鲐鱼冷海水贮藏期间特定腐败菌的鉴定
2021-05-15郑振霄戴志远
郑振霄,戴志远*
(1 浙江工商大学海洋食品研究院 杭州310012 2 浙江工商大学 浙江省水产品加工技术研究联合重点实验室 杭州310012)3 浙江工商大学 海洋食品精深加工关键技术省部共建协同创新中心 杭州310012)
鱼类在捕获后很容易发生腐败变质,细菌活动是引起鱼类腐败变质的主要因素。新鲜渔获物中的细菌组成多样,包含渔获物所生长环境中的细菌和鱼体自带的细菌。然而,在鱼体捕获后,贮藏条件(如温度、氧气、盐分等)对鱼体的菌相组成产生重要影响,能够适应贮藏条件的细菌大量繁殖,不适应的细菌被淘汰。这些适应环境的细菌中,又有一少部分可以降解鱼体的营养物质,代谢产生三甲胺和硫化氢,这些物质会产生强烈的腐败臭味,具有很强的致腐能力[1-6]。20世纪90年代,丹麦国科学家Dalgaard[7]在总结前人的诸多研究的基础上,将这一少部分细菌定义为特定腐败菌,这一概念对于后来人们研究水产的腐败,具有重要意义。
鲐鱼是一种中、上层鱼类,随着我国海洋渔业资源分布的变迁,鲐鱼等中、上层鱼类渔业资源的地位不断攀升[8]。鲐鱼的渔汛一般在夏季,温度高,渔获量大,此外,鲐鱼属于鲭科鱼类,渔获不及时处理,易发生组胺积累,导致鲐鱼比其它水产更容易发生腐败劣变。因此,鲐鱼捕获后应立即采取保鲜措施,否则难以保质。鉴于渔船上的实际条件,能够有效执行的保鲜方法有冰鲜、冻藏保鲜和冷海水保鲜。冰鲜是最常用的保鲜方法,它可以较大程度地保持鲐鱼肌肉的原始特征,从而保持较好的口感,然而冰鲜的处理量较少,需要占用较大的空间;冻藏是另一种常用的保鲜方法,冻藏的最大特点就是保鲜时间长,然而冻藏不仅能耗高,而且冻藏的样品解冻后不能完全保持原有特征,且冻藏期间的氧化劣变也是不可忽视的。相比于上述两者,冷海水保鲜采用的冷却介质为液体,液体的传热效率远远大于空气,这样鱼体的温度能够在较短时间内降下来,利于后期处理。此外,冷海水保鲜的处理量大,广泛应用于鲐鱼等中、上层鱼类捕获后的暂存。然而,冷海水保鲜的贮藏环境也与上述两者有很大差别,使贮藏期间微生物种群的变化与前两者有较大差别[9-10],而对于特定腐败菌,它不仅随鱼种的变化而变化,而且还随贮藏环境的变化而变化[7]。
关于鲐鱼不同贮藏过程中的菌相组成及变化的研究报道较多。翁佩芳等[11]采用16S rDNA 测序技术研究鲐鱼在冻藏期间优势菌群的变化,结果表明不动杆菌为贮藏前期的优势菌群,随着贮藏时间的延长,优势细菌种群变为梭杆菌属和希瓦氏菌属,冻藏20 d 的样品中优势种群为能产生组胺的哈夫尼菌属和沙雷菌属。孟璐[12]采用宏基因组测序的方法研究气调包装后,在冷藏条件下鲐鱼中的菌相变化,结果表明鲐鱼在贮藏过程中菌相组成逐渐单一,由初始的以海洋科细菌和盐单胞菌科细菌为主过渡到以希瓦氏菌为主,货架终点的优势微生物为发光杆菌。虽然许多学者对鲐鱼贮藏过程中的微生物变化做过相关研究,但是关于鲐鱼冷海水贮藏期间的菌相变化及特定腐败菌的研究甚少。本文以新鲜鲐鱼为对象,采用机制冷海水对其进行保鲜处理,采用选择性培养基和16S rDNA 序列法分析鲐鱼在贮藏前、中、后期的菌相组成及变化,并通过对腐败样品中优势腐败菌致腐能力的测定,确定鲐鱼冷海水保鲜的特定腐败菌,为鲐鱼的质量安全控制提供参考。
1 材料与方法
1.1 材料、试剂与仪器
新鲜鲐鱼,捕获于中国东海,与渔船工作协商将最后一网捕获的鲐鱼作为试验原料。
假单胞菌(CFC)选择性培养基(Pseudomonas CFC selective agar)、硫代硫酸钠柠檬酸胆盐蔗糖(TCBS) 琼脂 (Thiosulfate citrate bile sucrose agar)、结晶紫中性红胆盐葡萄糖 (VRBGA) 琼脂(Violet red bile glucose agar,VRBGA)、MRS 琼脂(MRS agar)、铁琼脂(Icron agar)、2216E 琼脂(2216E agar)、STAA 琼脂培养基(STAA agar mesium),青岛海博生物科技有限公司;盐酸、氯化钠等常规化学试剂(分析纯级),国药集团(上海)有限公司。
恒温恒湿培养箱,上海一恒科学仪器设备有限公司。
1.2 模拟冷海水保鲜系统的建立
洁净的海水取自中国东海,装入25 L 的不锈钢容器中(下有流水出口),将此容器放置在冷藏室中(设定温度为-1~0 ℃)。新鲜的鲐鱼捕获于中国东海,捕获后立即放入装有冷海水的容器中,6 h 内运到实验室。运至实验室后,挑取规格统一、体形完整的样品放进实验室自制的冷海水系统中(海水与鱼的质量比为7∶3),试验期间每2 d 更换1 次冷海水。贮藏期为12 d,其中第0,3,6,9,12天为取样时间节点,每次取3 条,采集鲐鱼的背部肌肉用于后续试验。
1.3 感官评定
取不同贮藏期的鲐鱼肉样品,参考Manju等[13]的方法对其进行感官评定。感官评价的标准设定为色泽、质地和气味。由6 位经过培训的感官评定人员组成评定小组,对鲐鱼的色泽、质地和气味进行打分,综合评分为1~9 分,1 分品质最差,9分品质最好,4 分作为可接受下限。
1.4 理化分析
挥发性盐基氮的测定参照《食品中挥发性盐基氮的测定》(GB 5009.228-2016)[14]中的方法进行测定;三甲胺的测定参照《食品中三甲胺的测定》(GB 5009.179-2016)[15]中的方法进行测定;组胺的测定参照《食品中生物胺的测定》(GB 5009.208-2016)[16]中的方法进行测定。
1.5 选择性培养基分析菌群群落变化
分别采用不同的选择性培养基,对不同贮藏时间的鲐鱼样品中相关的菌落进行计数,具体培养条件见表1[17-19]。
1.6 菌种鉴定
样品中DNA 的提取方法参照DNA Kit(Omega Bio-tek,Norcross,GA,U.S.A)试剂盒说明书的方法进行。引物采用27F 和1492R 这2 种通用引物,扩增所得产物通过琼脂糖凝胶电泳分析其纯度,测序由上海桑尼生物科技有限公司完成。
1.7 优势腐败菌致腐能力的测定
将从腐败样品中得到的优势腐败菌置于液体培养基中,制成105CFU/mL 的菌悬液,按照鱼肉∶菌悬液=1∶0.1 (g/mL) 的料液比接种到无菌鱼块上,以未接种腐败菌的无菌鱼肉块作为对照,贮藏在0 ℃冰箱中,定期测定样品中的挥发性盐基氮、三甲胺、组胺和菌落总数,以评价优势腐败菌的致腐能力[20-21]。
2 结果与分析
2.1 感官评价
贮藏期间感官评分变化见图1。在整个贮藏期间,感官评分和贮藏时间呈负相关,感官评分的最大值和最小值分别为8.5 分和1.6 分。新鲜鲐鱼体表鳞片有光泽,有透明黏液附着在其表面,肌肉坚实有弹性,感官评价为8.5 分。随着贮藏时间的延长,感官评分不断下降,在贮藏第6 天时,样品出现了显著软化,并伴随破肚现象,评分下降到5分;贮藏到第9 天,破肚样品进一步增多,且散发出特有的腐败臭味,感官评分降低至3.1 分;贮藏结束时,多数样品出现了破肚现象,散发出浓重的腐败恶臭,感官评分低至1.5 分。综上所述,以感官评价为指标,鲐鱼冷海水保鲜的保鲜期为9 d。对比前人研究[10,22-23],冷海水保鲜的货架期与冰鲜贮藏方法较为接近,然而明显短于冻藏。
2.2 菌相变化
选择性培养基是根据某类微生物特殊的营养要求,或者该类微生物的某些特殊化学、物理因素的抗性而设计的,能够选择性区分这一类微生物的培养基。利用选择性培养基,可使混合菌群中的某一类微生物变成优势种群,提高筛选效率。然而,腐败菌的种类繁多,通常使用1 种培养基不能对腐败菌进行全面筛选,通常需要几种选择性培养基一起使用,才能够对整体的腐败菌做到有效筛选[9,24-25]。本文选取了6 种常用于筛选腐败菌的选择性培养基,对鲐鱼冷海水贮藏期间的菌相变化进行分析,不同选择性培养基上菌相变化如图2所示。整体来看,贮藏初期样品在不同选择性培养基上的菌落数都处于较低水平,数值分别为2.54 lg(CFU/g)(VRBGA),2.63 lg (CFU/g)(2216E),2.15 lg (CFU/g)(CFC),1.50 lg (CFU/g)(IA),1.31 lg(CFU/g)(MRS)和1.10 lg(CFU/g)(STAA)。随着贮藏时间的延长,2216E、MRS 和STAA 培养基上的菌落总数呈现先增加后下降的趋势,最大值分别出现在第6 天[4.60 lg (CFU/g)]、第9 天[(2.78 lg(CFU/g)]和 第3 天[(3.61 lg (CFU/g)]。VRBGA、CFC 和IA 培养基上的菌落总数在整个贮藏过程中始终处于增加趋势,最大值为5.50,4.81,6.17 lg(CFU/g),分别是初始值的2.17,2.23,4.11 倍。结果表明,2216E、MRS 和STAA 培养基上的种群在贮藏过程中不能适应所处的环境,因而逐渐被淘汰,而VRBGA、CFC 和IA 培养基上的种群能够适应冷海水贮藏环境,逐渐成为优势种群。
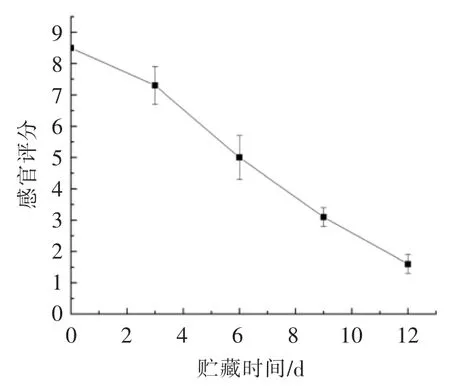
图1 感官评分的变化Fig.1 Changes in sensory score
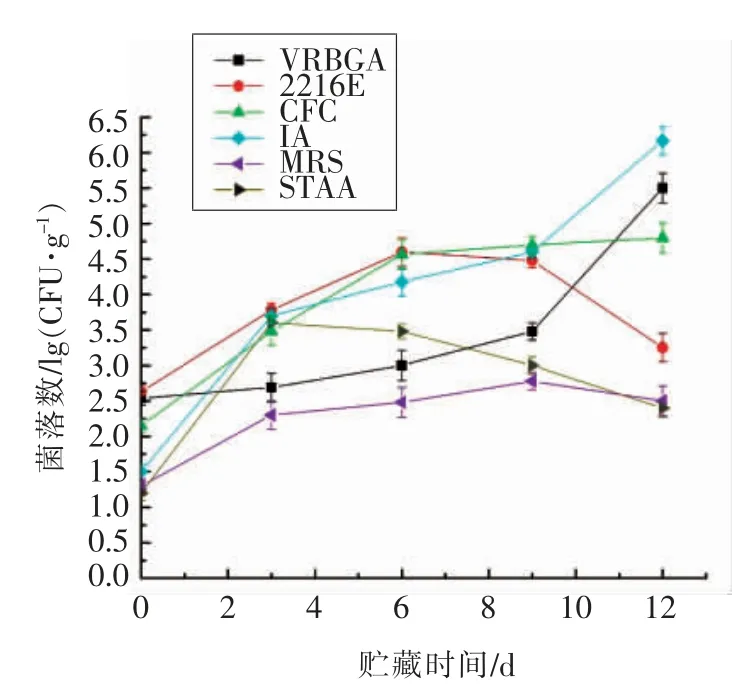
图2 菌群组成的变化Fig.2 Changes in the composition of the flora
2.3 菌种鉴定
采用16S rDNA 序列分析法对鲐鱼贮藏初期(0 d)、中期(6 d)和末期(12 d)的样品中的代表性微生物进行菌种鉴定,鉴定结果如表2所示。贮藏初期的样品中共纯化得到12 株具有代表性菌株,它们分别来自3 种选择性培养基,VRBGA(3 株),2216E(5 株),CFC(4 株)。其中,6 株被鉴定为嗜冷杆 菌 (Psychrobacter),3 株被鉴定为假单胞菌(Pseudomonas),2 株被鉴定为伽马变形菌(Oceanisphaera sp.),1 株被鉴定为类芽孢杆菌(Paenibacillus sp.),嗜冷杆菌占多数,说明嗜冷杆菌是鲐鱼贮藏初期的优势微生物。贮藏中期的样品中共纯化得到30 株具有代表性的菌株,分别为CFC(11 株),VRBGA(6 株),IA(5 株),2216E(5株),STAA(2 株),MRS(1 株)。在30 株菌中,有12株被鉴定为假单胞菌,占绝大多数,说明假单胞菌是贮藏中期的优势微生物,其次为希瓦氏菌(Shewanella,6 株)和嗜冷杆菌(6 株)。此外,还有2 株被鉴定为肠杆菌(Enterobacter),短杆菌(Brevibacterium sp.)、伽马变形菌、发光杆菌(Photobacterium phosphoreum)和稳杆菌(Empedobacter)各有1株。贮藏末期的样品中共纯化得到41 株具有代表性菌株,分别为IA(12 株),VRBGA(11 株),CFC(8 株),2216E(4 株),MRS(3 株),STAA(3 株)。在这41 株菌株中,数量较多的有希瓦氏菌(14 株)、肠杆菌(6 株)和柠檬酸杆菌(Citrobacter,6 株),此外还有4 株被鉴定为假单胞菌,2 株被鉴定为葡萄球菌 (Staphylococcus saprophyticus),2 株被鉴定为热死环丝菌 (Brochothrix thermosphacta),2株嗜冷杆菌,变形杆菌(Proteus vulgaris)、伽马变形杆菌、发光杆菌、勒克斯菌(Leclercia adecarboxylata)、稳杆菌各1 株。希瓦氏菌是一种嗜冷菌,它可以还原水产中的氧化三甲胺为三甲胺,此外还可以将含硫的氨基酸降解成挥发性的硫化氢。三甲胺和硫化氢都是具有鱼腥恶臭的代谢产物,与水产腐败臭味密切相关,许多学者将希瓦氏菌认定为水产品在低温贮藏条件下的主要腐败微生物[26-28]。肠杆菌是一类常见的兼性厌氧革兰氏阴性菌,某些肠杆菌具有致病性,尤其对抵抗力差的人群,更容易受其感染而患病[29]。柠檬酸杆菌可以分泌胞外酶,这些酶具有降解组氨酸成组胺的能力,非常容易导致人体出现过敏反应[30-31]。希瓦氏菌、肠杆菌和柠檬酸杆菌所占的比例分别为34.15%,14.63%和14.63%,其它菌株的占比均小于10%。这说明希瓦氏菌、肠杆菌和柠檬酸杆菌是腐败样品中的优势腐败菌。为研究这3 类细菌在鲐鱼腐败过程中所发挥的作用,后续试验以这3种细菌的不同组合接种于灭菌鱼块上,并以没接种细菌的灭菌鱼块为对照,以鱼块中挥发性盐基氮、三甲胺、组胺和菌落总数为指标,研究这3 种细菌在鲐鱼腐败过程中的致腐能力。具体分组为第1 组、第2 组、第3 组分别接种希瓦氏菌、肠杆菌、柠檬酸杆菌,第4 组、第5 组、第6 组分别接种等量的希瓦氏菌和肠杆菌、希瓦氏菌和柠檬酸杆菌、肠杆菌和柠檬酸杆菌,第7 组接种等量的希瓦氏菌、肠杆菌和柠檬酸杆菌,第8 组为空白对照组。
2.4 优势腐败菌致腐能力的测定
2.4.1 TVB-N 挥发性盐基氮是由细菌分泌的内源性蛋白酶降解样品中蛋白所产生的一种含氮碱性物质,是评判水产腐败的重要指标[32]。新鲜的水产中挥发性盐基氮的含量很低,在腐败进程中,其含量会不断上升,30 mg/100 g 通常被认定为可食用上限[32-33]。接种不同腐败菌的鱼块中的挥发性盐基氮随时间的变化如图3a 所示。挥发性盐基氮的初始值为10.58 mg/100 g,表明原料新鲜,品质好。各试验组中的TVB-N 含量随时间延长逐渐增加,比较第1,2,3 组,第1 组的增量最多,贮藏结束时TVB-N 值为50.21 mg/100 g,TVB-N 含量在第2组和第3 组中的增加速度很慢,贮藏结束也未超过30 mg/100 g。比较第4,5,6,7 组,第6 组的TVB-N 含量显著低于其它3 组,贮藏结束时TVB-N 含量也没有超过可接受上限,其它3 组在第0~6 天时,增加缓慢,之后增加速度加快,在第9 天超过了30 mg/100 g。整个贮藏过程中对照组的TVB-N 含量最低,贮藏结束仍未超过可接受限量。从结果可以看出,接种了希瓦氏菌的样品中挥发性盐基氮的增加显著高于未接种希瓦氏菌的样品组,结果表明希瓦氏菌对于挥发性盐基氮的贡献大于肠杆菌和柠檬酸杆菌。
2.4.2 三甲胺 三甲胺是评价海水鱼鲜度的另一项常用指标,它的产生是由于微生物代谢产生的还原酶对氧化三甲胺的还原作用。三甲胺在不同鱼中的限量值略有不同,在多数的多脂性鱼类中的限量标准不超过5 mg/100 g,其它鱼类的标准通常介于5~10 mg/100 g 之间[34]。接种不同菌株的鱼块中三甲胺的含量随贮藏时间的变化如图3b 所示。比较第1,2,3 组,第1 组中三甲胺的含量最高,贮藏结束后达20.15 mg/100 g,远远超过了限量值,第2 组和第3 组的三甲胺含量增加缓慢,贮藏结束时分别为9.11,5.59 mg/100 g。比较第4,5,6,7 组,第6 组的增加量要显著低于其它3 组,贮藏结束为8.51 mg/100 g,其它组的三甲胺含量均超过了20 mg/100 g。
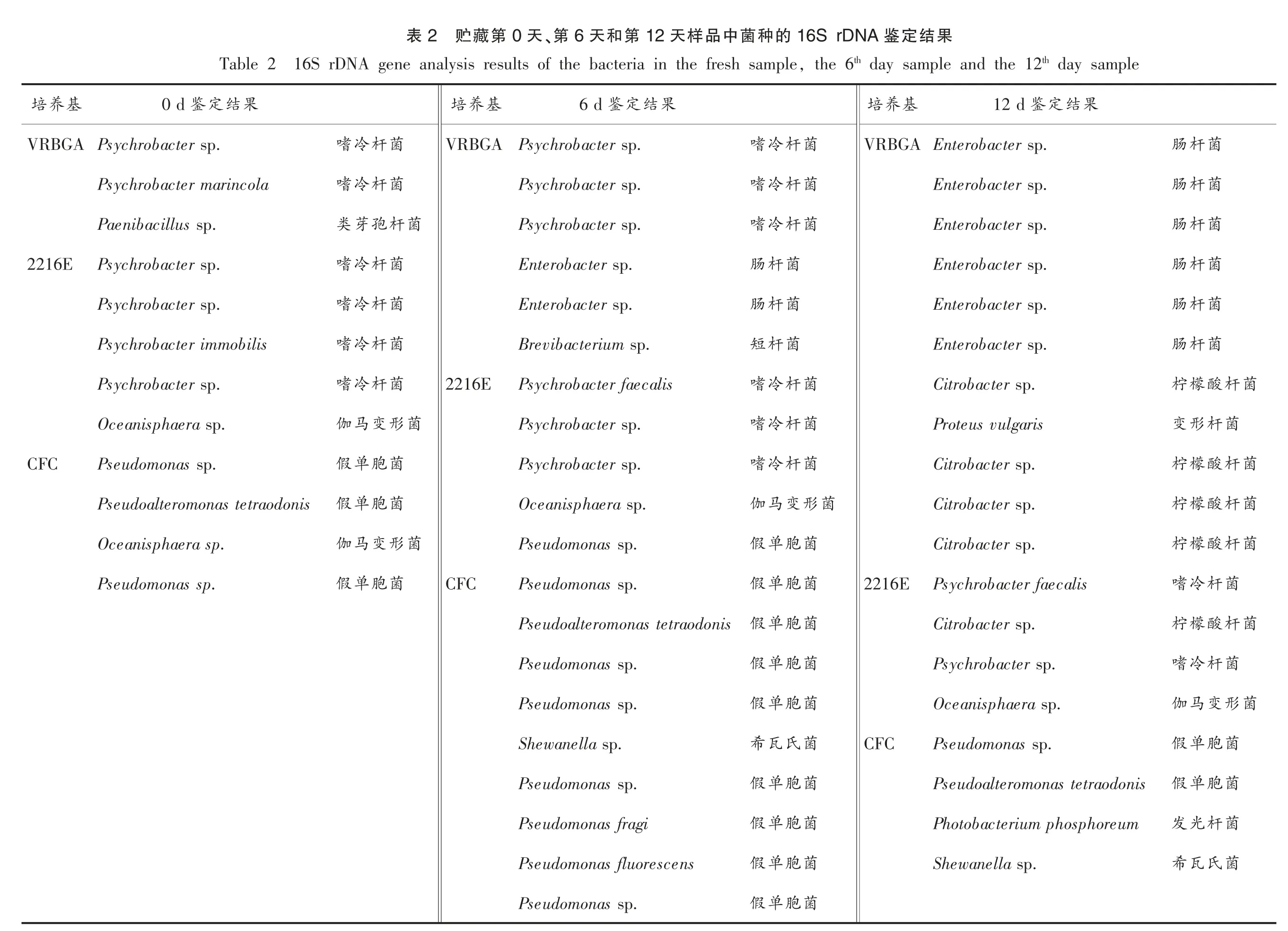
菌菌菌菌菌菌杆菌杆杆杆菌杆菌形菌菌菌菌菌杆肠菌杆肠菌杆肠菌杆肠菌杆肠菌杆肠酸檬柠杆形变酸檬柠酸檬柠酸檬柠杆冷嗜酸檬柠杆冷嗜变马伽胞单假胞单假杆光发氏瓦希果结定鉴16S rDNA的种菌中品样天12第和天6、第天0第藏贮2表th day sample th day sample and the 12,the 6 16S rDNA gene analysis results of the bacteria in the fresh sample Table 2 果结定鉴12 d基养培果结定鉴6 d基养培果结定鉴0 d基养培VRBGA Enterobacter sp.菌杆冷嗜VRBGA Psychrobacter sp.菌杆冷嗜VRBGA Psychrobacter sp.Enterobacter sp.菌杆冷嗜Psychrobacter sp.菌杆冷嗜Psychrobacter marincola Enterobacter sp.菌杆冷嗜Psychrobacter sp.菌杆孢芽类Paenibacillus sp.Enterobacter sp.菌杆肠Enterobacter sp.菌杆冷嗜Psychrobacter sp.2216E Enterobacter sp.菌杆肠Enterobacter sp.菌杆冷嗜Psychrobacter sp.Enterobacter sp.菌杆短Brevibacterium sp.菌杆冷嗜Psychrobacter immobilis Citrobacter sp.菌杆冷嗜Psychrobacter faecalis 2216E菌杆冷嗜Psychrobacter sp.Proteus vulgaris菌杆冷嗜Psychrobacter sp.菌形变马伽Oceanisphaera sp.Citrobacter sp.菌杆冷嗜Psychrobacter sp.菌胞单假Pseudomonas sp.CFC Citrobacter sp.菌形变马伽Oceanisphaera sp.菌胞单假Pseudoalteromonas tetraodonis Citrobacter sp.菌胞单假Pseudomonas sp.菌形变马伽Oceanisphaera sp.Psychrobacter faecalis 2216E菌胞单假Pseudomonas sp.CFC菌胞单假Pseudomonas sp.Citrobacter sp.菌胞单假Pseudoalteromonas tetraodonis Psychrobacter sp.菌胞单假Pseudomonas sp.Oceanisphaera sp.菌胞单假Pseudomonas sp.Pseudomonas sp.CFC菌氏瓦希Shewanella sp.Pseudoalteromonas tetraodonis菌胞单假Pseudomonas sp.Photobacterium phosphoreum菌胞单假Pseudomonas fragi Shewanella sp.菌胞单假Pseudomonas fluorescens菌胞单假Pseudomonas sp.
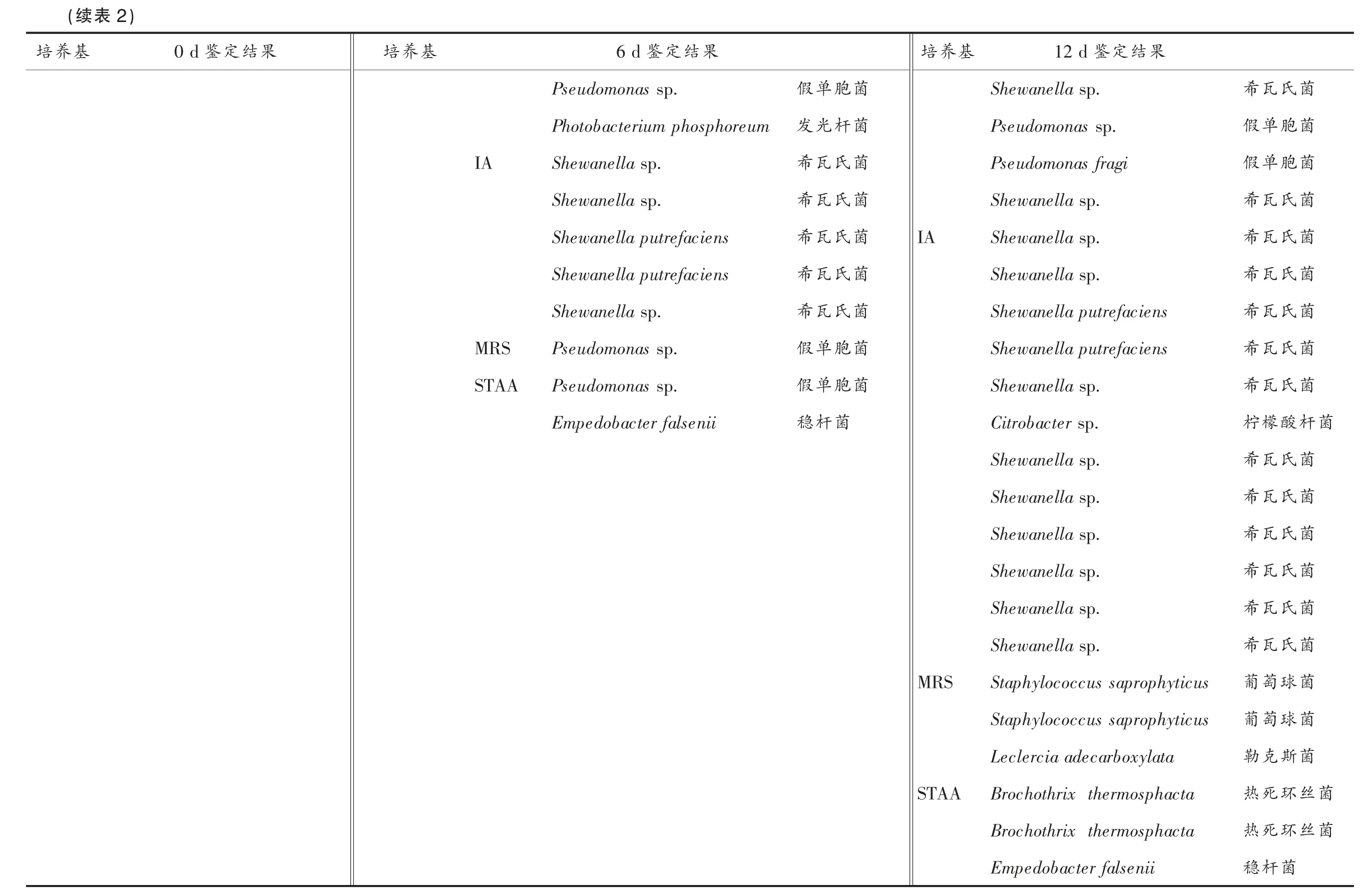
菌菌菌菌菌菌菌菌菌菌菌菌杆菌菌菌菌菌菌菌菌菌丝丝氏瓦希胞单假胞单假氏瓦希氏瓦希氏瓦希氏瓦希氏瓦希氏瓦希酸檬柠氏瓦希氏瓦希氏瓦希氏瓦希氏瓦希氏瓦希球萄葡球萄葡斯克勒环死热环死热菌杆稳果结定鉴12 d Shewanella sp.Pseudomonas sp.Pseudomonas fragi Shewanella sp.Shewanella sp.Shewanella sp.Shewanella putrefaciens Shewanella putrefaciens Shewanella sp.Citrobacter sp.Shewanella sp.Shewanella sp.Shewanella sp.Shewanella sp.Shewanella sp.Shewanella sp.Staphylococcus saprophyticus Staphylococcus saprophyticus Leclercia adecarboxylata Brochothrix thermosphacta Brochothrix thermosphacta Empedobacter falsenii基养培IA MRS STAA菌菌菌菌菌菌菌菌菌胞单假杆光发氏瓦希氏瓦希氏瓦希氏瓦希氏瓦希胞单假胞单假菌杆稳果结定鉴6 d Pseudomonas sp.Photobacterium phosphoreum Shewanella sp.Shewanella sp.Shewanella putrefaciens Shewanella putrefaciens Shewanella sp.Pseudomonas sp.Pseudomonas sp.Empedobacter falsenii IA MRS STAA基养培果结定鉴0 d)2表(续基养培
2.4.3 组胺 组胺是一种化学性毒素,是鲭科鱼类发生中毒事件的主要因素。组胺在新鲜的鱼体中含量很低,然而如果在贮藏过程中操作不当,产组胺微生物会快速繁殖,鱼体内的组氨酸会在组氨酸脱羧酶的作用下降解成组胺[35]。鲐鱼是典型的鲭科鱼类,其肌肉中组氨酸的含量处于较高水平,由于误食造成的组胺中毒事件常发。因此,对于鲐鱼体内组胺积累情况的监测十分必要。接种不同菌种鲐鱼块中组胺含量随贮藏时间的变化如图3c 所示。鲐鱼原料中的组胺含量很低,仅有6.52 mg/kg,远低于FDA 规定的50 mg/kg 的限量[36]。随着贮藏时间的延长,各组中组胺的含量均呈增加趋势。比较第1,2,3 组,第3 组中组胺的增量最多,贮藏结束时含量高达100.33 mg/kg,第1 组和第2 组中组胺含量增加缓慢,贮藏结束时,仍低于50 mg/kg。比较第4,5,6,7 组,第4 组中组胺含量增加最少,贮藏结束时含量仅为32.57 mg/kg,其它3 组均快速增加,贮藏结束时的含量分别达到79.31,86.12,80.62 mg/kg。对照组中组胺的含量在整个贮藏过程中始终处于较低水平,贮藏结束时的含量仅为15.22 mg/kg。对所得的结果进行分析,发现接种柠檬酸杆菌的样品,其组胺含量要显著高于未接种柠檬酸杆菌的样品,说明柠檬酸杆菌是鲐鱼中典型产组胺菌,与组胺的产生密切相关。
2.4.4 菌落总数 灭菌鱼块接种不同菌株后的菌落总数随时间的变化如图3d 所示。鱼块中菌落总数的初始值为0.7 lg(CFU/g),符合灭菌鱼块的标准[18]。第1 组中菌落总数增加迅速,并在第6 天超过了6 lg(CFU/g),贮藏结束时达到8.8 lg(CFU/g),对照组的菌落总数始终维持在较低水平,且变化不大。其它几组菌落总数介于第1 组和对照组之间。致腐能力试验结果表明,希瓦氏菌在鲐鱼腐败过程中挥发性盐基氮和三甲胺的积累方面起到了主要的作用,而柠檬酸杆菌则与组胺的产量具有更加紧密的关联,两者都有可能是鲐鱼冷海水贮藏的特定腐败菌。然而,根据课题组先前研究,发现鲐鱼冷海水贮藏过程中的组胺含量始终在较低水平,因此,产组胺菌对冷海水贮藏下鲐鱼的品质影响较小,此外,接种过柠檬酸杆菌的鱼块中的菌落总数远低于特定腐败菌的标准,这些结果都表明将柠檬酸杆菌作为鲐鱼冷海水贮藏的特定腐败菌是不合适的。因此,研究认定希瓦氏菌为鲐鱼冷海水贮藏的特定腐败菌。

图3 优势腐败菌的致腐能力测定Fig.3 Spoilage potential test of the spoilage bacteria
3 结论
本文通过选择性培养基和16S rDNA 序列分析法,对鲐鱼冷海水贮藏期间的菌相组成和变化进行研究,并通过对优势腐败菌致腐能力的测定,确定了鲐鱼冷海水贮藏的特定腐败菌。结果表明,鲐鱼冷海水贮藏期间的主要菌相组成由嗜冷杆菌变为假单胞菌再到希瓦氏菌。通过对腐败样品中菌相组成的分析,得出鲐鱼冷海水保鲜下的优势腐败菌为希瓦氏菌、肠杆菌和柠檬酸杆菌。通过致腐能力分析表明,希瓦氏菌具有较强的致腐能力,被认定为鲐鱼冷海水贮藏下的特定腐败菌。
