基因突变提高毕赤酵母表达内切β-1,3-葡聚糖酶活性
2021-05-18金树霞李菲菲刘丽萍詹晓北郑志永高敏杰
金树霞, 王 力, 朱 莉, 李菲菲, 刘丽萍,詹晓北, 郑志永,4, 高敏杰*
(1. 江南大学 生物工程学院,江苏 无锡214122;2. 江苏艾津农化有限责任公司,江苏 南京211511;3. 无锡格莱克斯生物科技有限公司,江苏 无锡2141254;4. 江南大学 环境与土木工程学院,江苏 无锡214122)
热凝胶主要由土壤杆菌在氮源受限的情况下合成[1],是一种不溶于水且由β-1,3-糖苷键连接的直链线性葡聚糖[2],其水解产物β-1,3-葡寡糖具有多种生理功能,例如诱导分泌白细胞介素[3],触发人类单核细胞产生肿瘤坏死因子α(TNF-α)[4],刺激植物防御反应[5],并抑制人的α-淀粉酶[6],在保健品、食品、农业、医药等领域受到越来越多的关注。 目前酶解法与酸解法是商品热凝胶水解的常用方法[7],酸解法反应条件复杂、难以控制、产率低且易造成环境污染;酶解法反应条件温和、易于控制与操作、产率高,是当下热凝胶水解的研究热点,但是市面上销售的β-1,3-葡聚糖酶是内切酶与外切酶的混合物,不能专一的水解热凝胶且高活性的葡聚糖酶比较少。
作者所在研究室前期研究中利用哈茨木霉GIM3.442 生产β-1,3-葡聚糖酶, 发现其能有效水解热凝胶[8]。 基于此,作者所在研究室建立了土壤杆菌-哈茨木霉及毕赤酵母-土壤杆菌[9-10]两种共培养体系,可直接获得热凝胶寡糖。 但是仍然存在很多不足:1)哈茨木霉在产生内切酶的同时还会产生外切酶,且外切酶占总酶量的70%~80%[11],其水解产物主要是葡萄糖;2)哈茨木霉对剪切力敏感且会在一定程度上抑制土壤杆菌的生长且不易放大[10];3)毕赤酵母中表达出来的酶活比较低,远远达不到我们的需求,有待于进一步提高。近年来,定向进化越来越受到学者们的关注,酶的定向进化是一种蛋白质的非合理设计, 可人为创造特殊的进化条件,模拟自然进化机制,在体外改造基因,从构建的突变库中定向选择出符合目的的突变酶[12]。 由于操作简单、快捷且有效,易错PCR 成为定向进化最为常用的手段之一[13-14]。 有关定向进化内切葡聚糖酶的报道比较多[15],虽然酶活性提高方面也有报道[16-18],但主要集中在pH 与热稳定性等方面的研究[19-21]。作者通过定向进化策略对来源哈茨木霉GIM3.442 的内切β-1,3-葡 聚 糖 酶 基 因 (GenBank Accession:X84085.1)进行定向进化,通过高通量筛选手段获得了高酶活的突变酵母菌株,并将获得的突变体酶与亲本酶蛋白质纯化,进行了部分酶学性质的研究。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 菌株与载体见表1。
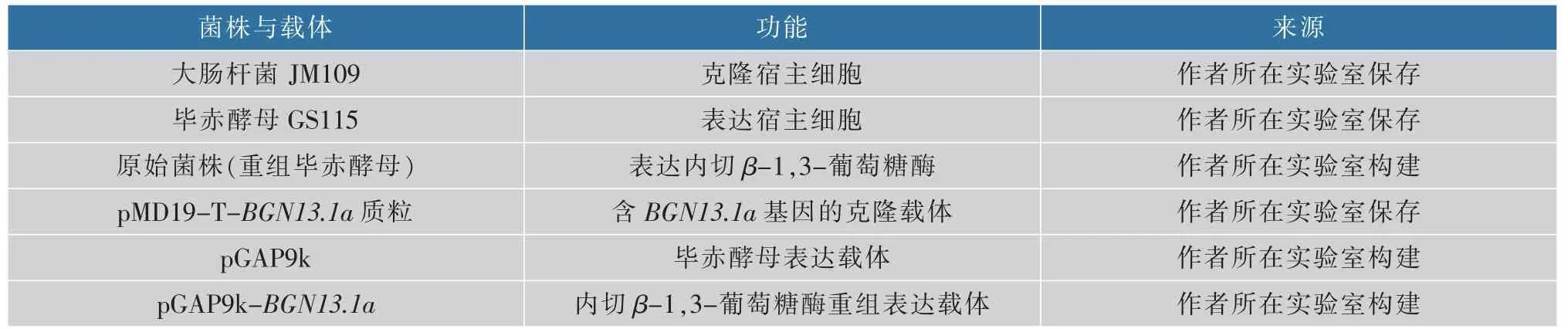
表1 菌株与载体Table 1 Strains and vectors
1.1.2 酶和试剂 限制性内切酶EcoR I、Not I、Sal I、Taq Premix、DNA maker、DNA 连接酶(Solution I) 、Diversify PCR Random Mutagenesis Kit 和DNA 纯化试剂盒:购自TaKaRa 生物公司(大连);Ezup 柱式酵母基因组DNA 抽提试剂盒、 质粒小量抽提试剂盒、氨苄青霉素:购自上海生工有限公司;DNA 胶回收试剂盒: 购自上海康宁公司; 薄层层析板(Thin layer chromatography,TLC Silica gel 60): 购自北京绿百草科技有限公司;其他生物与化学试剂:均为国产或进口试剂。
1.1.3 培养基 平板培养基(组分g/L):葡萄糖20,酵母粉10,蛋白胨20,琼脂20。
种子培养基(组分g/L):葡萄糖20,酵母粉10,蛋白胨20。
发酵培养基(组分g/L):甘油20,酵母粉10,蛋白胨20,YNB 13.4, 生物素4×10-4, 琼脂20;100 mmol/L 磷酸钾缓冲液(pH 6)。
双层鉴别培养基: 下层为甘油20 g/L, 酵母粉10 g/L,蛋白胨20 g/L,YNB13.4 g/L,生物素4×10-4g/L,琼脂20 g/L,100 mmol/L 磷酸钾缓冲液(pH 6);上层为2 g/L 的热凝胶悬浮液1 mL,琼脂0.04 g/L,100 mmol/L 乙酸缓冲液 (pH 5.5) 5 mL,1 g/L 的刚果红5 mL,1 mol/L NaCl 5 mL。
1.2 方法
1.2.1 易错PCR 扩增及突变体库构建 根据已知的哈茨木霉(T. harzianum)GIM3.442 的内切β-1,3-葡聚糖酶基因(GenBank Accession:X84085.1)设计引物, 同时在上下游引物中引入EcoR I 和Not I 两个酶切位点,引物见表2,由上海生工合成,PCR 扩增出的片段为2 340 bp。

表2 引物序列Table 2 Primer sequence
以质粒pMD19-T-BGN13.1a 为模板,易错PCR扩增体系中各种试剂的添加量按照Diversify PCR RandomMutagenesisKit 的说明书进行,体系为50 μL,见表3。PCR 程序为:94 ℃预变性30 s;接着运行以下25 个循环(94 ℃变性30 s,68 ℃延伸150 s);然后68 ℃再延伸60 s,最后4 ℃保温。
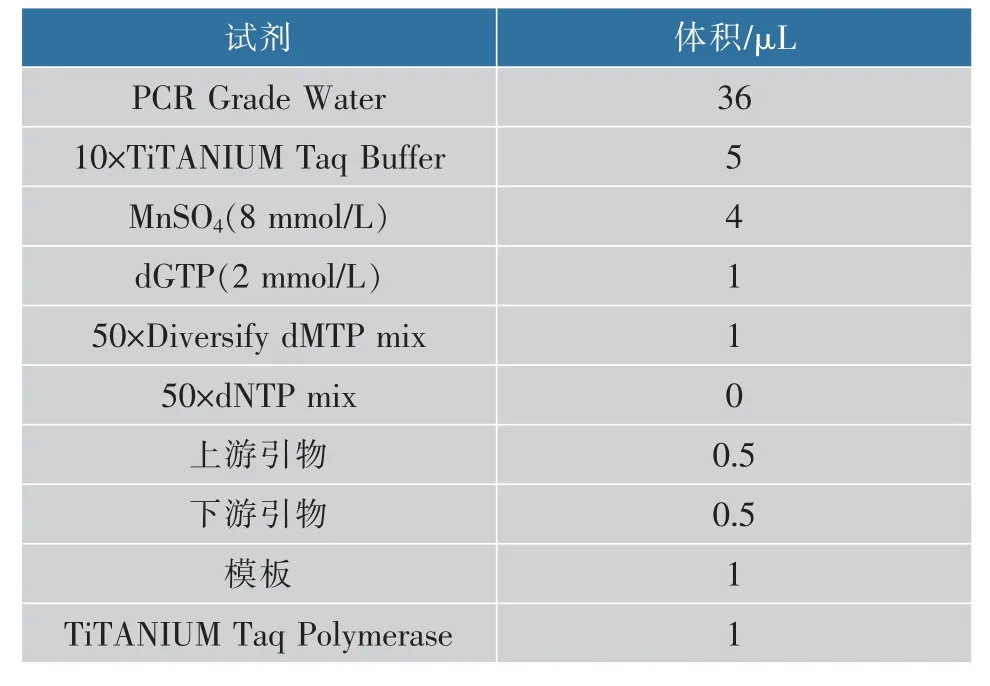
表3 易错PCR 扩增体系Table 3 Error-prone PCR amplification system
易错PCR 产物经1 g/dL 的琼脂糖凝胶电泳后,纯化回收目的条带。纯化后的易错PCR 产物与表达载体pGAP9k 经EcoR I 和Not I 于37 ℃双酶切后进行胶回收,将胶回收后的目标条带与表达载体按照比例混合后加入DNA 连接酶连接过夜, 转化至大肠杆菌JM109 感受态细胞中并涂布于LB (含有100 μg/mL 的Amp) 平板上(表面涂有IPTG 和XGal),37 ℃培养过夜,筛选阳性转化子,得到大量渗入不同突变的重组质粒。 提取这些重组质粒并经Sal I 线性化后, 于2.5 kV 电转入感受态P. pastoris GS115 中,涂布于MD 平板,30 ℃培养2~3 d,构建内切β-1,3-葡聚糖酶基因突变体库。
1.2.2 阳性转化子的筛选与鉴定 将MD 平板上长出的转化子点种到YPD 平板上,并同时复制到一块BMGY 平板上。 YPD 平板于30 ℃培养2~3 d,转移到YPD 液体培养基中,提取基因组,进行PCR 验证。BMGY 平板于30 ℃培养3~4 d,取5 mL 热凝胶半固体 (2 g/dL 热凝胶悬浮液1 mL,0.8 g/dL 琼脂,5 mL 100 mmol/L 的醋酸盐缓冲液) 覆盖在上面,待凝固后,用封口膜封闭好,将其置于50 ℃保温2 h,加入0.1%的刚果红染色,用刚果红染色法筛选阳性转化子。
挑取比值大且含有目的基因的转化子于30 ℃、200 r/min 进行摇瓶发酵,培养4 d,同时将原始菌株(重组毕赤酵母) 活化并在30 ℃、200 r/min摇瓶发酵4 d。 将发酵液于4 ℃、8 000 r/min 离心10 min,取上清液作为粗酶液测定酶活力,并在相同的操作下测定原始菌株与突变菌株的酶活,重复测定3 次,每次3 个平行,统计数据。
酶活测定参照文献[7]略做改动,酶反应体系为:1.5 mL 预处理的2 g/dL 的热凝胶悬浮液、2.5 mL pH 5.5 的0.1 mol/L 的醋酸盐缓冲液,1 mL 酶液,50 ℃反应2 h,沸水浴10 min,参照文献[22]对蒽酮比色法稍作改动,并测定水解液中的总糖质量浓度。 1 个酶活力单位定义为在上述反应条件下每分钟释放1 μmol 还原糖(以葡萄糖为标准)所需要的酶量。
1.2.3 突变株遗传稳定性验证 将获得的突变菌株K827 进行5 次传代,经过摇瓶(100 mL/500 mL)在30 ℃、200 r/min 的条件下培养4 d, 测定其各代的内切酶的酶活、生物量、发酵结束后的pH 等来验证菌种的产酶能力是否稳定。
1.2.4 突变株K827 的SDS-PAGE 分析 将突变株K827 与原始菌株进行摇瓶发酵培养4 d,按测酶活方法获得粗酶液,进行蛋白质纯化,将经过纯化的酶液与4×SDS-PAGE loading buffer 混匀,沸水浴处理10 min,冷却,离心后取20 μL 上清液进行SDSPAGE 凝胶电泳分析,操作过程见文献[23]。
1.2.5 突变酶K827 水解产物分析 将突变体酶K827 的发酵液于4 ℃、8 000 r/min 离心10 min,取上清得到粗酶液,按照文献[7]配制酶反应体系:1.5 mL 预处理的2 g/dL 的热凝胶悬浮液、2.5 mL pH 5.5 的0.1 mol/L 的醋酸盐缓冲液,1 mL 酶液,置于50 ℃水浴锅,保温2 h,煮沸10 min, 8 000 r/min离心10 min,取上清液(水解产物)测定酶活,重复测定3 次,将水解液进行TLC 薄层色谱及MALDITOF MS 分析。
1.2.6 突变株K827 内切-β-1,3-葡聚糖酶酶学性质分析
1) 突变株K827 内切β-1,3-葡聚糖酶的最适反应pH 与酸碱稳定性 在2.5 mL 0.1 mol/L 不同pH 的醋酸盐缓冲液中分别加入1.5 mL 预处理的2 g/dL 的热凝胶悬浮液和1 mL 适当稀释的酶液,置于50 ℃水浴锅,保温2 h,煮沸10 min,8 000 r/min离心10 min,取上清液,测定酶活,确定酶的最适反应pH。 将原酶液置于不同pH 值的缓冲液中,4 ℃保温12 h, 然后在其最适pH 值下配制酶反应体系,置于50 ℃水浴锅,保温2 h,煮沸10 min,8 000 r/min 离心10 min,取上清液,测定剩余酶活,从而确定pH 稳定性。
2) 突变株K827 内切β-1,3-葡聚糖酶的最适反应温度与热稳定性 首先,在最适反应pH 条件下配制酶反应体系:1.5 mL 预处理的2 g/dL 的热凝胶悬浮液,2.5 mL 0.1 mol/L 的醋酸盐缓冲液,1 mL适当稀释的酶液,分别在不同的温度梯度下保温2 h,煮沸10 min,8 000 r/min 离心10 min,取上清液,然后测定酶活,确定酶的最适反应温度。 然后再将适当稀释的酶液置于不同的温度梯度下,保温100 min,每隔20 min 取样,再配制酶反应体系,在50 ℃水浴锅中保温2 h,煮沸10 min,8 000 r/min 离心10 min,取上清液,测定剩余酶活,从而测定其热稳定性。
2 结果与分析
2.1 易错PCR 与突变体库的构建
目的基因BGN13.1a 的大小为2 340 bp, 以质粒pMD19-T-BGN13.1a 为模板,按照Diversify PCR Random Mutagenesis Kit 的说明书进行易错PCR,见图1。 在2 400 bp 附近有条带出现(泳道1),说明目的基因已经成功渗入突变。
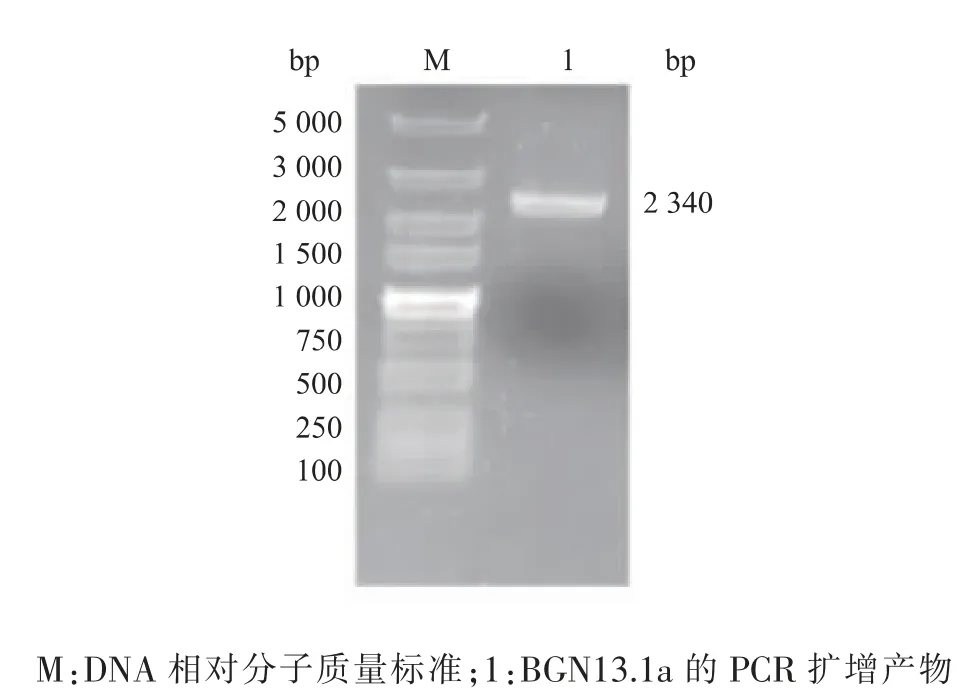
图1 易错PCR 的电泳图Fig. 1 Patterns of error-prone PCR
经Amp 抗性及蓝白斑双重筛选后,将长出的白色菌落全部接种到一个摇瓶里培养并提取重组质粒, 线性化电转入感受态P. pastoris GS115 酵母菌中,MD 平板上长出的转化子即为构建好的目的基因突变体库。
2.2 阳性转化子的筛选与鉴定
提取阳性转化子的基因组, 进行PCR 扩增,结果显示每一个阳性转化子均能扩增出2 340 bp 的条带,说明渗入突变的内切β-1,3-葡聚糖酶基因已经成功导入到毕赤酵母中。对BMGY 平板长出的菌落, 进行底物覆盖2 h 及质量分数0.1% 刚果红染色1 h,再用1 mol/L NaCl 脱色多次。 若转化子具有酶活,那么菌落周围会出现明显的透明光圈,透明光圈与菌落的比值越大,说明转化子的酶活越高[24]。最终筛选得到一株高酶活的突变株,命名为K827。
挑取突变株K827,进行扩大培养,同时将原始菌株进行培养,离心取上清液,测定两者的酶活,突变株K827 水解热凝胶的比酶活为(82.42±0.75)U/mg,比原始菌株(65.84±0.62) U/mg 提高了25%。
2.3 突变菌株遗传稳定性验证
为检测最终获得的突变株K827 产BGN13.1a的遗传稳定性,连续培养了5 代,测定各代发酵参数,结果见表4。

表4 突变株遗传稳定性验证Table 4 Genetic stability verification of mutant strains
从表4 可以看出, 各代菌株发酵结束时的pH在6.38 左右,生物量在8.00 左右,酶活在62.22 U/mL左右。 发现该突变株在每一次传代培养结束时的酶活、生物量及发酵结束后的pH 均未发生明显变化,说明该突变株产酶性能稳定, 各项参数较为稳定,具有良好的遗传稳定性。
2.4 突变酶K827 的SDS-PAGE 分析
软件Expasy 预测目的基因表达的蛋白质相对分子质量在70 000 左右,见图2。 泳道M、1、2 依次为蛋白质Marker(High)、纯化的野生型BGN13.1a、纯化的突变株K827, 其中泳道1、2 在70 000 附近都出现条带,说明表达的目的蛋白质与预期相符。

图2 野生型和突变体酶K827 的SDS-PAGE 电泳图Fig.2 SDS-PAGE analysis of the wild-type and the mutant enzyme K827
2.5 突变酶K827 水解产物分析
通过TLC 薄层色谱及MALDI-TOF MS 分析突变体酶K827 水解热凝胶的产物,结果见图3。由图3 可知, 目标糖的质荷比 (m/z) 分别为365.146、527.149、689.177、851.249、1 013.324、1 175.393、1 337.46、1 499.51、1 661.558, 显示突变体酶K827 水解热凝胶产物的聚合度主要为DP=2~10,这与TLC 薄层色谱结果一致。 同时通过TLC薄层色谱与Image J 软件定量得出DP=2~10 的寡糖产量分别为0.348、0.519、0.419、0.079、0.017、0.078、0.015、0.011、0.006 g/L,共1.492 g/L。
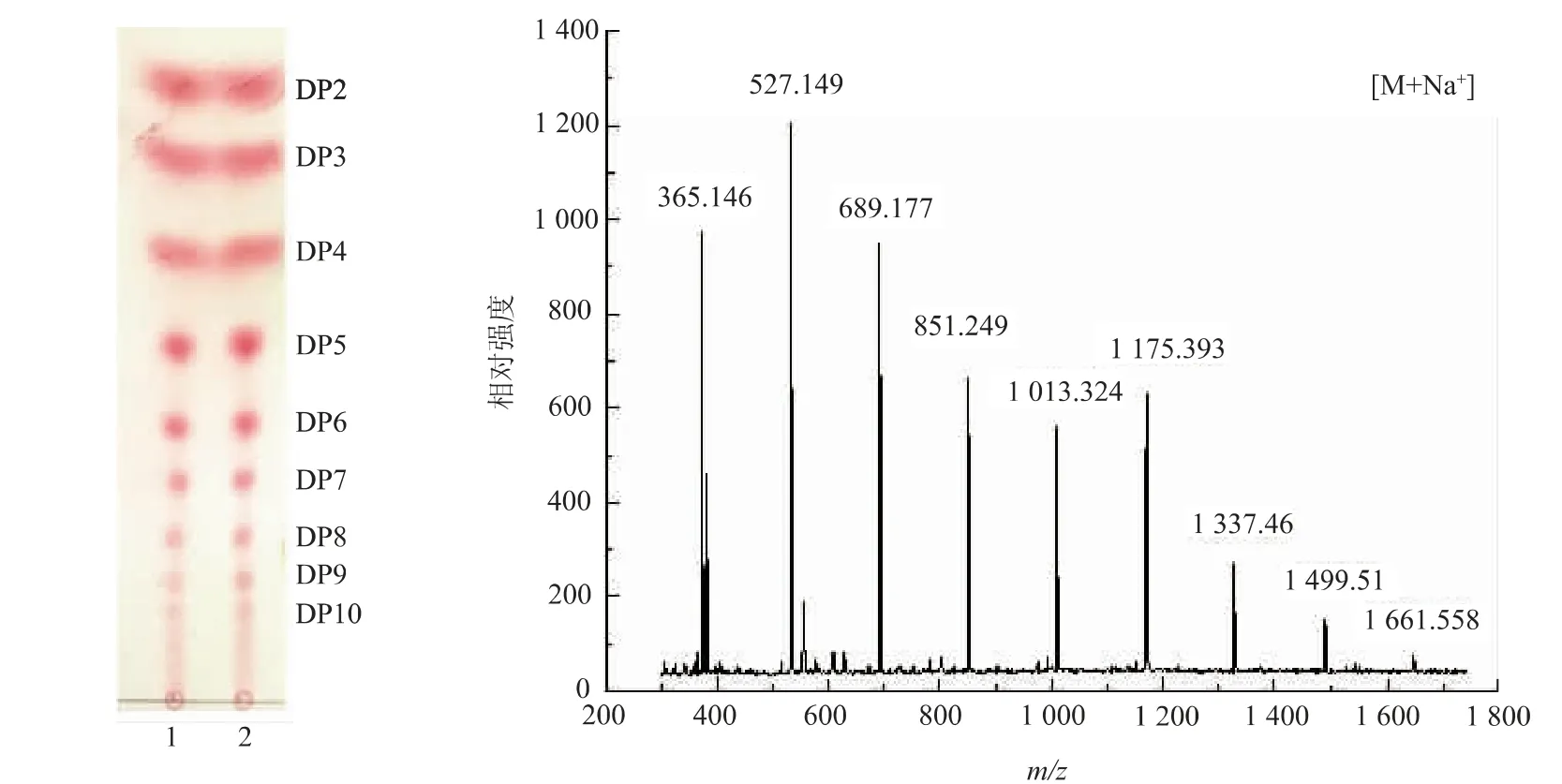
图3 突变体酶K827 水解产物薄层层析和MALDI-TOF MS 分析结果Fig. 3 TLC and MALDI-TOF MS analysis of mutant enzyme K827 hydrolysate
2.6 突变株K827 内切-β-1,3-葡聚糖酶酶学性质分析
2.6.1 突变株K827 酶的最适反应pH 将酶反应的0.1 mol/L 的醋酸盐缓冲液的pH 配成4.5~7.5(梯度为0.5),按照文献[7]配制酶反应体系,置于50 ℃水浴锅,保温2 h,煮沸10 min,测定酶活,重复测定3 次,定义最高酶活为100%。结果表明:突变体酶与原始酶的最适反应pH 值为5.5, 突变体酶在pH 4.5~6 的酶活力可达到80%以上, 而原始酶的酶活力在pH 5~5.5 才能达到80%以上,结果见图4(a),由此可见,与原始酶相比,突变体酶在更宽的pH 范围,都能保持较高的酶活性。
2.6.2 突变株K827 酶的pH 稳定性 将酶置于pH 4.5~7.5(梯度为0.5) 0.1 mol/L 的醋酸盐缓冲液中,于4 ℃处理12 h,按照文献[7] 配制酶反应体系,置于50 ℃水浴锅,保温2 h,煮沸10 min,测定酶活,重复测定3 次,定义最高酶活为100%。 结果表明,突变体酶与原始酶在pH 4.5~6.5 的缓冲液中4 ℃处理12 h, 突变体酶在pH 5~6 剩余酶活力保持90%以上, 而原始酶只能在最适pH 附近剩余酶活达到90%以上,结果见图4(b),由此可见,与原始酶相比,突变体酶的pH 稳定性有明显提高。

图4 突变体酶的最适p H 与pH 稳定性Fig. 4 Optimal pH and pH stability of the mutant endonuclease
2.6.3 突变株K827 酶的最适反应温度 将按照文献[7]配制的酶反应体系置于30~70 ℃(梯度为5 ℃)的水浴锅中,保温2 h,煮沸10 min,测定酶活,定义最高酶活为100%,见图5(a)。结果表明,突变体酶与原始酶的最适反应温度为50 ℃, 突变体酶在40~55 ℃范围内酶活可达到60%以上,而原始酶在50 ℃附近才能达到60%以上。 由此可见,突变体酶在更宽的温度范围内都能保持较高的酶活性。
2.6.4 突变株K827 酶的热稳定性 将突变体酶与原始酶的酶液用pH 5.5 的缓冲液稀释后,置于30~70 ℃(梯度为10 ℃)中保温,每隔20 min 取样,取完后立即冰浴,并按照文献[7]配制酶反应体系,于50 ℃测定剩余酶活, 定义最高酶活为100%(图5(b)、(c)、(d)、(e)、(f))。 结果表明,突变体酶在30、40 ℃处理100 min 后,剩余酶活仍可保持在60%以上,而原始酶在30~70 ℃范围内处理100 min,其剩余酶活都不能达到60%。 由此可见,与原始酶相比,突变体酶的热稳定性有明显提高。
3 结 语
作者通过对野生型酶BGN13.1a 进行基因突变,获得了最佳突变体酶K827,酶活较原始菌提高了25%。 在获得热凝胶寡糖的水解体系中,相比较野生型酶, 同等条件下的突变体酶K827 能够提高热凝胶的水解效率、缩短热凝胶水解时间,寡糖产量为1.492 g/L(聚合度2~10),该突变菌株具有较强的开发和应用潜力。

图5 突变体酶的最适温度及温度稳定性Fig. 5 Optimal temperature and temperature stabilityof mutant endonuclease
