鱼类致病嗜水气单胞菌外膜蛋白P5的原核表达、免疫保护及抗血浆杀菌作用研究
2021-04-26简思杰伍娜娜
荣 娜,简思杰,孙 薇,康 超,伍娜娜,刘 祥
(陕西理工大学 生物科学与工程学院,中德天然产物研究所,陕西汉中 723001)
嗜水气单胞菌 (Aeromonashydrophila)属于弧菌科气单胞菌属,是一种在水生环境中广泛存在的条件致病菌,可诱发水生动物局部出血和败血症,给水产养殖业带来巨大的收入损失[1-2]。对于该菌的防治,传统上主要采用抗生素药物,但长期使用会导致菌株抗药、机体抗生素残留和水体污染等问题[3]。疫苗免疫安全性高、无毒害,但目前普遍应用于水产养殖的灭活疫苗存在免疫效果欠佳和二次感染的风险[4-5],因此有必要研制有效防治嗜水气单胞菌的新型疫苗。亚单位疫苗是一种以纯化的蛋白、核酸或多糖等免疫活性片段作为组分的新型疫苗,具有安全稳定且免疫持久等优点[6-7],受到广泛关注。
外膜蛋白(Outer membrane protein,OMP)是细菌外膜的主要构成部分,在维持细菌结构完整性和物质转运等方面有着重要作用[8-9]。此外,外膜蛋白具有高度的免疫原性,是一种潜在的保护性抗原[10]。P5蛋白是革兰氏阴性细菌的一个主要外膜蛋白,也是OmpA蛋白的超家族成员,参与了细菌在体内的定植[11]。有研究表明,P5蛋白免疫原性较强,可激发机体体液免疫产生保护性抗体,是参与机体免疫应答的早期抗原[12]。谢乐新等[13]将副猪嗜血杆菌外膜蛋白P5分段表达,发现表达的3个蛋白均表现出良好的免疫原性和抗原性。王海峰等[14]研究显示,P5免疫动物后能诱导产生高水平特异性抗体,并可显著保护豚鼠抵抗副猪嗜血杆菌感染。P5也是嗜水气单胞菌的主要外膜蛋白,然而关于该蛋白的免疫功能研究尚未见报道。
基于以上现状,本研究选取嗜水气单胞菌主要外膜蛋白P5为研究对象,原核表达纯化P5蛋白并确定其最佳诱导表达条件;将纯化的P5蛋白免疫小鼠制备P5多克隆抗血清,探究P5抗血清对红鲫鱼的免疫保护效果;体外评价P5蛋白抵抗鱼血浆杀菌的作用,结合P5蛋白的生物信息学分析,以期为嗜水气单胞菌蛋白亚单位疫苗的研究奠定理论基础。
1 材料与方法
1.1 材 料
1.1.1 菌株及供试动物 嗜水气单胞菌ATCC 7966、Escherichiacoli(E.coli) DH5α、BL21菌株及pET-32a质粒由陕西理工大学中德天然产物研究所保存;4~5周龄昆明鼠购自西安交通大学医学部,试验前饲养3 d,每日定量喂食供水;红鲫鱼购自汉中水产市场,外表健康无病,平均体质量约30 g,身长15 cm左右,试验前于养殖水箱饲养1周,每3 d换水1次,每次换掉1/3,养鱼用水经除氯气处理,水温控制为(27±1) ℃,每天早晚按照体质量1%投喂鲫鱼专用鱼粮(厦门华普水产有限公司),24 h供氧。1周后红鲫鱼呈健康状态,未出现任何感染症状。
1.1.2 主要试剂 rTaq聚合酶、T4-DNA连接酶、内切酶EcoR Ⅰ 和HindⅢ购自TaKaRa公司;细菌基因组提取、质粒提取试剂盒购自上海生工公司;IPTG、TMB显色液为西安赫特生物科技有限公司产品;HRP标记山羊抗小鼠IgG购于Sigma公司;引物合成、基因测序由天润奥科生物公司提供。
1.2 方 法
1.2.1 生物信息学分析 从NCBI数据库获取不同血清型嗜水气单胞菌和不同种类气单胞菌的P5蛋白序列:嗜水气单胞菌ATCC 7966(GenBank:ABK37458.1)、GYK1(ABK37458.1)和ML09-119(AGM45792.1),达氏气单胞菌(登录号:WP_026142030.1),兽生气单胞菌(WP_043554548.1),鲑鱼气单胞菌(WP_099994858.1),豚鼠气单胞菌(WP_109111770.1),沙氏气单胞菌(WP_131731585.1)。通过GeneDoc软件绘制多重序列比对图,利用MEGA软件构建系统发育树;采用ProParam数据库预测P5的氨基酸数、分子质量大小、等电点、稳定指数等参数,并从ProScale analysis数据库中获取亲水性图谱;采用TMHMM 2.0和Signal P 3.0软件预测P5的跨膜区域和信号肽位置,并利用SOPMA和SWISS-MODEL软件分别进行二级结构和三维结构预测。最后使用STRING软件在线构建P5与其他蛋白的相互作用关系网络。
1.2.2 重组菌株的构建 根据NCBI数据库中嗜水气单胞菌ATCC 7966全基因组序列(Ge-nBank:CP000462.1),设计引物扩增P5基因。F-P5:CAAGAATTCATGAATAAAACACTG ATT;R-P5:CATAAGCTTTCACTGCTGA-ACTTCCGA(划线部分分别为EcoR Ⅰ和HindⅢ的酶切位点)。以全基因组为模板,利用rTaq聚合酶实现P5基因扩增,反应体系为50 μL,退火温度设为55 ℃。扩增产物经核酸电泳鉴定并回收,EcoR Ⅰ和HindⅢ同时双酶切PCR产物与载体pET-32a,在T4-DNA连接酶作用下将P5基因导入质粒pET-32a,转化E.coliDH5a中,获得重组质粒。双酶切和基因测序进一步验证重组质粒,将构建成功的重组质粒转化至E.coliBL21中,构建P5原核表达菌株。
1.2.3 重组蛋白的诱导表达及纯化 挑取P5表达菌的单菌落过夜培养,以1∶100转接培养至OD600为0.6,加入0.1 mol/L的IPTG,摇床诱导6 h,取1 mL菌液收集菌体,加入300 μL 2×SDS吹打混匀,高温变性蛋白5 min,离心取上清10 μL进行SDS-PAGE电泳,检测P5的表达情况,结合包涵体洗涤和SDS-PAGE蛋白电泳,切胶法纯化P5蛋白[15]。
1.2.4 重组蛋白表达条件的优化 以L9(34)模型(表1)进行P5的最佳诱导表达条件研究。简要过程为:以1∶100将过夜培养的P5饱和菌液转接至600 mL培养液中,37 ℃、200 r/min培养至设定的不同OD600值,按照不同试验组合要求加入不同终浓度的IPTG,并在规定条件下诱导,每个组合进行3组重复。收取1 mL诱导菌液,沉淀加入300 μL 2×SDS,高温变性蛋白5 min,样品通过SDS-PAGE电泳获得不同诱导条件下P5蛋白的表达图谱。使用软件Phoretix 1D和SPSS 13.0分别进行表达图谱光密度和不同因子的显著性分析。

表1 P5表达条件试验的因子与水平Table 1 Factors and levels of P5 expression condition test
1.2.5 蛋白小鼠抗血清的制备及特异性检测 昆明鼠试验组和对照组各15只,试验组每只取纯化的P5蛋白100 μg混合100 μL弗氏完全佐剂,腹腔注射免疫小鼠。14 d后,等量蛋白混合100 μL弗氏不完全佐剂,二次免疫小鼠。7 d后,同二免,进行第三次免疫,对照组免疫PBS。1周后眼球取血,静置,待血清自然析出后于4 ℃、3 000 r/min离心10 min,收集抗血清于-80 ℃保存,备用。Western blotting验证P5抗血清特异性,步骤简述如下:SDS-PAGE电泳嗜水气单胞菌全蛋白,恒压80 V转NC膜1 h,NC膜充分封闭 2 h,然后与不同稀释倍数(1∶400,1∶800,1∶1 600,1∶3 200)的P5抗血清于37 ℃轻摇孵育40 min,对照为阴性抗血清。用TNT缓冲液冲洗3次,再与1∶3 000倍稀释的二抗于37 ℃轻摇孵育1 h,洗涤后通过DAB显色法确定P5抗血清的特异性。
1.2.6 体外模拟蛋白抗血清与嗜水气单胞菌的相互作用 ELISA[16]检测蛋白抗血清与嗜水气单胞菌的相互作用:培养嗜水气单胞菌至OD600约1.0,菌体用φ=1%的甲醛生理盐水80 ℃灭活,生理盐水调整菌液OD600=0.2,分装为1 mL/管(菌量108CFU),小管菌分别与100 μL不同稀释倍数的蛋白抗血清(1∶400,1∶800,1∶1 600,1∶3 200,1∶6 400)孵育90 min,对照为阴性血清,再与200 μL 1∶3 000倍稀释的二抗孵育1 h,洗涤菌体并混合20 μL PBS至酶标板中,加入200 μL TMB显色液,避光显色10 min,加入终止液2 mol/L H2SO450 μL,450 nm读数。
1.2.7 抗血清的被动免疫保护试验 红鲫鱼试验组和对照组各15尾,试验前已饲养1周,确定未感染嗜水气单胞菌。取嗜水气单胞菌甘油菌接种至600 mL的LB培养基中,28 ℃培养至OD600为1.0,菌体经生理盐水洗涤并调整OD600至1.0(菌浓度109CFU/mL)。依据抗血清效价,取P5抗血清腹腔注射15尾红鲫鱼,每尾60 μL(对照注射生理盐水),2 h后,以嗜水气单胞菌对红鲫鱼的半数致死量(4×109CFU)感染试验鱼,观察并记录试验鱼的病变及死亡情况,当死亡数稳定时,统计试验组与对照组受试鱼的死亡率,并计算免疫保护率(RPS)。RPS=1-(试验组死亡率/对照组死亡率)×100%[15]。
1.2.8 蛋白抵抗鱼血浆的杀菌作用 28 ℃过夜培养嗜水气单胞菌,以1∶100转接至100 mL新的培养液中,培养至OD600为0.5,菌体用φ= 0.85%的生理盐水洗涤并调整OD600为1.0,收取7管1 mL菌液离心获取菌体,小管菌分别与600 μL不同稀释倍数的鱼血浆(未稀释血浆,1∶2,1∶4,1∶8,1∶16,1∶32)于28 ℃、200 r/min孵育5 h,对照为生理盐水。孵育后菌体用生理盐水洗涤3次,SDS-PAGE分离全蛋白并转NC膜,NC膜充分封闭后依次与一抗、二抗孵育,通过DAB显色液进行显色。对比特异性条带的亮度,分析P5的差异表达以评价其抵抗鱼血浆的杀菌作用。
2 结果与分析
2.1 生物信息学分析
2.1.1 P5同源性与系统发育分析 通过GeneDoc软件绘制不同菌株P5蛋白氨基酸序列比对图(图1),结果显示,不同血清型的嗜水气单胞菌以及气单胞菌属间的P5蛋白同源性均较好。利用MEGA软件构建系统发育树(图2),发现3种不同血清型嗜水气单胞菌的亲缘关系最为紧密,且不同气单胞菌间亲缘关系也较近。因此推测,P5蛋白免疫动物产生的抗体可能为不同种类的气单胞菌提供交叉免疫保护作用。
2.1.2 P5理化性质、结构与蛋白互作预测 ProtParam数据库分析显示:P5含有351个氨基酸,分子质量为36 830.20 u,等电点为4.65,半衰期大于10 h,不稳定指数为25.68,归类为稳定蛋白;亲水性图谱分析得出P5亲水性平均值为-0.178,属于亲水性蛋白(图3-A);跨膜结构预测图谱(图3-B)显示,全序列中预测到的跨膜区域可信度较低,该蛋白属于外膜蛋白;信号肽预测结果(图3-C)显示,P5的N端1~21位氨基酸组成了信号肽序列,21~22位氨基酸之间存在切割位点,可信度达0.967 7;SOPMA软件的二级结构预测显示,α-螺旋占全序列24.1%,延伸链占 23.6%,无规则卷曲占52.3%(图3-D);SWISS-MODEL软件的预测结果显示,P5的三维结构为桶状(图3-E);通过STRING数据库构建蛋白互作网络,发现有10种蛋白可与P5相互作用,其中蛋白rpsL和rplD与P5作用关系最为紧密(图3-F)。
2.2 P5重组质粒的构建及蛋白表达纯化
利用引物F-P5、R-P5扩增P5基因,电泳显示在约1 000 bp处存在单一条带(图4-A),大小与理论值1 059 bp相符。重组质粒pET-32a-P5经EcoRⅠ和HindⅢ双酶切,得到一条约1 000 bp的条带,大小符合预期(图4-B)。目的基因经测序比对,显示与 NCBI 数据库所公布的P5基因序列一致,表明成功构建重组质粒。重组质粒转化至E.coliBL21获得P5表达菌,经IPTG诱导后,电泳显示目标蛋白成功表达,分子质量大小约59 ku;利用包涵体洗涤和SDS-PAGE电泳切胶法纯化获得P5蛋白(图4-C)。
2.3 P5蛋白表达条件的优化
诱导菌经SDS-PAGE电泳获得不同诱导条件下P5的表达图谱(图5),图谱光密度值如表2所示,所得数据极差和方差分析如表3和表4所示。从表3可得出,在诱导P5菌株表达时,最佳条件组合为A3B3C2D2,即在菌液OD600为1.0时加入终浓度为0.5 mmol/L的IPTG,32 ℃诱导8 h。表4分析结果显示,加诱导剂时菌液OD600值达到显著性,由此可得诱导时菌液浓度对于P5蛋白的高效表达尤为重要。
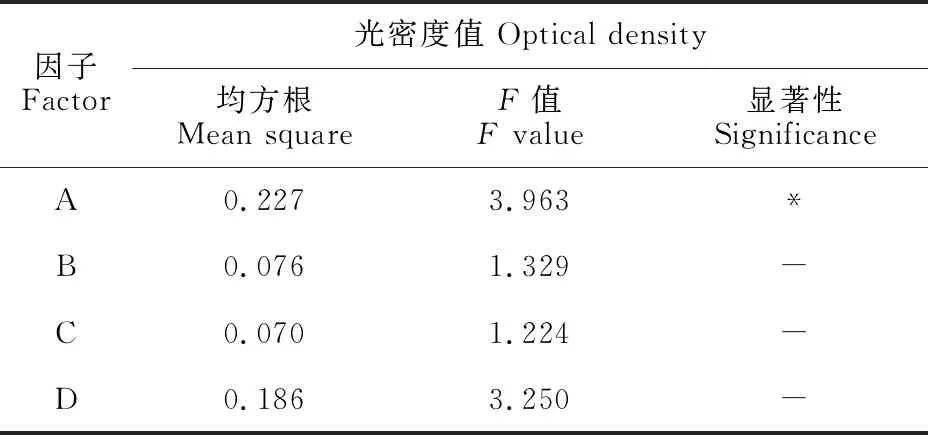
表4 P5表达图谱光密度数值的方差分析Table 4 Variance analysis of optical density of P5 expression map

表2 P5蛋白表达图谱光密度分析Table 2 Opticaldensity analysis of P5 expression map

表3 P5表达图谱光密度数值的极差分析Table 3 Range analysis of optical density of P5 expression map
2.4 P5蛋白抗血清的特异性与效价检测
Western blotting结果显示P5抗血清与P5蛋白特异性结合,呈现出明显的单一条带,阴性对照无条带,表明P5具有良好的免疫原性,可刺激机体产生特异性抗血清,效价达到1∶1 600 (图6)。
2.5 P5蛋白抗血清与嗜水气单胞菌的相互作用
ELISA体外模拟P5抗血清与嗜水气单胞菌的相互作用,结果显示,随着抗血清稀释倍数的增大,P5抗血清与嗜水气单胞菌的结合能力逐渐减弱,当抗体滴度达1∶3 200时,仍可检测到两者的相互作用(图7)。可见,P5抗血清与嗜水气单胞菌在体外存在相互作用,表明P5蛋白具有良好的抗原性。
2.6 P5蛋白抗血清对红鲫鱼的被动免疫评价
P5抗血清被动免疫红鲫鱼,嗜水气单胞菌攻毒后观察试验鱼15 d,发现攻毒后3 d内试验组鱼体出现部分死亡,死亡情况于第4天稳定。对照组48 h内鱼体出现大量死亡,随后死亡率降低,至第5天死亡情况稳定,试验组与对照组试验鱼存活曲线如图8所示。试验鱼死亡症状为:体表有血斑,鱼鳃和各鳍基部充血,腹部突出,肛门红肿。经统计计算,第5天即死亡情况稳定时试验组死亡率为32.3%,对照组73.3%,P5抗血清的免疫保护率为56.0%,SPSS 13.0分析显示与对照组相比差异显著(表5),表明P5蛋白抗血清免疫红鲫鱼对嗜水气单胞菌感染具有一定的免疫保护作用。
2.7 P5蛋白抵抗鱼血浆的杀菌作用
体外模拟P5蛋白抵抗鱼血浆对嗜水气单胞菌的杀菌作用,采用Western blotting检测P5蛋白与鱼血浆作用后的差异表达情况。结果显示,试验组与对照组P5蛋白均检测到特异性条带,且随着血浆浓度的降低,P5蛋白的表达呈上调趋势(图9),推测P5蛋白可能为孔道蛋白,关闭孔道可以有效抵抗血浆杀菌因子进入细胞而杀伤菌体。
3 结论与讨论
嗜水气单胞菌是水产养殖鱼类爆发性传染病的主要病原菌,也是一种人、畜及水生动物共患的条件致病菌。P5蛋白是嗜水气单胞菌的主要外膜蛋白之一,具有较强的免疫原性[17],本研究发现P5蛋白免疫小鼠产生的抗体可增强红鲫鱼对病菌的抵抗力,因此,在疫苗研制方面,P5蛋白具有很好的应用前景。
生物信息学可准确预测蛋白的理化性质、结构与抗原表位,为蛋白功能研究提供参考价值[18]。本研究分析发现,P5为稳定的亲水性蛋白,在气单胞菌属间进化保守,推测P5蛋白在不同气单胞菌间功能相近,所产生的抗体可能为多种气单胞菌提供交叉免疫保护。此外,其二级结构主要以无规则卷曲为主,该结构易扭曲盘旋而暴露在蛋白外层,形成优势细胞抗原表位的可能性较大[19-20]。蛋白表达由于受到原核表达系统的影响,其最佳表达条件往往是唯一的,本研究发酵试验发现,菌液OD600为1.0,IPTG终浓度为0.5 mmol/L,32 ℃诱导8 h,P5获得最大表达量。有研究报道IPTG具有一定毒性,诱导蛋白表达时应添加合适浓度的IPTG,另外,长时间诱导会导致蛋白被细菌蛋白酶降解,不利于蛋白的高效表达[21-22],本研究也发现适度的诱导剂浓度与诱导时间更利于P5蛋白的高效表达。
研究显示,外膜蛋白是嗜水气单胞菌的主要毒力因子之一,能够激发机体产生高水平保护性抗体,且利用原核表达纯化的蛋白免疫动物是制备蛋白多抗的普遍方法[23-24]。毛然然等[25]用纯化的OprM蛋白肌肉注射斑马鱼,在转录水平检测到相关免疫基因表达上调,揭示该蛋白可在斑马鱼体内引起免疫应答。Maiti等[26]利用外膜蛋白OmpW在兔体内制备了抗重组蛋白的多克隆抗体,并证实抗体的特异性反应,表明OmpW可能是一种潜在保护原。Khushiramani等[27]用利用重组外膜蛋白Omp48制备了兔抗免疫血清,且免疫Omp48的鱼体可有效抵抗嗜水气单胞菌感染,免疫保护率达到69%。本研究用纯化获得的嗜水气单胞菌P5蛋白免疫小鼠制备特异性P5多克隆抗血清,抗血清被动免疫红鲫鱼,发现鱼体可有效抵抗嗜水气单胞菌的感染,较对照组免疫保护率达到显著的56%,可见,嗜水气单胞菌外膜蛋白P5具有较强的免疫原性和免疫保护功能。为进一步探究P5的抗原性,体外模拟试验显示P5抗血清与嗜水气单胞菌存在体外识别作用,表明P5也具有良好的抗原性。
体外模拟鱼血浆杀菌作用,发现嗜水气单胞菌进入鱼体后,血浆存在非特异性的杀菌作用,P5蛋白表现为下调以抵抗血浆因子识别,该现象可能与所预测P5蛋白三维结构为桶状相关,推测P5可能通过关闭孔道以减少与血浆杀菌因子的结合,从而达到保护细菌的作用。
综上所述,嗜水气单胞菌外膜蛋白P5在气单胞菌间进化保守,二级结构以无规则卷曲为主,三维结构呈桶状。P5蛋白具有较强的免疫原性和抗原性,对红鲫鱼具有显著的免疫保护作用。此外,P5蛋白可有效抵抗鱼血浆对嗜水气单胞菌杀伤作用。本研究表明嗜水气单胞菌外膜蛋白P5是一种良好的保护性抗原,有望作为防治嗜水气单胞菌蛋白亚单位疫苗的候选抗原成分。
