马铃薯StCML基因家族鉴定及表达分析
2021-05-25张满仓卢海彬
张满仓,张 超,2,赵 朋,陈 越,陈 勤,卢海彬*
(1 西北农林科技大学 农学院/旱区作物逆境生物学国家重点实验室,陕西杨陵 712100;2 陕西省土地工程建设集团有限责任公司,西安 710753;3 西北农林科技大学 食品科学与工程学院/旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
植物在生长发育中会受到外界环境的多种胁迫作用,为了适应多变的环境,植物需要对环境信号做出反应,而Ca2+作为信号传递的第二信使具有至关重要的作用。当植物受到外界刺激时,会引起Ca2+内流[1-2],Ca2+信号的强弱和持续时间与刺激的性质有关[3]。钙感受器可以接收Ca2+信号并将其传递至下游[4]。
Ca2+感受器主要有钙调蛋白(calmodulin,CaM)、类钙调蛋白(CaM-like protein,CML)、钙依赖型蛋白激酶 (calcium-dependent protein kinase,CDPK)和类钙调神经素B (calcineurin B-like protein,CBL)[5]。CML是一类重要的钙离子感受器,含有数目不等的EF-hand结构域且不具有其他结构域[6]。EF-hand结构由29个氨基酸组成螺旋-环-螺旋(helix-loop-helix)结构,位于中心的12个氨基酸残基形成一个与Ca2+结合相关的转环(turn-loop)结构[7-8]。EF-hand结构中的天冬氨酸和谷氨酸与Ca2+结合,具有高度的保守性[9]。CML参与植物生长发育和多种外界胁迫的响应当中,对植物的生长有着重要的作用[10-12]。
目前已在多种植物中发现了CML基因,如拟南芥中鉴定到50个CML基因[13],水稻中有32个CML基因[14],番茄含有52个CML基因[15],大豆含有144个CML基因[16]。已有研究发现在NaCl处理、低温处理和脱落酸(ABA)处理下都能诱导拟南芥AtCML9基因的表达,其中NaCl对种子萌发的抑制作用主要通过ABA介导[17]。AtCML25可以通过调节K+内流调控拟南芥花粉萌发和花粉管的伸长[18]。AtCML42参与茉莉酸(JA)信号的转导,使用JA处理Atcml42突变体植株发现产生了更高的Ca2+内流,提高了对根长的抑制效果并且降低了食草性昆虫的取食,表明AtCML42负调控植物对JA的敏感性,进而提高了植物对草食性昆虫的抗性。此外,干旱胁迫下Atcml42植株的脱落酸含量升高,这表明AtCML42在干旱胁迫下抑制ABA的生物合成[19]。拟南芥AtCML43基因在根尖中特异表达,在水杨酸(SA)处理后表达量升高,进而调控植物免疫[20]。在番茄中,CML43参与了对丁香假单胞杆菌的免疫反应[21]。水稻类钙调蛋白基因OsMSR2在低温、干旱和高温多种环境胁迫下表达量均强烈上调。OsMSR2转基因拟南芥对ABA表现出超敏反应,表明OsMSR2通过ABA途径调控植株对干旱和盐分胁迫响应[22]。
马铃薯作为主要的粮食作物之一,具有种植范围广、产量高、营养价值丰富的特点[23-24]。本研究通过对马铃薯基因组数据库的筛选获得80个StCML基因,利用生物信息学分析了StCML家族基因的系统进化、保守基序、基因结构、基因复制、RNA-seq表达谱、GO富集分析和种间共线性,并检测了7个基因在低温、高温、盐胁迫和青枯菌侵染下基因的表达量,对StCML表达分析做了初步研究,对后续功能验证提供了思路,也为研究马铃薯StCML基因在生长发育和抗逆作用中的分子机制提供了理论基础。
1 材料和方法
1.1 植物材料
以马铃薯Désirée品种为实验材料。切取长势一致的脱毒苗顶芽放于含有MS培养基的组培瓶中,在植物生长箱生长15 d后进行相关实验。培养条件:温度22 ℃、光照时间16 h光照/8 h黑暗,相对湿度70%,光照强度10 000 Lx。
1.2 方 法
1.2.1 马铃薯StCML基因家族成员鉴定与系统进化分析在马铃薯基因组数据库网站下载蛋白序列(http://solanaceae.plantbiology.msu.edu/),在Pfam数据库(http://pfam.xfam.org/)下载CML家族的结构域模型文件(EF-hand_1:PF00036、EF-hand_6:PF13405、EF-hand_7:PF13499、EF-hand_8:PF13833),使用HMMER软件筛选马铃薯蛋白序列中含有EF-hand结构域的序列。将筛选到的蛋白序列在Pfam网站(http://pfam.xfam.org/search)和SMART网站(http://smart.embl-heidelberg.de/)进行结构域比对。使用Expasy网站(https://web.expasy.org/protparam/)分析StCML蛋白的理化特性。
使用MEGA10.1.7软件将马铃薯StCML和拟南芥AtCML蛋白序列构建系统进化树。采用邻接法(neighbor-joining,NJ),选用Poisson model,进行1 000次 bootstrap测试,AtCML蛋白序列由文献获得[13]。
1.2.2 马铃薯StCML染色体定位与基因复制分析在马铃薯基因组数据库网站下载基因的gff文件获取基因位置信息,使用TBtools软件[25]绘制染色体定位图。将序列相似性在70%以上且基因间隔在5个基因以内的基因定义为串联重复基因;在植物基因组复制数据库PGDD(http://chibba.agtec.uga.edu/duplication/)中查找StCML基因的片段复制基因。
1.2.3 马铃薯StCML基因保守基序、基因结构与顺式作用元件分析在MEME网站(http://meme-suite.org/)在线分析StCML蛋白基序种类及组成。从马铃薯基因组数据库网站下载StCML基因外显子、内含子和染色体位置数据文件,使用TBtools软件绘制基因结构图。选取基因前1 500 bp序列提交到PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站分析顺式作用元件,使用GSDD2.0网站(http://gsds.gao-lab.org/index.php)绘图。
1.2.4 马铃薯StCML基因GO富集与种间共线性分析提交StCML的氨基酸序列到Blast2GO软件(https://www.blast2go.com/)进行GO富集,分析分子功能、生物学过程和细胞组成。
使用perl脚本比对拟南芥AtCML与马铃薯StCML间的共线性基因,比对参数设为E≤10-10。
1.2.5 马铃薯StCML基因表达模式分析与多种胁迫处理在马铃薯基因组数据库网站下载StCML基因转录组测序数据,删除所有组织和胁迫下FPKM值均小于1的基因,计算Log2值并使用TBtools绘制热图。
选取长势一致的组培苗进行胁迫处理:低温处理将组培苗放入4 ℃植物生长箱培养6 h;高温处理组培苗放入35 ℃植物生长箱培养6 h;盐胁迫处理在超净工作台内将组培苗从培养基取出,转移到含有 150 mmol/L NaCl溶液的组培瓶中(使用dH2O配制并灭菌处理),放回植物生长箱培养12 h;青枯菌侵染在超净工作台内将组培苗从培养基取出,转移到含有GMI1000菌液(OD600= 0.01)的组培瓶中(使用dH2O配制并灭菌处理),放回植物生长箱培养12 h。使用未经处理的组培苗作为实验对照,每个处理有3个生物学重复。低温、高温和盐胁迫处理后,取植物的叶和茎;青枯菌处理后取植物的根系组织,使用液氮速冻后在-80 ℃冰箱保存。使用Thermo RNA提取试剂盒(#K0801)提取马铃薯植株RNA,使用艾科瑞逆转录试剂盒(AG11711)合成cDNA,使用QuantStudio 7荧光定量仪进行qRT-PCR反应。定量试剂使用Genestar 试剂盒(A301)。反应条件:95 ℃预变性 30 s,95 ℃变性 5 s,60 ℃退火 30s,72 ℃延伸 30 s,40个循环,使用2-ΔΔCT计算基因相对表达量。
2 结果与分析
2.1 StCML基因家族鉴定与系统进化分析
利用CML结构域模型文件,使用HMMER软件筛选出200个马铃薯蛋白序列,在Pfam网站和SMART网站分析序列的结构域,剔除无EF-hand结构的序列和含有其他结构域的序列后剩余97个蛋白,删除冗余基因和钙调蛋白基因后鉴定到80个StCML基因(表1)。使用Expasy网站预测蛋白生化特征,发现StCML蛋白的氨基酸数量主要在69~344之间,只有3个StCML蛋白的氨基酸数量大于500,蛋白分子质量主要在8.05~36.52 kD之间,有3个大于56 kD,等电点在3.69~9.48之间,大部分为酸性(88.75%)。

表1 马铃薯StCML基因家族序列特征Table 1 Sequence information of StCML genes in potato
为了分析马铃薯StCML基因的进化关系,使用相应蛋白质的氨基酸序列构建了系统进化树(图1),包含拟南芥的50个AtCML和马铃薯的80个StCML。它们被分为5个亚家族,每组所含蛋白质个数差异不大,分别含有32、24、20、25和29个蛋白,其中马铃薯StCML分别为18、12、14、12和24个。StCML和AtCML在5个亚家族中均有分布,StCML数目多于AtCML可能是马铃薯作为四倍体植物的基因组复杂性所致。
2.2 StCML染色体定位与基因复制分析
为了确定StCML基因在染色体的分布,在马铃薯网站查找了基因位置并绘制染色体定位图(图2)。StCML在12条染色体上均有分布,4号染色体最多,含有12个基因,8号染色体最少,只有1个基因,StCML1-4可能由于在染色体gap区域附近未被定位到12条染色体上。
StCML中含有13个串联复制基因,其中2组在4号染色体,另2组分别在10号和11号染色体。StCML中还含有18个片段复制基因,其中大部分集中在2号染色体,有2对基因均位于2号染色体,5对基因中的1个在2号染色体。由此推测StCML基因家族的扩张是由串联复制和片段复制共同作用产生的,其中2号染色体是片段复制的重要区域。
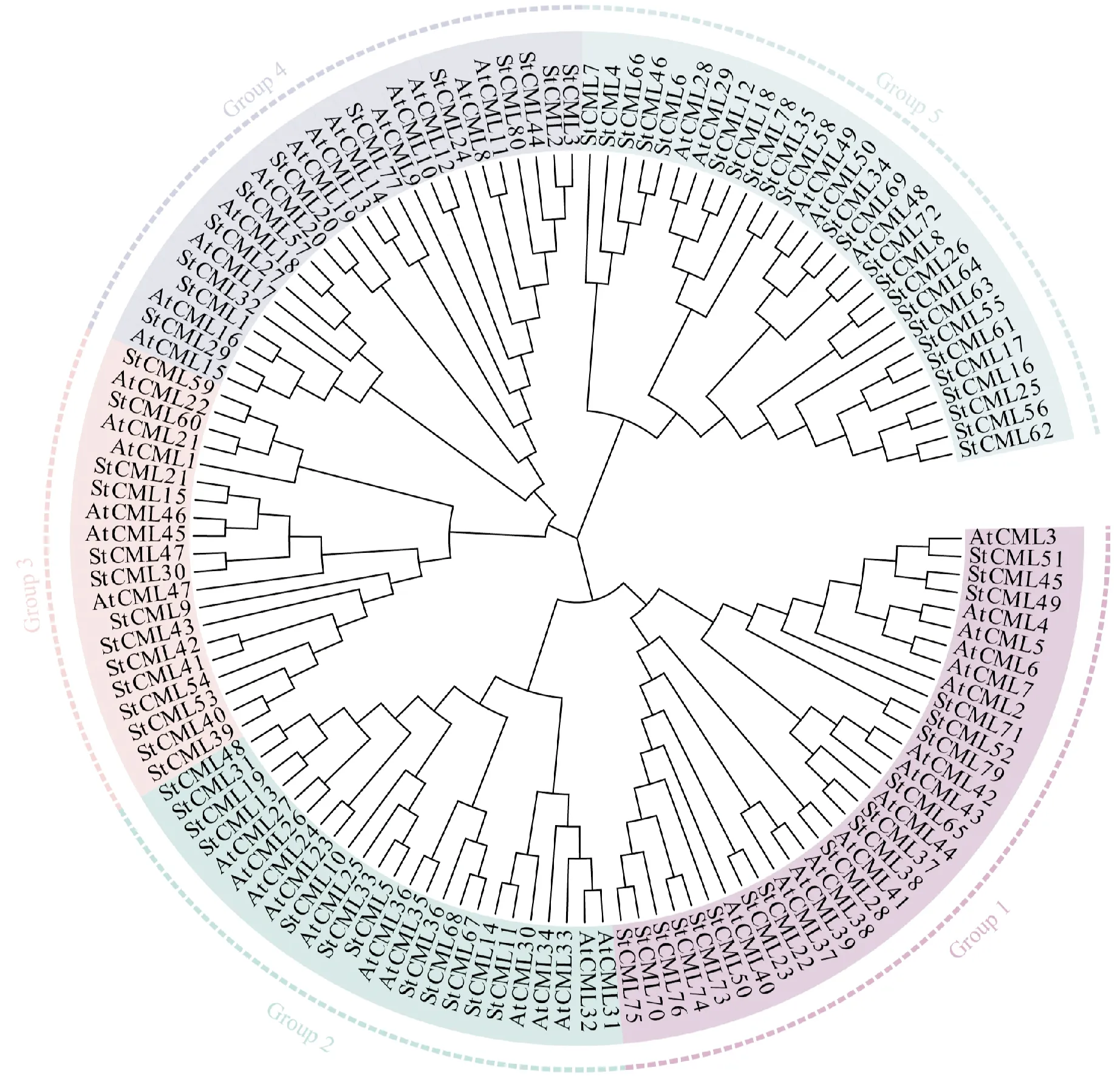
蓟色为第1亚家族;粉蓝色为第2亚家族;粉红色为第3亚家族;浅紫色为第4亚家族;浅青色为第5亚家族图1 马铃薯与拟南芥CML基因家族系统进化分析The thistle color is the first subfamily;powder blue is the second subfamily;pink is the third subfamily;lavender is the fourth subfamily;and light cyan is the fifth subfamilyFig.1 Phylogenetic relationship of CML genes in Arabidopsis and potato
2.3 StCML基因保守基序、基因结构与顺式作用元件分析
CML基因特征为具有EF-hand结构域,使用MEME网站预测StCML基序,结果显示StCML主要含有5个保守基序(图3,A),它们的长度在21~33个氨基酸(表2)。通过查询Pfam网站发现基序Motif 2、Motif 3、Motif 4属于EF-hand结构域。所有StCML均含有EF-hand结构域,其中除StCML32、StCML40和StCML70外均含有Motif 2;44个StCMLs中含有Motif 3基序;33个StCMLs含有Motif 4。Motif 2在5个亚家族中广泛分布,Motif 3和Motif 4主要分布在第1、2和4亚家族中,表明这3个亚家族具有更高的保守性,而不同亚家族间不同的基序结构则可能表明了它们功能的多样性。

表2 StCML蛋白保守基序信息表Table 2 List of the conserved motifs of StCML proteins
StCML基因结构如图3,B所示,外显子数目在1~14个之间,其中大部分基因(61个,76.25%)没有内含子;5个基因有1个内含子;其余14个基因含有多个内含子。第1、2和第3亚家族基因结构都较为保守,大部分基因均只有1个外显子,其比例分别为88.99%、91.67%和85.71%。第4和第5亚家族基因结构差异较大,表明StCML基因结构较为复杂。

左侧比例尺表示染色体长度,绿色方框表示串联重复基因,红色连线表示片段复制基因,染色体蓝色线条表示基因密度图2 StCMLs染色体定位和基因复制The scale on the left provides the relative length of chromosomes,green boxes indicate tandem duplicate genes,red lines indicate segmentally duplicate genes,blue lines on chromosomes indicate gene densityFig.2 Chromosomal location and gene duplication of StCMLs
为了研究StCML基因可能受到的调控机制,将基因起始密码子前1 500 bp序列提交到PlantCARE网站预测顺式作用元件分布,并使用GSDS2.0在线网站绘制顺式元件分布图(图4)。本研究发现2个与马铃薯生长发育相关的顺式作用元件:调节昼夜节律元件(circadian)和调控分生组织表达元件(CAT-box);与5种激素相关的顺式作用元件:脱落酸(ABRE)、茉莉酸甲酯(CGTCA-motif)、水杨酸(TCA-element和W-box)、生长素(TGA-element)、乙烯(ERE);5个胁迫响应的相关顺式作用元件:缺氧胁迫(ARE)、机械损伤(WUN-motif)、低温(MBS)、光照(I-box)和TC-rich repeats反应元件。StCML15启动子含有的顺式作用元件最多,有20个,StCML5启动子含有顺式作用元件最少,只有2个。StCML基因共含有126个ABRE,分布在56个基因中;含有103个ARE,分布在58个基因中;含有97个ERE,分布在52个基因中。响应逆境的相关顺式作用元件在所有StCML基因中均有分布。顺式作用元件分析表明StCML基因可能参与到了植物生长发育和多种激素及胁迫响应中。
2.4 StCML基因GO富集与种间共线性分析
为了进一步了解StCML蛋白功能,使用Blast2GO软件进行了GO富集,分析它们在生物学过程(Biological process,BP)、分子功能(Molecular function,MF) 和细胞成分(Cellular component,CC)中的作用(图5,A)。由GO富集分析发现80个StCML蛋白均具有离子结合能力(ion binding),这表明了它们可能具有Ca2+结合功能。在生物学过程分析中,发现27个蛋白参与了细胞对刺激的反应(cellular response to stimulus),除StCML78外,其余26个蛋白(StCML1~3、5、6、22~24、27~29、32、33、43、44、46、50、57、58、66、70、73~76、80)还参与了细胞调节(regulation of cellular process)、细胞通讯(cell communication)和信号转导(signal transduction),其中17个蛋白(StCML1、5、22、23、27~29、32、33、43、50、57、70、73~76)还参与了细胞成分的构成(cellular component organization or biogenesis),并且这17个蛋白在分子功能中具有酶调节活性(enzyme regulator activity)。具有这些功能的StCML主要分布在第1亚家族和第4亚家族。8个蛋白(StCML4、6、7、11、27、46、58、66)参与细胞质(cytoplasm)的构成,它们与StCML30、StCML47和StCML20共11个蛋白参与了细胞器(organelle)和细胞内部结构(intracellular anatomical structure)的构成。根据结果推测StCML在环境刺激下通过接收Ca2+信号参与了细胞调控并且可能参与了细胞多个成分的构成。
绘制AtCML和StCML基因共线性图发现存在20对共线性基因(图5,B),在5个亚家族中均有共线性基因分布。马铃薯2号染色体是含共线性基因最多的染色体(7个);拟南芥3号染色体含共线性基因最多(6个)。StCML15和StCML21均与AtCML45和AtCML46为共线性基因;StCML19与AtCML23和AtCML24为共线性基因;StCML31与AtCML26和AtCML27为共线性基因。AtCML45与StCML15、StCML21和StCML30是共线性基因。其中StCML15、StCML21和StCML31为片段复制基因,AtCML23和AtCML24与AtCML26和AtCML27分别为2对片段复制基因[13]。这表明了马铃薯与拟南芥CML基因具有一定的相似性。
2.5 StCML基因表达分析
为了研究StCML基因在马铃薯生长发育和胁迫响应下的表达,在马铃薯基因组数据库网站查找StCML基因在不同组织中和多种胁迫下的RNA-seq数据并绘制热图(图6)。共统计了StCML基因在花、叶、叶柄、根、芽、雄蕊、匍匐茎、块茎、愈伤、花瓣、心皮和萼片中的组织表达,以及植株在盐(Salt)、热(Heat)、6-苄氨基嘌呤(BAP)、甘露醇(Mannitol)、脱落酸(ABA)、生长素(IAA)、赤霉素(GA3)、晚疫病菌(P.infestans)、β-氨基丁酸(BABA)和苯并噻二唑(BTH)处理下基因的表达。6个基因(StCML10、24、25、28、47、71)在公共数据库各个组织中表达量未被检测到,12个基因(StCML6、10、12、15、20、24、25、26、28、47、71、78)在公共数据库多种胁迫处理下未被检测到表达。除去这些表达量极低的基因,其余基因至少在一个组织中表达。特异表达的组织主要有花、叶柄、芽、雄蕊、匍匐茎和块茎,分别有8、5、7、12、18和4个基因特异表达。StCML基因主要响应盐、热、干旱和赤霉素处理,分别有13、10、8和9个基因表达量显著上调。
2.6 StCML基因多种胁迫响应分析
为了进一步研究StCML基因对非生物胁迫的响应,使用低温、高温、盐和青枯病菌GMI1000处理马铃薯材料Désirée进行qRT-PCR检测StCML基因的表达,选取7个在转录组数据中表达量较高且对盐和热胁迫有响应的基因(StCML8、11、13、21、39、53、60)作为研究对象(图7)。结果发现在4种胁迫处理下均有基因响应,且不同基因的表达存在较大差异。与未处理的植株相比,在低温胁迫下StCML13、StCML21和StCML53表达量分别上调至6.47、6.76和2.47倍;高温胁迫下StCML11、StCML21和StCML39表达量分别上调至3.43、2.43和6.20倍,StCML8和StCML60下调至0.49和0.29倍;盐胁迫下StCML21和StCML60表达量上调至8.2倍和4.5倍。青枯菌处理下只有StCML53上调至2.6倍,StCML8、StCML13、StCML21和StCML60分别下调至0.35、0.03、0.04和0.13倍。StCML21在4种处理下均有响应,其他基因也至少在一种处理下表达量有显著变化,这表明StCML基因在多种环境胁迫中都可能具有作用。

下方比例尺表示蛋白长度(A)和基因长度(B)图3 StCML基因家族保守基序(A)及基因结构(B)分析The scales below provide the length of proteins (A) and genes (B) respectivelyFig.3 Conserved motif(A)and gene structure(B)analysis of StCML genes

下方比例尺表示基因启动子长度图4 StCML启动子顺式作用元件预测The scales below provide the length of promotersFig.4 Predicted cis-elements in StCML promoters
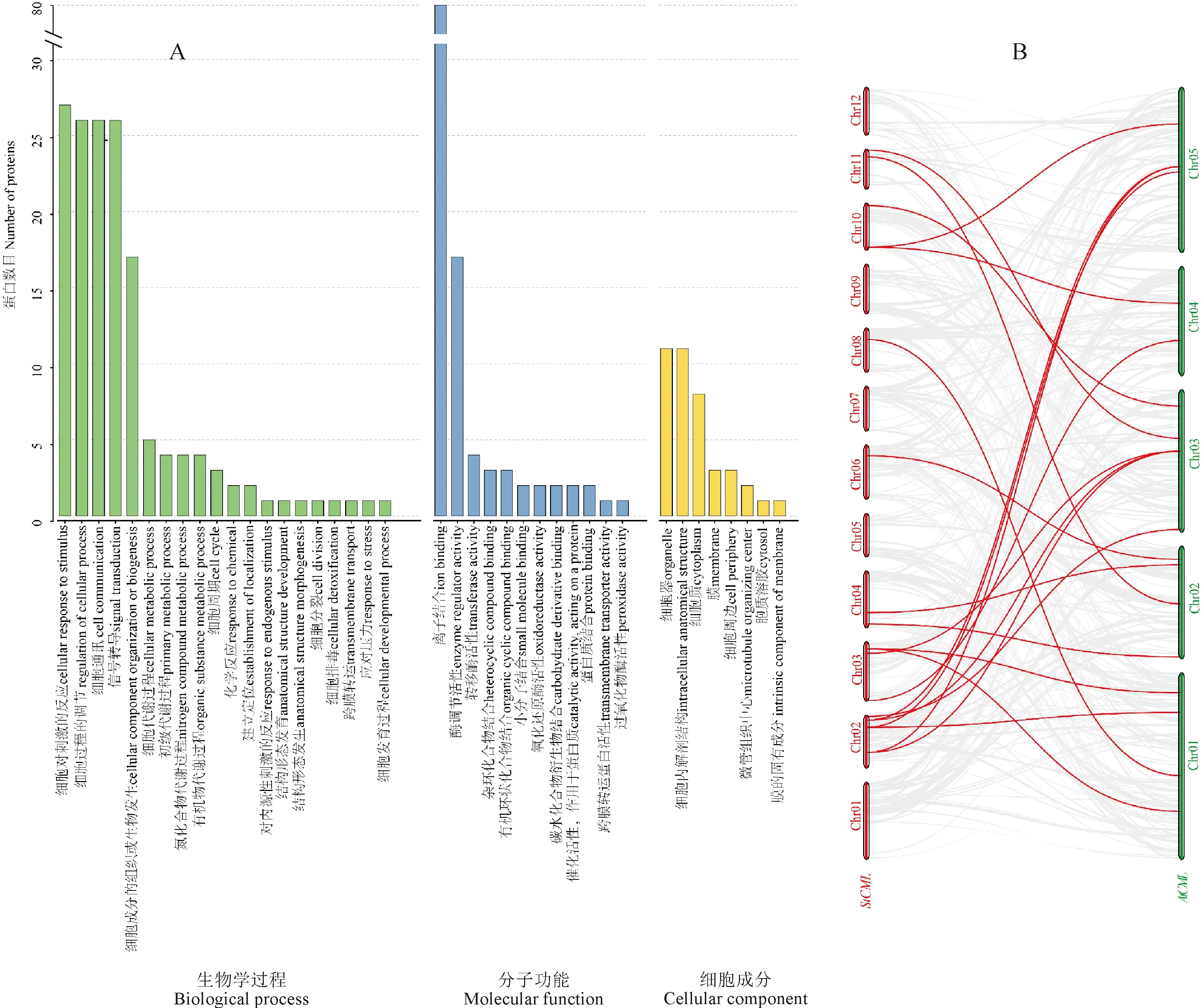
图5 StCMLs蛋白的GO富集分析(A)及拟南芥和马铃薯CMLs共线性分析(B)Fig.5 The information of gene ontology of StCMLs(A) and Orthologous relationship of CML genes in Arabidopsis and potato(B)
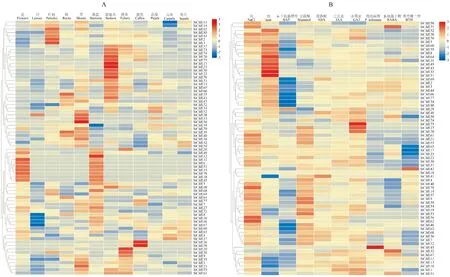
图例表示标准化的FPKM值。红色表示高表达水平,蓝色表示低表达水平图6 StCML基因在不同组织和器官中的表达谱(A)及多种胁迫下的表达模式(B)Legends represent the normalized FPKM values.Red means high expression level,while blue means low expression levelFig.6 Expression profiles of StCMLs in different tissues and organs (A) and expression patterns under different stresses(B)

不同小写字母表示基因表达量存在显著差异(P<0.05)图7 多种胁迫下StCML基因的相对表达Different lowercase letters indicate significant difference in gene expression (P<0.05)Fig.7 Relative expression levels of StCML genes under different stresses
3 讨 论
本研究通过生物信息学方法鉴定到了80个马铃薯StCML基因,StCML具有典型的EF-hand结构域,在马铃薯12条染色体上均有分布。与拟南芥AtCML构建系统进化树发现StCML可分为5个亚家族,每个亚家族都有至少12个StCML,分布较为均衡。StCML基因家族中分别含有13个(16.25%)串联复制基因和18个(22.5%)片段复制基因。基因复制事件主要发生在2号染色体。拟南芥中含有11个(22%)串联复制基因和27个(54%)片段复制基因[13]。相比之下拟南芥中基因复制的比例更高。
马铃薯StCML基因中61个(76.25%)基因没有内含子,与之相似的是拟南芥AtCML基因家族中有37个(74%)基因不含内含子[13],水稻OsCML基因家族中有24个(75%)基因不含内含子[14],番茄SlCML基因家族中有31个(59.62%)基因不含内含子[15]。有研究表明无内含子或内含子较少的基因在植物中可以更快地表达,这使得植物可以迅速应对外界刺激[26],这可能是CML基因快速参与调控的一个原因。
马铃薯StCML与拟南芥AtCML具有20对共线性基因,每对共线性基因均在同一亚家族,这表明了StCML同组间基因的保守性。与StCML具有共线性的拟南芥基因已有多个被报道。如与StCML49共线性的AtCML5位于高尔基体,参与内小泡的运输[27];与StCML24共线性的AtCML8基因突变体Atcml8在水杨酸和NaCl处理下表达量均上调[28],并且在抵御丁香假单胞杆菌的侵染中发挥作用[29];与StCML49共线性的AtCML24和AtCML23共同参与调控植物对光照时间的感知,通过Ca2+信号调节NO的积累,调控植物开花[30];与StCML15共线性的AtCML46是植物免疫的负调节因子,负调节水杨酸的积累,双突变体cml46cml47提高了对丁香假单胞杆菌的抗性[31]。
通过马铃薯基因组数据库网站的RNA-seq数据和qRT-PCR实验发现StCML基因在多种胁迫下均有响应,这与预测到的顺式作用元件功能相吻合,但不同基因间的表达具有较大差异。前人研究表明番茄SlCML在低温、干旱、盐、脱落酸和乙烯利处理下均有基因响应[15];白菜BrCML在高温胁迫下38个基因上调,18个基因下调[32];潘那利番茄SpCML在干旱、盐和低温胁迫下根中分别有14、12和21个基因表达量上调,在脱落酸、赤霉素和水杨酸处理下,叶片中分别有24、40和44个基因表达量上调[33]。这些研究表明CML基因在多种胁迫下均有响应,可能参与到多种植物对胁迫的调控作用,也印证了StCML基因功能的多样性。
本研究鉴定了马铃薯StCML基因家族成员,通过生物信息学方法研究了基因的物理特性、家族进化、基因结构、顺式作用元件、基因定位和基因复制等方面,并通过qRT-PCR分析了其在多种环境胁迫下的表达。为后续深入研究StCML基因的功能奠定了基础,也为寻找胁迫调控基因提供了参考。
