何爱彬负责修订及审校论文。
利益冲突:无