增温改变高寒草甸植物群落异速生长关系
2021-05-12徐满厚杨晓辉秦瑞敏
徐满厚,杨晓辉,杜 荣,秦瑞敏,温 静
(太原师范学院地理科学学院,山西 晋中 030619)
工业革命后,人类生产活动日益频繁,导致大气中CO2、CH4和N2O 等温室气体不断增加,使全球平均气温和海洋温度逐渐升高,在全球尺度上对生态系统产生了明显影响[1-2]。目前全球平均气温已经比工业革命前上升1 ℃左右,如果按现有速度发展,今后每10 年全球平均气温将上升0.2 ℃,到2052 年世界平均气温将比工业革命前高1.5 ℃[3]。全球气候变化导致的气温升高,改变了各地的气候资源,对植物群落产生了直接影响[4-6]。
全球气候变暖对植被的影响已经成为气象学、生态学等各个学科的研究热点。温度是植物生长发育最重要的外界环境因子之一,在植物生长发育全过程中起到重要调节作用,温度升高会对植物的生长产生不可逆的影响[7]。植物的生长主要包含两种关系:同速生长和异速生长,其中同速生长表达为直线函数,异速生长表达为曲线函数[8]。异速生长是生物体两个性状间不成比例生长的现象,它能够揭示植物生长的内在规律,但不同物种常因遗传特性的差异会有不同的生长特点;而异速生长规律主要是描述个体大小与生物体相关属性之间非线性的数量关系,通常用幂函数表达[9]。关于异速生长理论的研究,前人做了大量积累,研究对象多集中于木本植物(乔木、灌木)[10-18],研究指标涉及茎(枝长[10-11]、基径[10]、枝茎[12]、胸径[13])、叶(叶片形态与结构[13])、生物量(茎生物量[14]、叶生物量[12,15]、地上生物量[16-17]、地下生物量[16-17])、物质能量代谢[18](光合速率、呼吸速率)以及养分含量(氮、磷)[18]等,通过对这些生长指标之间的关系进行深入研究,可以更清晰地揭示出木本植物结构、功能及生理属性间的异速关系[18]。已有对草本植物异速生长关系的研究,主要是对草本植物的生物量[8,19-21]、物质能量代谢[20]、养分含量[22-23]等指标间的关系进行分析。虽然从木本植物到草本植物各生长指标间都有异速关系的体现,但是当分布区域[16]、树种[16,24]、密度[20,23]、径级[25]、立地环境[10,17]、竞争强度[20]等条件不同时,植物的生长关系会发生改变,导致植物的生长关系在同速生长和异速生长之间发生转化。如在不同沙地的研究表明,黄柳(Salix gordejevii)丛生枝枝长与基径在流动沙地表现为异速生长关系,在半固定沙地和固定沙地则调整为等速生长,于丘间低地再次转变为异速生长[10];在不同海拔的研究也表明,弓杠岭高海拔区陇蜀杜鹃(Rhododendron przewalskii)小枝茎长与茎生物量之间的关系表现为异速生长,而低海拔区则表现为等速生长关系[14]。
青藏高原海拔较高、气候类型独特,且对气候变暖具有特殊的敏感性,所以被称为“世界第三极”[26-27]。高寒草甸作为青藏高原隆起和长期低温产生的特殊产物,对全球变暖也异常敏感[28-29]。气候变暖是导致青藏高原高寒草甸植被个体水平(物候与生长、光合与呼吸)、群落水平(生物量生产与分配、群落结构、物种多样性)改变的重要因素,其对高寒草甸的影响有利有弊,应采取积极有效的措施来缓解环境因子变化的负面效应[7]。徐满厚和薛娴[28]研究认为,高寒草甸植被在短期(1 年) 增温下,其地上部分近似等速生长,地下部分为异速生长,整体表现为异速生长。然而,高寒植被生长对温度升高的响应不仅取决于植物种类和水热环境,还与增温的持续时间有关。长期增温不仅影响植物的生长发育,还有可能改变由短期增温引起的生长模式,因此需要注意增温试验的时间效应[28]。本研究利用为期8 年的增温试验对高寒草甸植被的生长关系进行深入研究,可以进一步揭示高寒草甸植被生长关系在长期增温下的变化规律。针对异速生长理论的研究很多,但研究高寒草甸植被的不多,且关于模拟增温对异速关系的影响更是少见。因此,本研究以青藏高原高寒草甸植被为研究对象,基于长达8 年(2011-2018 年) 的野外模拟增温试验,分析对照和增温处理下高寒草甸植物群落的生长关系,进而探讨模拟增温对青藏高原高寒草甸植物群落生长关系的影响,以期为高海拔地区高寒植被的有效保护与合理利用提供数据支持和理论指导。
1 材料与方法
1.1 研究区概况
本研究以青藏高原高寒草甸为研究对象,研究区位于青藏高原北麓河试验站(中国科学院冻土工程国家重点实验室青藏高原研究基地,图1a),地处五道梁与风火山之间,34°49′34″ - 34°49′37″ N,92°55′57″ - 92°56′06″ E[28,30]。试验站海拔在4 620~4 640 m,该地区属于青藏高原干寒气候,以干旱、寒冷为主要气候特征,气温随海拔升高而降低,气温日较差大,干湿分明,年均气温-5.9 ℃,年均降水量267.6 mm,年均相对湿度57%,降水主要集中在6 月 -8 月,风向主要以西北风为主。该研究区的植被为高寒草甸类型,以莎草科和菊科为主,生长季植被盖度高达83%;土壤为高山草甸土,以沙粒居多,剖面发育具有薄层性和粗骨性特点[28]。
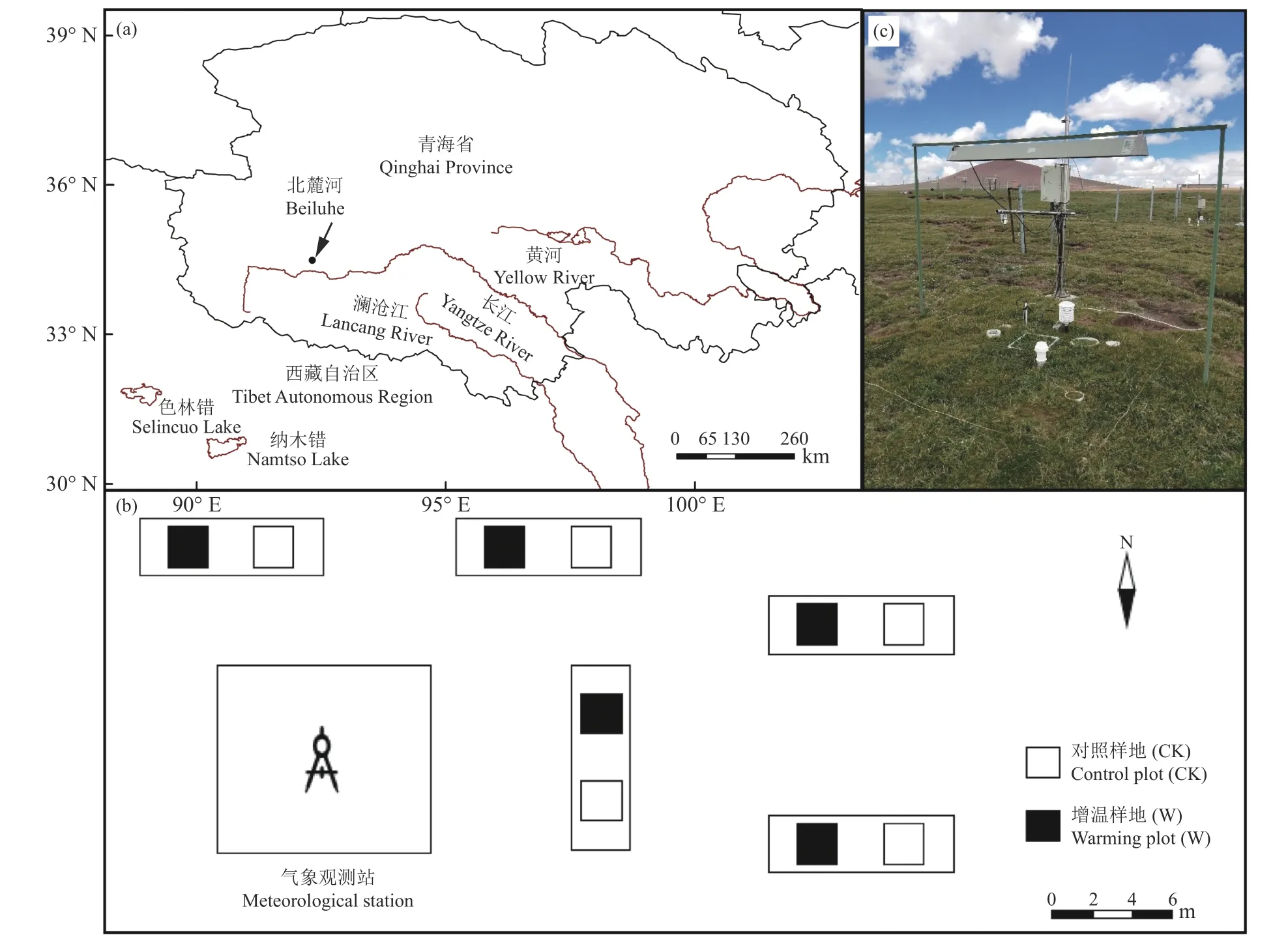
图 1 研究区地理位置及样地设计示意图Figure 1 Geographical position and design sketch of sample site
1.2 样地设置
试验样地位于距北麓河试验站约300 m 的典型高寒草甸地段,该地段地势较为平坦,植被分布均匀,未受放牧、鼠兔啃食等干扰。试验设置不增温(对照)和增温两个处理,每个处理设置5 个样方(5 个重复),共10 个样方(图1b)。于2010 年开始采用红外线辐射器进行模拟增温,8 年内可使样地空气温度平均增加0.19 ℃、土壤温度平均增加1.23 ℃[28,31-33]。该增温仪器由美国Kalgolo Electronics 公司生产,灯体为三棱形,灯管为圆柱体,架设在样地正上方距离地面1.5 m 处(图1c);对照样地只架设相同的支架,以消除支架遮阴对植被造成的影响;所有样地外围均用铁丝网进行防护。试验中所用到的数据采集器均由美国Campbell Scientific 公司生产,样地及仪器的具体描述详见参考文献[28, 31-33]。
1.3 植被特征调查
本研究从2010 年7 月到2018 年10 月进行长达8 年不间断的模拟增温,分别在2011、2012、2013、2016、2017、2018 年植物生长季末期(8 月底至9 月初)[28,31-33]进行群落生长特征调查,使用27 cm ×27 cm 的样方框(边宽2 cm,内部面积为25 cm ×25 cm)[28,31-33]测定,样方框用硬铁丝网分割成2.5 cm ×2.5 cm 的网格100 个,涉及到的指标为密度、高度、盖度、频度、地上生物量、地下生物量、总生物量和根冠比。密度指单位面积或单位空间内该物种的数目,高度为该物种自然状态下垂直于地面的高度,盖度为样方框中植被覆盖的格子数,频度为该物种出现在每一小格中的次数。地上生物量指植被在某一特定时刻单位空间的重量,在试验样地(对照、增温)周围选取高度和盖度与之较为一致的植被作为临时性样方,采用刈割法齐地面收割地上生物量,建立回归方程推导试验样地的地上生物量;地下生物量使用土钻法获取,以10 cm 为一层,采集0 - 10 cm、10 - 20 cm、20 - 30 cm、30 - 40 cm、40 - 50 cm 共5 层土壤的地下生物量,为区分根系中的活根与死根,将根斜切之后根据断面的颜色、根的柔韧性及是否附着毛根进行判断,即断面颜色灰白、柔韧性强、附着毛根的根定为活根[28,33]。把刈割下的地上部分和筛分出的活根放进80 ℃烘箱中烘干至恒重,称重后分别获得地上生物量和地下生物量;总生物量为地上生物量与地下生物量之和;根冠比为地下生物量与地上生物量之比。
1.4 数据分析
数据采用Excel 2020 整理与统计,使用的指标(密度、高度、盖度、频度、地上生物量、地下生物量、总生物量、根冠比) 共8 个,时间尺度(2011、2012、2013、2016、2017、2018 年)共6 年,在两个不同处理(对照、增温)下有10 个样方。使用SPSS 23 软件进行如下分析:1)通过描述性统计分析各植被生长指标在对照和增温两个处理下的基本数量特征;2)采用相关分析法进一步探究各生长指标之间的相互关系;3) 对生物量指标(地上生物量、地下生物量、总生物量) 与其他生长指标(密度、高度、频度、盖度)进行曲线拟合,在指数函数、线性函数、对数函数、幂函数4 个常用基本函数中选择最优方程(选择标准:决定系数R2值最大,显著性水平P 值最小且小于0.05),研究对照和增温处理下植物群落的生长关系,进而探讨模拟增温对青藏高原高寒草甸植物群落异速生长关系的影响。
2 结果与分析
2.1 不同处理下植被生长指标的基本数量特征
增温对植物生长产生一定影响,但各生长指标对增温的响应不一(表1)。其中,密度、地下生物量、总生物量、根冠比的数据范围呈缩小趋势,变异系数均小于对照,数据更加集中;高度、频度、盖度的数据范围略有扩大,变异系数均高于对照,数据更加离散;地上生物量的数据范围呈扩大趋势,但其变异系数略有减小,数据趋于集中。在增温处理下,地上生物量、地下生物量、总生物量的均值均大于对照,且地上生物量的变异系数为0.38,低于地下生物量(0.79)及根冠比(0.80),这说明增温促进了植被生物量积累,且对生物量在地上、地下部分的分配产生了影响。另外,对各植被指标在对照和增温处理间的差异进行单因素方差分析,发现只有植被高度达到显著水平(P = 0.030),说明增温能够显著增加植物高度。
2.2 不同处理下植被生长指标间的相互作用
相关分析(表2)表明,在对照处理下,地上生物量与密度、高度、盖度均呈显著或极显著正相关关系,三者的相关系数分别为0.380 (P < 0.05)、0.647(P < 0.01)、0.798 (P < 0.01),说明地上生物量随着密度、高度、盖度的增加而显著增加,并受盖度影响最大;地下生物量、总生物量和根冠比与密度、高度、频度、盖度均呈负相关关系,但相关性未达到显著水平。

表 1 不增温对照与增温处理下的植被生长指标Table 1 Basic quantitative characteristics of vegetation growth indexes under control and warming treatments
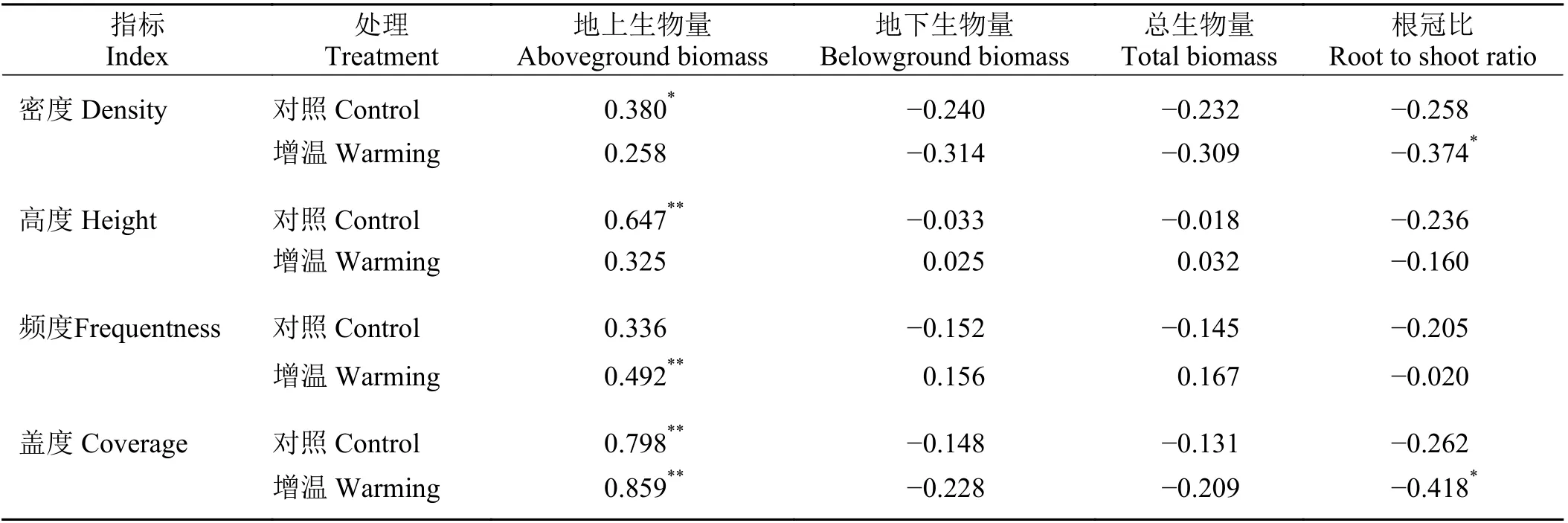
表 2 不增温对照与增温处理下植被生长指标间的相关系数Table 2 Correlation coefficient of vegetation growth indexes under control and warming treatments
在增温处理下,地上生物量与频度和盖度均呈极显著正相关关系(P < 0.01),并受盖度影响大;根冠比与密度和盖度均呈显著负相关关系(P < 0.05),且受盖度影响大,说明随着密度、盖度的增加,生物量更多的分配到地上部分;地下生物量、总生物量与密度、高度、频度、盖度均呈负相关关系,但相关性未达到显著水平。
因此,随温度升高,盖度对地上生物量的影响增强(P < 0.01),频度对地上生物量的影响由不显著(P > 0.05)变为极显著正相关(P < 0.01),密度、高度对地上生物量的影响由显著正相关变为不显著;地下生物量、总生物量与密度、盖度的相关性增强,但仍未达到显著水平;地下生物量、总生物量与高度、频度的相关性均由负相关变为正相关,同样未达到显著水平;密度和盖度对根冠比的影响也由不显著(P > 0.05)变为显著负相关(P < 0.05),高度、频度与根冠比的相关系数变小且不显著。
2.3 不同处理下植被生长关系的类型及其变化
异速生长规律通常用幂函数表达,故选取4 个常用基本函数(指数函数、线性函数、对数函数、幂函数)对各植被生长指标进行最优方程拟合(表3),结果发现,幂函数更加符合青藏高原高寒草甸植被的生长关系,且幂指数的变化范围在-1.097~0.570,说明高寒草甸植被符合异速生长理论。

表 3 不增温对照与增温处理下植被生长指标间的最优函数方程Table 3 Optimal functional equations of vegetation growth indexes under control and warming treatments
对照处理下,地上生物量与频度呈幂函数关系,高寒草甸表现为异速生长;地上生物量与密度、高度、盖度均呈线性函数,植被表现为等速生长模式;地上生物量与盖度的决定系数(0.635 9)大于高度(0.405 4)和密度(0.144 7),地上生物量与密度、高度、盖度呈正相关关系,并受盖度影响最大;总体而言地上部分表现为等速生长。就地下生物量而言,与盖度呈指数函数,与频度则呈线性函数关系;密度、高度与地下生物量均呈幂函数;除去频度外,其余3 个变量与地下生物量均符合异速生长规律;因此,高寒草甸地下部分表现为异速生长。总生物量与密度、高度、频度、盖度均呈负相关关系;由于总生物量与高度符合幂函数关系,因而高寒草甸植被在对照处理下整体符合异速生长。综上,在对照处理下,高寒草甸植被地上部分表现为等速生长,地下部分为异速生长,整体表现为异速生长关系。
在增温处理下,地上生物量与密度、高度、频度的幂指数均为正数,说明地上生物量会随着密度、高度、频度的增加而增加,但3 个函数的幂指数均较小,表明高寒草甸植被地上部分符合异速生长关系。高度、盖度与地下生物量均表现为幂函数,因此植被表现为异速生长关系,由于幂指数为负,所以地下生物量随着高度与盖度的增加呈递减趋势;而地下生物量与频度则表现为线性模型,符合等速生长规律。总生物量与高度、盖度均呈幂函数,幂指数分别为-0.335 和-0.599,说明高寒草甸植被表现为异速生长;总生物量与密度呈指数函数关系,虽然底数大于1,但是指数为负,说明总生物量随着密度的增加而减少;总生物量与频度同样呈指数函数关系,但指数为正,因此总生物量随着频度的增加而增加。总体而言,在模拟增温下,高寒草甸的地上部分与地下部分均符合异速生长理论,均表现为异速生长关系。
因此,随温度升高,高寒草甸植被地上部分由等速生长关系变为异速生长关系,地下部分的异速生长关系进一步增强,即增温促进了青藏高原高寒草甸植物群落的异速生长关系。
3 讨论
3.1 模拟增温对植被生物量的影响
气候变暖是造成植物生物量生产、分配改变的重要因素[7,34]。本研究发现,增温导致植物群落特征变化出现差异,其中高度、地上生物量、地下生物量、总生物量呈增加趋势,高度显著增加,但密度、频度、盖度呈减小趋势;同时增温促使地上生物量、地下生物量、总生物量、根冠比、密度的数据更加集中,而高度、频度、盖度的数据更加离散。
关于增温对植被生物量的影响,不同学者得到的结论差异较大。如有研究发现,增温导致植被的盖度、高度均有不同程度增加[35],且随着增温幅度越大,盖度和高度的增幅也越大[36],但干珠扎布[1]认为高增温下,植被盖度降低;有研究发现,增温促进植被生物量(地上生物量、地下生物量、总生物量)的增加[36-37],但也有研究发现,增温造成叶片生物量和根生物量的减少[38];还有研究认为,增温造成植被根冠比的显著降低[37-38]。此外,Yang 等[39]却认为,气候条件的改变不会造成青藏高原高寒草地地上与地下部分的分配关系的变化。
一般认为,由于受水分和植被类型不同的影响,增温可导致高寒草地生物量变化存在很大的区域性差异[28,33]。在全球变暖背景下,温度升高已是必然,同时造成了降水格局的改变[40]。降水是干旱、半干旱地区植被生长的制约因子[41-42],随着降水量的增加,草本植物地上生物量[40,43-45]、地下生物量[43,46]、总生物量[47]均呈线性增加趋势,且群落生物量与浅层土壤含水量显著正相关[48]。当降水增加30%时,地上生物量达到最大值[47],但也有学者得出地上生物量在降水量增加60% 时最大[49]。分析认为这可能与植被类型差异有关,水分添加对禾本科植物生物量的影响最明显,而对杂类草植物生物量的影响不明显[50]。
3.2 模拟增温对植被异速生长关系的影响
本研究发现,幂函数更加符合青藏高原高寒草甸植被的生长关系,说明青藏高原高寒草甸植被符合异速生长理论;在不增温对照下,高寒草甸植被地上、地下部分表现出不同的生长模式,地上为等速生长,地下部分为异速生长,整体表现为异速生长关系;在模拟增温后,地上部分和地下部分均表现为异速生长关系,总体也体现为异速生长,说明在增温处理下高寒草甸植被地上部分由等速生长关系变为异速生长关系,地下部分的异速生长关系进一步增强。
对不同功能型植被的研究发现,天然常绿阔叶林[16]、杉木人工林[16]、天然次生混交林[11]、檵木(Loropetalum chinense)[25]、 新 疆 绢 蒿(Seriphidium kaschgaricum)[51]等植物的地上和地下生物量分配均符合等速生长规律,而对节刺(Sageretia pycnophylla)[51]、骆驼刺(Alhagi sparsifolia)[51]等灌木植物则表现为异速生长,这表明植物生长关系受植被功能型影响较大,不同物种的生长关系也不一样。对同一物种的不同个体来说(如檵木),随着个体数量增大,各器官间的生长关系由等速生长变为异速生长,生物量也更多分配于地上部分[25]。因此,同一物种不同器官间的生长关系随着个体密度的变化而发生转变。
另外,植物生长关系的转化与生物量的分配也有关系,而生物量的生产及分配受降水、温度影响较大,因此在某种程度上植物生长关系的转化受限于气候变化。研究表明,降水减少可造成生物量向地下分配的比例增加[52];随着温度升高,高寒草甸群落的高度增加[53]、盖度降低[54],生物量更多分配于地上部分[55],而高度增温则会造成高寒草甸地上生物量减少[54]。徐满厚和薛娴[28]研究表明,高寒草甸植被地上部分近似等速生长、地下部分为异速生长、总体而言表现为异速生长,与本研究在对照处理下的结果相近。此外,刘冠志[10]在分析黄柳(Salix gordejevii)的异速生长关系时,发现水分梯度的增加会改变枝长与基径的生长关系,即从流动沙地的异速生长,调整为半固定沙地和固定沙地的等速生长,于丘间低地再次调整为异速生长,说明水分因子也会造成植被生长关系的变化。
通过以上分析可知,植被各器官间的生长关系大多符合异速生长理论,但由于所选取的植被功能型、物种组成、个体密度不同以及所处的气候条件有差异,导致植被生长关系在等速生长和异速生长之间出现转化。因此,在本研究中,高寒草甸地上部分在增温处理下由等速生长变为异速生长,很可能是由增温引起的密度、频度、盖度等指标发生改变而导致的,且根冠比与密度、盖度均呈显著负相关关系,这说明随着密度、盖度增加,高寒草甸生物量更多分配到地上部分。
4 结论
青藏高原高寒草甸地上、地下部分的环境因子差异较大,且物种具有独特的生物学特征,所以高寒草甸地上与地下生物量对增温响应不同,增温使植被的地上生物量增加,但造成地下生物量与总生物量的减少。青藏高原高寒草甸植被符合异速生长理论,但地上与地下的分配模式不同。在不增温对照下,地上部分表现为等速生长,地下部分与整体均表现为异速生长关系;增温处理下,地上部分、地下部分及总体均表现为异速生长关系,说明增温会造成植被异速生长关系的变化,具体表现为增温使植被地上部分由等速生长关系变为异速生长关系,地下部分的异速生长关系进一步增强。
本研究采用了为期8 年的增温试验数据,探究了对照、增温两个处理下高寒植被的生长关系变化。今后应对时间尺度重新划分,更加具体地分析短期增温、长期增温下植被生长关系的变化,且增加一些气候因子的数据,对造成植被生长关系变化的原因进一步阐释。本研究结果对正确理解气候变化如何影响高寒草甸植物生长与群落结构具有科学理论意义,同时对指导高海拔地区高寒植被的有效保护与合理利用具有实践应用价值。
