家蚕丝氨酸蛋白酶基因BmSPH33对BmNPV诱导的免疫响应
2021-05-25张永红邵榆岚苏振国白兴荣
张永红,邵榆岚,苏振国,白兴荣
(云南省农业科学院蚕桑蜜蜂研究所,云南 蒙自 661101)
【研究意义】丝氨酸蛋白酶(Serine protease,SP)是一类以丝氨酸为催化中心的蛋白质水解酶基因家族,在昆虫体内参与了食物消化、发育、先天免疫等[1-3]。家蚕丝氨酸蛋白酶基因家族包括BmSP(Bombyxmoriserine protease,BmSP)和同源体BmSPH(Bombyxmoriserine protease homolog,BmSPH)[4],它们是家蚕先天免疫通路中调控级联的关键酶类[5],所以研究BmSPs基因在家蚕免疫应答方面的作用具有重要意义。【前人研究进展】丝氨酸蛋白酶(SPs)普遍存在于生物体中,是一类含有保守催化三联残基His、Asp和Ser的蛋白质水解酶[6],它们通常以非酶催化活性的前体存在,其前肽必须被修剪才能激活,赵萍等[7]在家蚕基因组中共鉴定到143个BmSPs基因,分别标记为BmSP1—BmSP143,其中包括51个BmSPs和94个BmSPHs。BmSPs基因在家蚕体内组织和时期表达量不同,进而发挥着不同的功能[8]。已有研究发现BmSPs基因家族成员中BmSP2作为中肠消化酶,对家蚕核型多角体病毒(Bombyxmorinucleopolyhedrovirus,BmNPV)具有免疫作用[9],且BmSP25基因在BmNPV的感染诱导下产生了免疫响应[10]、BmSP142在家蚕感染BmNPV与BmBDV(Bombyxmoribidensovirus,BmBDV)后均表现为上调[11],BmSP36与BmSP141参与家蚕的食物消化过程[12-13]、BmSP95在家蚕蛹皮蜕化过程中发挥着功能[14]。丝氨酸蛋白酶同源物(SPHs)在氨基酸序列上与SPs相似,但由于一个或多个催化残基发生突变而缺乏酶催化活性[15],但它们参与了前酚氧化酶(proPO)激活系统,导致黑色素化。研究发现家蚕丝氨酸蛋白酶同源体BmSPH-1具有幼虫-蛹转化、蛹表皮黑色素化和家蚕天然免疫的双重作用[16]。【本研究切入点】家蚕丝氨酸蛋白酶BmSPs在家蚕肠道内主要参与食物消化,还参与宿主的免疫应答过程,至今未见BmSPH33基因对BmNPV免疫响应机理方面的报道。【拟解决的关键问题】获得BmSPH33基因序列并结合其转录情况,另外家蚕感染BmNPV后该基因表达模式明确诱导响应机理,为弄清BmSPH33基因在参与家蚕先天免疫方面发挥作用提供依据。
1 材料与方法
1.1 供试材料
供试家蚕品种为大造(P50),纯化后的BmNPV病毒粒子由云南省农业科学院蚕桑蜜蜂研究所保存。TaKaRa宝生物工程(大连)有限公司产品:RNAiSo Plus Total RNA提取试剂盒、One Step RNA PCR Kit (AMV)与PrimeScriptTMRT reagent kit with gDNA Eraser反转录试剂盒、PCR Mixture、SYBR Primix ExTaqTMII (Tli RNaseH Plus) qRT-PCR试剂盒;生工生物工程(上海)股份有限公司产品:PCR引物、琼脂糖凝胶回收试剂盒等试剂。
1.2 试验方法
1.2.1 家蚕组织样品收集 家蚕大造(P50)品种幼虫在27 ℃条件下用新鲜桑叶饲养至5龄第3天,解剖幼虫并收集头部、血液、体壁、中肠、脂肪体、丝腺、精巢和卵巢;BmNPV添毒实验中,正常饲育的5龄起蚕平均分为2组,其中一组按照BmNPV多角体悬浮液5.13×107PIB·mL-1的浓度喂食家蚕10 μl·头-1;另外一组同样喂食10 μl ddH2O。两组家蚕添食后常规桑叶饲养。在不同时间4、8、12、24和48 h分别解剖家蚕并收集中肠样品,其中添食ddH2O家蚕的中肠样品为对照。
1.2.2 BmSPH33序列分析 登录家蚕基因组数据库SilkwormDB,获取BmSPH33基因ORF序列,利用生物学在线软件对BmSPH33蛋白的分子量、等电点、信号肽、蛋白质二级结构、三级结构进行预测,其中理论等电点与分子量预测软件ExPASy(http://web.expasy.org/)、信号肽预测软件SignalP v5.0(http://www.cbs.dtu.dk/services/SignalP/)、功能结构域软件CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)、二级结构软件PSIPRED(http://bioinf.cs.ucl.ac.uk/psipred/)和三级结构预测软件SWISS-MODLE(https://swissmodel.expasy.org/);利用MEGA5.0软件对BmSPH33与其它物种SPH33序列生成.fasta与.mega文件,结合GeneDoc软件进行多序列比对;根据MEGA5.0 中Neighbor-joining方法构建系统发生树[17]。
1.2.3 家蚕组织样品总RNA提取及cDNA合成 利用液氮将家蚕不同组织样品和不同时期处理的中肠样品充分研磨,按照RNAiSo Plus Total RNA提取试剂盒操作步骤抽提RNA,超微量核酸蛋白测定仪检测RNA样品浓度和纯度,将抽提的RNA根据One Step RNA PCR Kit (AMV)说明反转录成cDNA。
1.2.4 家蚕不同组织BmSPH33基因转录情况 设计BmSPH33基因RT-PCR引物(表1)。以家蚕肌动蛋白基因Bmactin3为内参基因,家蚕不同组织cDNA为模板进行PCR扩增,反应体系为25 μl:10×PCR buffer(Mg2+plus) 2.5 μl,2.5 mmol·L-1dNTPs 2 μl,5 U·μl-1r-Taq聚合酶 0.25 μl,10 μmol·L-1上下游引物 0.5 μl,cDNA模板0.5 μl,ddH2O补齐25 μl;反应条件为:94 ℃ 4 min;(94 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s)× 30 cycles;72 ℃ 10 min;12 ℃ forever,扩增产物用1 %的琼脂糖凝胶检测。
1.2.5 qRT-PCR检测BmSPH33基因对BmNPV感染的响应 利用primer5.0设计BmSPH33基因荧光定量特异引物(表 1),以不同时间段ddH2O与BmNPV处理的家蚕中肠组织cDNA为模板,Bmactin3为内参基因进行qRT-PCR,参照SYBR Primix ExTaqTMII (Tli RNaseH Plus)试剂盒说明进行配制,反应体系为20 μl(SYBR Premix ExTaqII 10 μl,ROX Reference Dye 50× 0.4 μl,F/R Primers 0.4 μl,cDNA 1 μl,ddH2O 补齐20 μl),扩增程序:95 ℃30 s;(95 ℃ 30s,60 ℃ 30 s)× 40 cycles。ABI StepOnePlus仪器记录实验结果,不同处理的样品设置3个重复,收集基因的Ct值数据,最后根据2-△△Ct计算BmSPH33基因的相对表达量[18]。
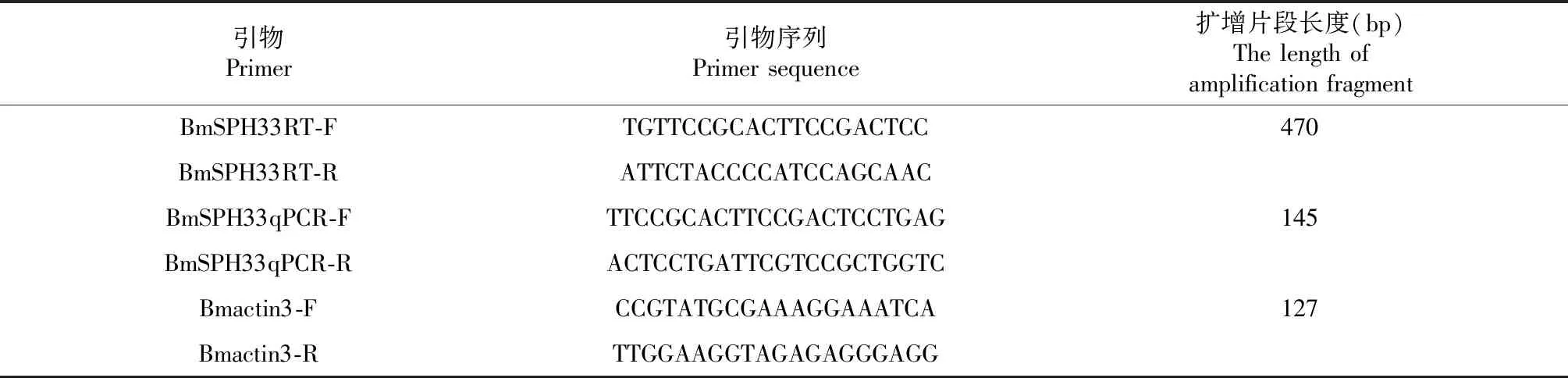
表1 试验引物Table 1 Research primers
2 结果与分析
2.1 BmSPH33基因序列生物信息学分析
对SilkwormDB公布的BmSPH33基因ORF进行序列分析,BmSPH33ORF全长为840 bp,编码279个氨基酸残基,软件预测无信号肽序列,为非分泌型蛋白,去除信号肽后理论分子量大小为22.95 kDa,等电点为5.98。NCBI CDD在线预测软件发现,40~277位氨基酸为类胰蛋白酶丝氨酸蛋白酶结构域(Trysin-like serine protease domain,Tryp_SPc domain)。利用PSIPRED Server在线软件预测BmSPH33蛋白质的二级结构(图1),预测结果发现,BmSPH33蛋白由5个α螺旋、14个β折叠和一些无规则卷曲构成;利用SWISS-MODEL软件预测三级结构(图2),其二级结构及三级结构预测特征一致而且以β折叠构成为主。

图1 预测的BmSPH33蛋白质二级结构Fig.1 Secondary structure for BmSPH33 protein by server
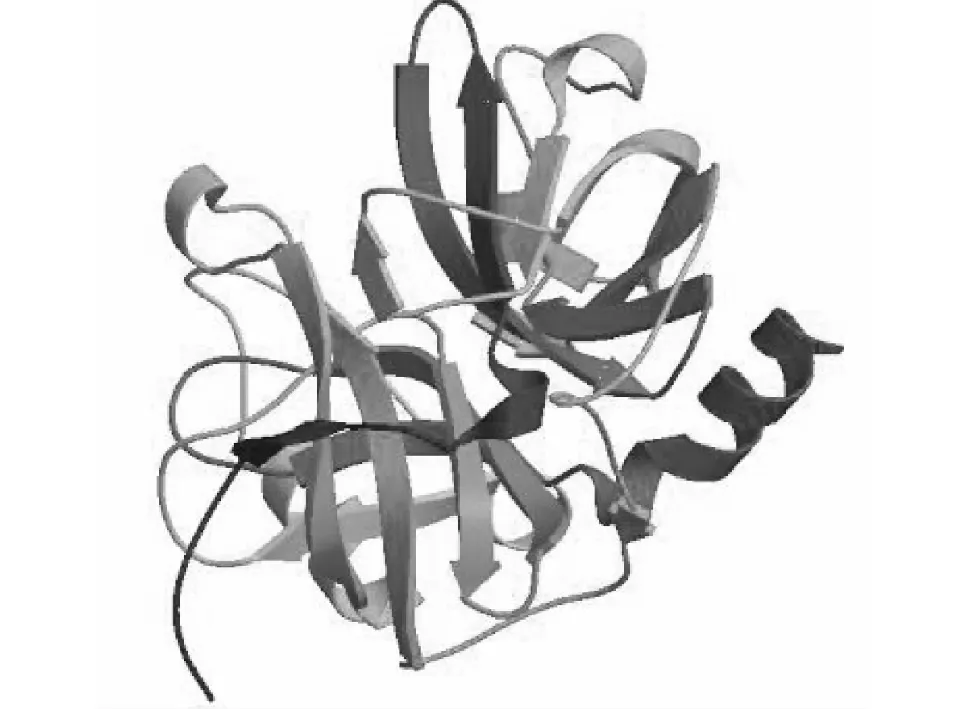
图2 预测的BmSPH33蛋白质三级结构Fig.2 Tertiary structure of for BmSPH33 protein by server
BmSPH33的氨基酸序列(GenBank登录号:AK378171.1)与NCBI上登录的蓓带夜蛾SPH33(GenBank登录号:ACR15983.2)进行序列比对。结果显示,各序列中均含有类胰蛋白酶丝氨酸蛋白酶结构域(图3)。基于不同物种SPH33蛋白氨基酸序列,利用MEGA5.0软件,采用邻接法对构建系统发生树(图4),遗传进化结果显示,BmSPH33与蓓带夜蛾(Mamestraconfigurata)和冬尺蠖蛾(Operophterabrumata)SPH33在亲缘关系上较近,其中不同节肢动物的物种SPH33未聚在同一分支,而植物大叶杜鹃(Danausplexippusplexippus)SPH33与家蚕(Bombyxmori)、蓓带夜蛾(Mamestraconfigurata)和冬尺蠖蛾(Operophterabrumata)遗传距离相近。

BmSPH33:家蚕SPH33;MacoSPH33:蓓带夜蛾SPH33BmSPH33:Bombyx mori SPH33;MacoSPH33:Mamestra configurata SPH33图3 BmSPH33与MacoSPH33氨基酸序列比对分析Fig.3 Sequence alignment of amino acid between BmSPH33 and MacoSPH33
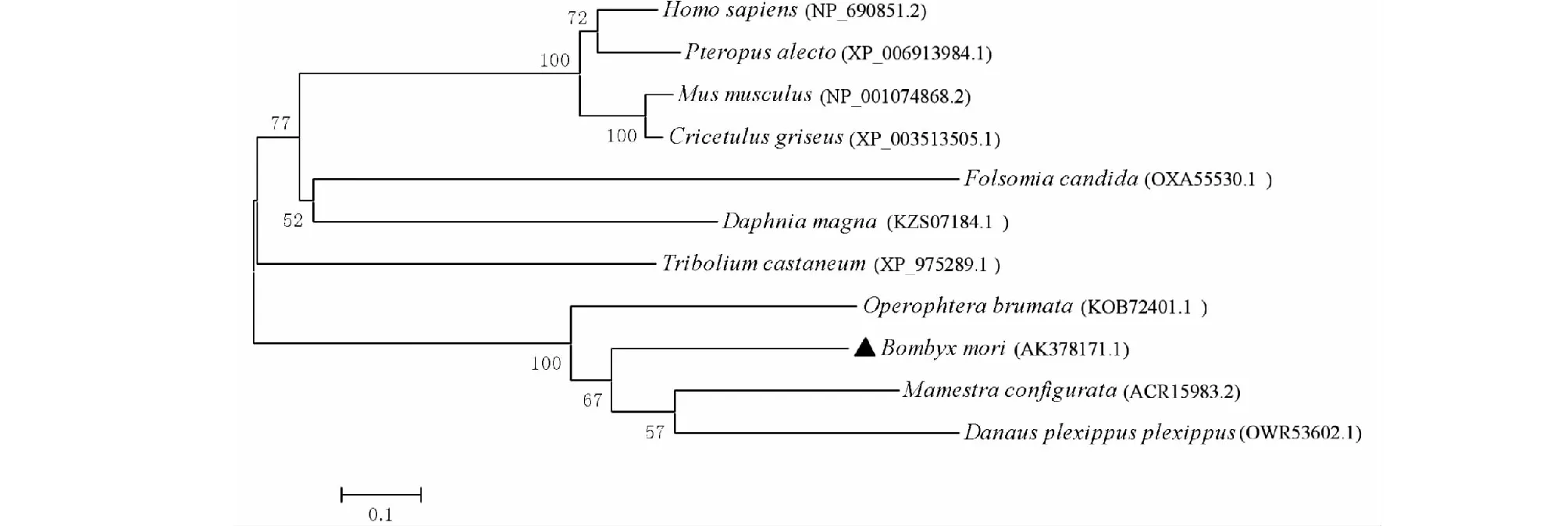
图4 BmSPH33与其同源氨基酸序列进化分析Fig.4 Evolution analysis of amino acid between BmSPH33 with other homologues sequence
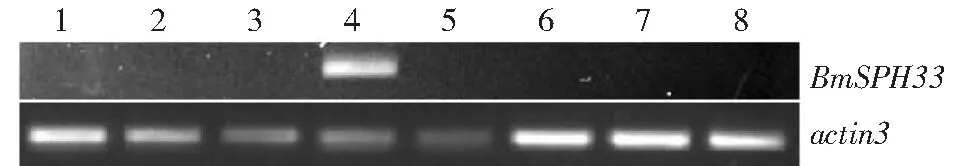
1:头;2:血液;3:体壁;4:中肠;5:脂肪体;6:丝腺;7:精巢;8:卵巢1. Head;2: Hemocyte;3: Integument;4: Midgut;5: Fat body;6: Silkgland;7: Testis;8: Ovary图5 家蚕不同组织BmSPH33基因转录分析Fig.5 Transcription analysis of BmSPH33 gene in different tissues of Bombyx mori
2.2 BmSPH33基因组织表达谱分析
利用RT-PCR对5龄第3天家蚕各组织BmSPH33基因转录情况进行检测,检测结果发现,BmSPH33在家蚕中肠组织中特异性表达(图5)。
2.3 BmNPV感染家蚕后BmSPH33的转录分析
为探讨BmSPH33基因在家蚕感染BmNPV病毒反应中可能的作用,利用实时荧光定量qRT-PCR对对照组(添食ddH2O)和实验组(添食BmNPV)BmSPH33基因表达情况进行检测,确定BmSPH33基因转录水平是否发生变化。定量结果发现家蚕在感染BmNPV后BmSPH33基因发生响应(图6),在感染BmNPV 4 h后BmSPH33基因转录水平升高,而感染8、12、24、48 h时呈现下调,这说明家蚕在感染BmNPV后产生了应激响应,其中BmSPH33基因mRNA转录水平发生变化。BmSPH33基因mRNA转录反应受到BmNPV病毒的诱导,提示BmSPH33基因在大造(P50)家蚕品种的抗病毒作用中发挥功能。
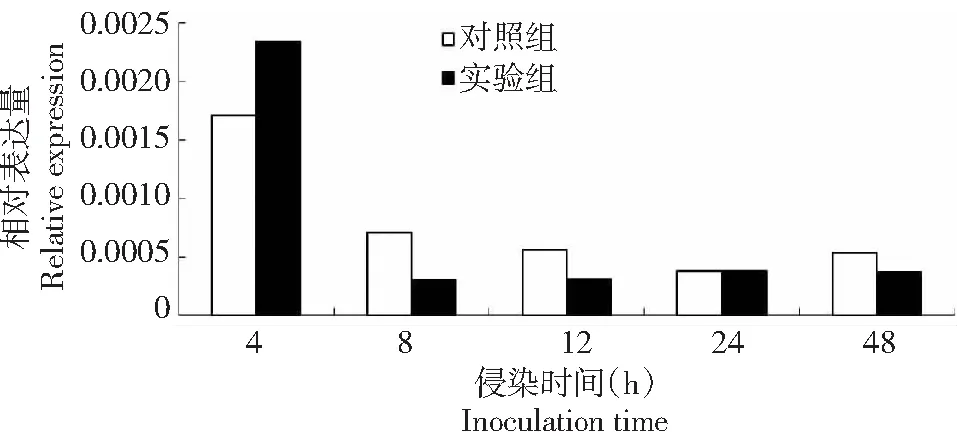
图6 BmNPV对BmSPH33基因转录水平的影响Fig.6 Effects of BmNPV inoculation on transcriptional level of BmSPH33 gene
3 讨 论
丝氨酸蛋白酶SPs参与了昆虫的消化、发育和免疫应答,是一类重要的蛋白水解酶。研究BmSPH33与MacoSPH33氨基酸序列比对发现,它们含有保守的Trypsin-like serine protease的结构域,类胰蛋白酶丝氨酸蛋白酶是一种Clip结构域丝氨酸蛋白酶,被认为能参与昆虫先天免疫的蛋白酶[19-20],其结构含有保守的TAAHC、DIAL和GDSGGP特征基序。昆虫经过丝氨酸蛋白酶等一些酶级联反应能将非活化前体酚氧化酶酶原(prophenoloxidase,proPO)催化形成活性的酚氧化酶原(Phenoloxidase,PO),PO能把酚类氧化成醌类物质,通过生成黑色素沉积在病原物表面,进而黑化和包裹作用消灭病原微生物[21]。SPs蛋白通过剪切羧基端的一些特殊氨基酸而活化proPO[5]。
DNA microarray研究发现64个表达的SPs和SPHs基因中有38个基因中肠转录,其中大部分在中肠表达,一些中肠SPHs基因可能由于缺少活性位点残基而处于非活动状态,但在其它鳞翅目昆虫中也发现了表达[22],而且SPH1和SPH2还参与烟草天蛾的proPO活化过程[23];家蚕BmSPH1证实了还参与家蚕的免疫响应[16, 24]。家蚕中肠是消化桑叶并吸收营养的重要场所,BmNPV病毒粒子随着食物进入中肠,它作为防御病毒入侵的第一道屏障,BmSPH33基因在家蚕中肠组织中特异表达(图5),研究结果发现,BmSPH33基因转录水平在感染4 h时上调,其余时间点(8、12、24和48 h)下调,基因转录水平发生变化,表明BmSPH33基因表达受BmNPV诱导而参与家蚕幼虫免疫应答过程。
丝氨酸蛋白酶在真核细胞和原核生物中是高度保守和普遍存在的,它们已经进化成一个丰富而功能多样的酶群。丝氨酸蛋白酶是一类主要的消化蛋白酶,占鳞翅目昆虫消化活性的95 %[25],由于BmSPH33与其BmSPs成员具有Tryp_SPc domain,昆虫丝氨酸蛋白酶家族作为丝氨酸为活性中心的典型蛋白质水解酶,结合其中肠特异表达,说明该蛋白主要参与食物消化,家蚕感染BmNPV后基因转录水平发生变化,推测该基因在家蚕免疫方面起到了辅助作用,但具体功能还有待进一步验证。
4 结 论
BmSPH33基因在家蚕中肠组织中特异表达,且在家蚕感染BmNPV后发生响应,推测该基因参与家蚕免疫应答过程,结合BmSPH33含有高度保守的胰蛋白酶样的底物基序,为弄清丝氨酸蛋白酶同源体BmSPHs在消化及免疫应答方面的机制提供参考。
