重金属Cd胁迫对荻和芦苇种子萌发与幼苗生长的影响
2021-04-25魏媛媛谢思敏梁宇鹏
魏媛媛, 张 杰, 谢思敏, 梁宇鹏, 张 辰
(1.安徽师范大学 生态与环境学院,安徽 芜湖 241002;2.安徽省重要生物资源保护与利用研究重点实验室,安徽 芜湖 241000)
土壤是人类赖以生存的重要资源和根本所在。现代工农业的快速发展,带来了严重的土壤重金属污染问题[1]。重金属污染物具有隐蔽性、不可逆性、难降解性等特点,在土壤中不断沉积,甚至可以转化为毒性更大的烷基化合物[2],一旦被作物吸收,将影响土壤肥力及作物质量安全[3-5]。Cd是植物生长发育中的非必需元素,其毒性强、污染面积大,是最严重的重金属污染物之一。重金属Cd侵入人体后,经积累会损伤结缔组织及生殖系统,导致人体机能衰退,尤其影响儿童的生长及智力的发展[6]。大量文献表明,重金属会干扰种子萌发和幼苗生长时期的代谢活动和生理生化过程。例如,呼吸系统受阻、自身贮藏物质的可利用率下降以及光合作用功能下降等,从而抑制植物生长[7-8]。植物修复是利用植物吸收土壤或水中的重金属及其他污染物质,达到净化土壤或水质的目的,是一种成本低、绿色型、生态型的重金属污染土壤修复途径[9-11]。
在植物修复研究的早期,人们把目光主要集中在超富集植物上,但超富集植物存在种类少、生长速度缓慢、修复年限过长、生长区域受限等问题,极大地限制了对重金属污染土壤的修复效率[12-14]。为解决这一问题,人们开始将目光投向能源植物。能源植物具有光合作用效率高、生物量大、生长速度快、适应能力强、分布广泛等特点,在修复重金属污染土壤的同时,可提供生物质能,将修复工作与能源生产有机结合,完成重金属由粮食链向能源植物链的迁移,既能保证粮食作物安全生产,又能解决土壤环境问题,是一类具有发展潜力的修复性植物[15-17]。薛忠财等发现能源植物甜高粱对Cd有较强的吸收能力和耐性,且Cd处理不会对甜高粱生产燃料乙醇造成影响,同时带来了环境效益与经济效益[18]。Zhang等对铜尾矿上生长的能源植物进行研究,发现荻、芒、芦苇、芦竹均能够提高铜尾矿中重金属有机结合态的比例及土壤酶活性,其中荻是一种极具修复潜力的能源植物[15]。
荻[Miscanthussacchariflorus(Maxim.)Benth.]和芦苇[Phragmitesaustralis(Cav.) Trin.ex Steud.]均属于多年生禾本科植物,分布广,根茎发达,株高,叶茂,耐寒,抗旱,生长迅速,生物量大,生长发育习性相近,常常相伴生于有水源的空旷地带,是我国河岸湿地防土固堤的先锋环保型植物。也是尾矿废弃地、重金属污染区域中改良土壤环境的首选物种。有人研究过Cd胁迫对荻萌发的影响[19-20],但Cd胁迫对荻和芦苇萌发机制的差异性尚未可知。萌发时期的种子对外界环境变化更敏感且要求较高,通过探究重金属Cd胁迫对荻和芦苇种子萌发与幼苗生长特性的影响,比较两种植物对重金属Cd的耐受能力,能够在一定程度上反映植物对重金属的耐受能力[21]。本研究以荻和芦苇种子为实验材料,通过萌发试验比较不同浓度Cd胁迫下荻和芦苇种子的发芽指标、生长指标、酶活性、光合色素含量及叶片组织的电导率,进而分析比较荻和芦苇两种能源禾草对重金属Cd的耐受能力,为能源植物修复Cd污染土壤提供一定的理论参考。
1 材料和方法
1.1 试验材料
供试种子于2018年11月下旬采自安徽芜湖青弋江江边,经自然风干,剥离颖壳后选取籽粒饱满的种子保存备用。用分析纯CdCl2·2.5H2O提供Cd2+,设置Cd2+处理浓度分别为1、5、10、15、25、50mg·L-1,每个处理组设置6个重复,以蒸馏水对照组,记为CK。
1.2 试验设计
萌发试验开始于2019年10月8日,地点在安徽师范大学生态学实验室。分别取荻和芦苇种子,浸种24h后,用5.2%的NaClO溶液消毒10min,再将种子用去离子水冲洗数次,用滤纸吸干种子表面水。在直径为8.5cm的培养皿中铺2张滤纸,用不同浓度的Cd2+处理液将滤纸完全浸湿,以蒸馏水为对照(CK)。每个培养皿中均匀地放入种子120粒,放置于光照/黑暗为12h/12h,温度为28±1℃,光照强度为80%的培养箱中,每12h向培养皿中分别加入相应浓度的Cd处理液5ml,以保证种子萌发所需温度和水分[19-20]。
1.3 指标测定
1.3.1 发芽指标的测定 每天同一时间记录种子的萌发数、幼根达种子长,幼苗达种子1/2长计为发芽。根据下列方法分别计算荻和芦苇种子的发芽率、发芽势、发芽指数、活力指数。
发芽率(%)=(前7天内萌发的种子数/供试种子数)×100%;发芽势(%)=(前4天内萌发的种子数/供试种子数)×100%;发芽指数=ΣGt/Dt(Gt第t天的发芽增值数,Dt相应时间的发芽天数);活力指数=种子发芽指数×苗高。
1.3.2 生理生化指标的测定 待播种6天后,用3,5-二硝基水杨酸比色法,测量淀粉酶活性,Folin-酚法测定蛋白酶活性,酸碱滴定法测定脂肪酶活性;13天时用80%的丙酮测定光合色素的含量;15天时用电导法测定叶片组织电导率。
1.3.3 生长指标的测定 待播种12天时,用精度为0.01mm的游标卡尺测定幼苗的根长、苗高,用天马衡基FA2104万分之一电子天平测定植物鲜重。
1.4 数据处理与分析
用Microsoft Excel 2010进行数据处理,用SPSS 25.0进行单因素方差分析、差异显著性检验及相关性分析。
2 结果与分析
2.1 Cd胁迫对荻和芦苇种子萌发的影响
由图1-图4可知,随着Cd处理浓度的增加,荻和芦苇的发芽率、发芽势、发芽指数和活力指数变化规律一致,相同浓度Cd胁迫下荻的发芽率远高于芦苇(50mg·L-1除外)。二者的发芽率在Cd2+浓度为1mg·L-1时达最大值,分别为90.69%和54.44%。当Cd2+≥1mg·L-1时,2种植物的发芽率、发芽势和发芽指数均随Cd处理浓度的增大而下降。Cd2+≤15mg·L-1时,处理组与对照组(CK)相比,荻和芦苇的发芽率、发芽势和发芽指数无显著性差异(P>0.01);当Cd2+≥25mg·L-1时,与CK相比均呈显著下降趋势(P<0.01);Cd2+为50mg·L-1时抑制作用最大,发芽率、发芽势和发芽指数均达到最小值,各项指标分别为对照组的14.44%、9.85%、13.31%和52.90%、45.12%、50.91%,说明高浓度Cd处理对荻的抑制作用更强。Cd处理浓度与荻和芦苇的活力指数呈极显著负相关(P<0.01),Cd处理浓度为50mg·L-1时,荻和芦苇的活力指数分别为对照组的1.77%和16.30%。
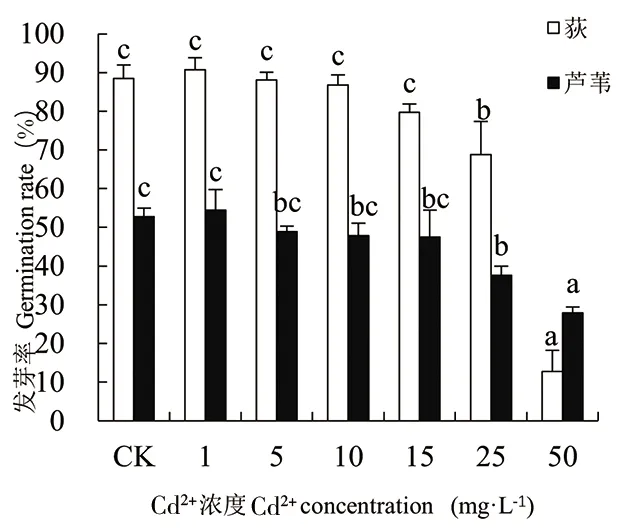
图1 Cd2+对荻、芦苇发芽率的影响
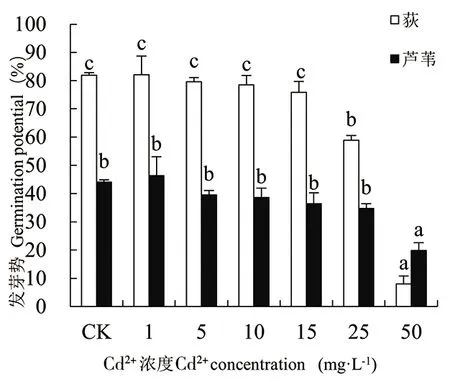
图2 Cd2+对荻、芦苇发芽势的影响
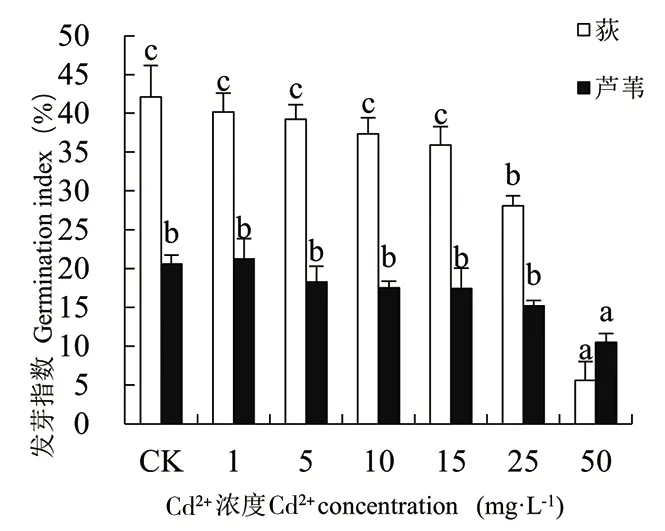
图3 Cd2+对荻、芦苇发芽指数的影响
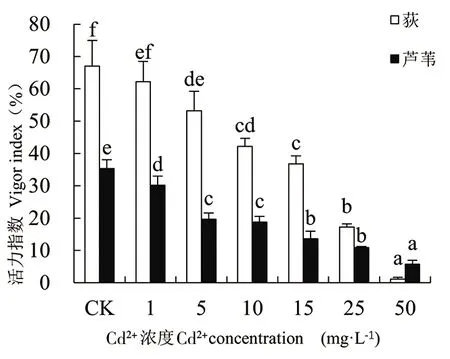
图4 Cd2+对荻、芦苇活力指数的影响
2.2 Cd胁迫对荻和芦苇幼苗生长情况的影响
由表1可知,随Cd2+处理浓度增加,对2种幼苗生长表现为抑制作用。Cd处理组的根长、苗高和鲜重均显著低于对照组(P<0.01)。低浓度处理下(≤1mg·L-1),幼根的生长情况优于幼芽,随Cd处理浓度的增加,对幼根的抑制作用增强,逐渐表现为幼芽的生长优势。Cd2+≥15mg·L-1时观察培养皿中荻和芦苇的生长状况,幼根长势较差且在突破种皮的数量上较低浓度处理时的减少量相当明显;当Cd2+浓度为25mg·L-1时,对荻和芦苇根长、苗高的抑制作用高达97.81%、61.01%和96.53%、58.14%,此时荻和芦苇生长状况基本上可视为只有胚芽没有胚根;当Cd2+浓度达50mg·L-1时,此时胚根被完全抑制。因此,在荻和芦苇幼苗的生长过程中,随Cd2+胁迫浓度增加,2种植物的生长受到抑制,且对幼根的抑制大于对胚芽的抑制。
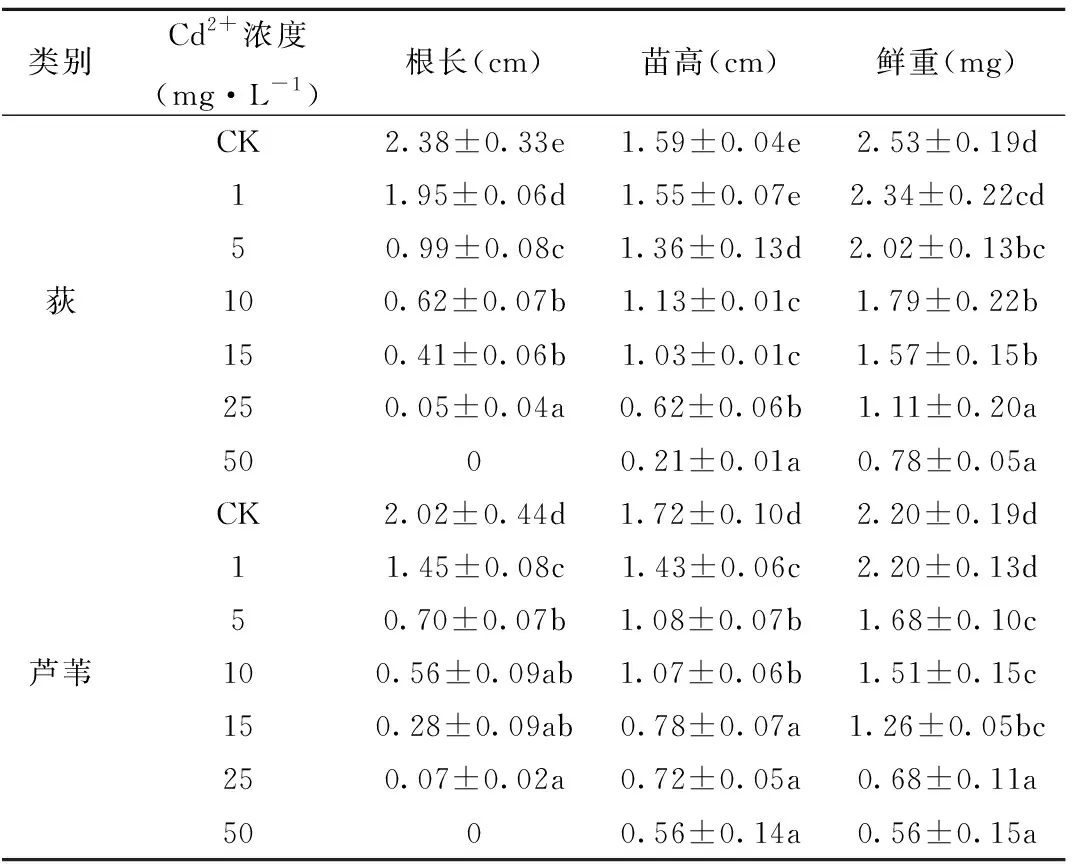
表1 Cd2+对荻、芦苇幼苗生长状况的影响
2.3 Cd对荻和芦苇萌发期间淀粉酶活性的影响
由图5可知,荻和芦苇体内α-淀粉酶、总淀粉酶活性均随Cd2+浓度的增大呈先上升后下降趋势,存在显著性差异(P<0.01)。荻和芦苇幼苗体内α-淀粉酶活性、总淀粉酶活性分别在Cd2+浓度为5mg·L-1和1mg·L-1时达到最大值,α-淀粉酶活性是对照组的1.51倍和1.16倍,总淀粉酶活性是对照组的1.21倍和1.41倍。Cd2+浓度为50mg·L-1时,淀粉酶活性最低,α-淀粉酶活性分别是对照组的38.47%和26.71%,总淀粉酶活性分别为对照组的67.03%和59.83%。结果表明,Cd胁迫对荻和芦苇体内淀粉酶活性起低促高抑作用,在Cd2+浓度由0~50mg·L-1范围内,荻幼苗体内α-淀粉酶活性变化较芦苇更显著。
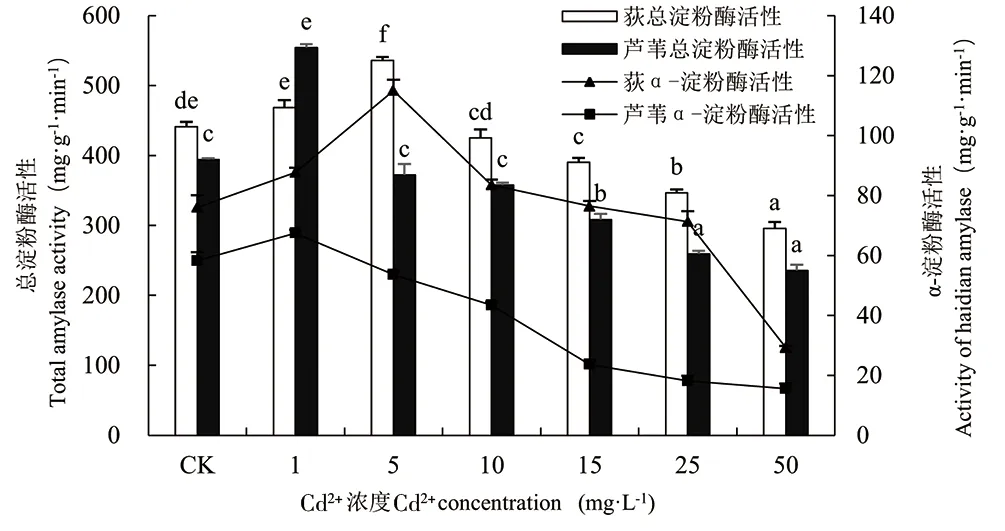
图5 Cd2+对荻、芦苇淀粉酶活性的影响Fig.5 Effect of Cd2+ on amylase activity of Miscanthus sacchariflorus and Phragmites australis图中相同植物不同小写字母表示差异显著(P<0.01)Different lowercase letters of the same plant in the picture indicate significant difference (P<0.01)
2.4 Cd对荻和芦苇萌发期间蛋白酶活性的影响
由图6可知,Cd胁迫对荻和芦苇幼苗蛋白酶活性产生不同的作用效果。荻幼苗蛋白酶活性随Cd2+浓度增加呈现上升后下降趋势,Cd2+浓度为1~10mg·L-1时,酶活性高于对照组,当Cd2+浓度大于10mg·L-1时,蛋白酶活性呈下降趋势,Cd2+浓度为50mg·L-1时,蛋白酶活性达最低值,但均无显著差异(P>0.05)。芦苇幼苗蛋白酶活性随Cd2+浓度增加呈先下降后上升再下降趋势,Cd2+浓度为1~15mg·L-1时,酶活性低于对照组,且存在显著性差异(P<0.05),Cd2+浓度为25mg·L-1时,蛋白酶活性达最大值,显著高于对照组(P<0.05),是对照组的1.44倍。
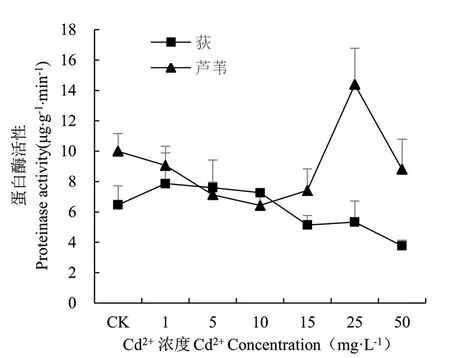
图6 Cd2+对荻、芦苇蛋白酶活性的影响
2.5 Cd对荻和芦苇萌发期间脂肪酶活性的影响
由图7可知,在一定的Cd胁迫范围下荻和芦苇幼苗脂肪酶活性变化存在一定差异。低浓度处理下,荻和芦苇幼苗脂肪酶活性变化规律一致,当Cd2+≤5mg·L-1时,随胁迫浓度的增加而增大,5~10mg·L-1区间内酶活性均呈下降趋势。Cd2+≥10mg·L-1时,2种植物幼苗脂肪酶活性变化存在差异。随Cd浓度增加,荻幼苗脂肪酶活性呈上升趋势,Cd2+浓度为50mg·L-1时,酶活性最高,存在显著性差异(P<0.05),是对照组的2.01倍;芦苇幼苗蛋白酶活性随Cd胁迫浓度的增大呈下降趋势,Cd2+浓度为50mg·L-1时酶活性最低,且存在显著性差异(P<0.05)。
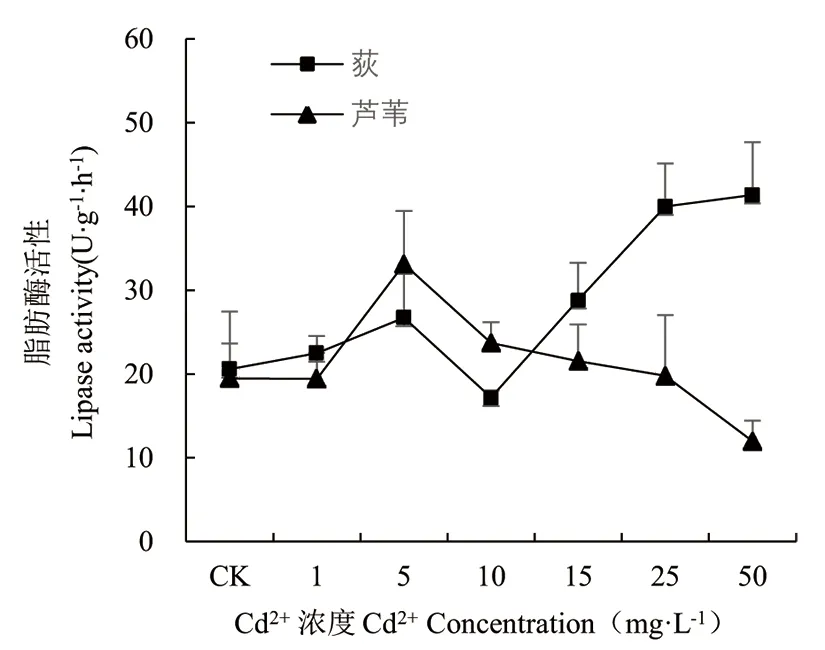
图7 Cd2+对荻、芦苇脂肪酶活性的影响
2.6 Cd对荻和芦苇幼苗光合色素含量的影响
由表2可知,随Cd2+胁迫浓度增加,2种幼苗体内叶绿素a、叶绿素b和叶绿素总量的变化趋势略有不同。随处理浓度增加,荻幼苗叶绿素a、叶绿素b及叶绿素总量均呈下降趋势,达到极显著差异水平(P<0.01);芦苇幼苗叶绿素a、叶绿素总量在Cd2+浓度为1mg·L-1达最大值,与对照组相比分别增加了5.78%、7.84%。3种色素含量均在Cd2+浓度为50mg·L-1时达最小值,类胡萝卜素的变化趋势不明显。低浓度的Cd胁迫已经对荻幼苗光合色素合成产生抑制作用,说明相同浓度梯度下,Cd2+对荻幼苗的迫害强度更大。
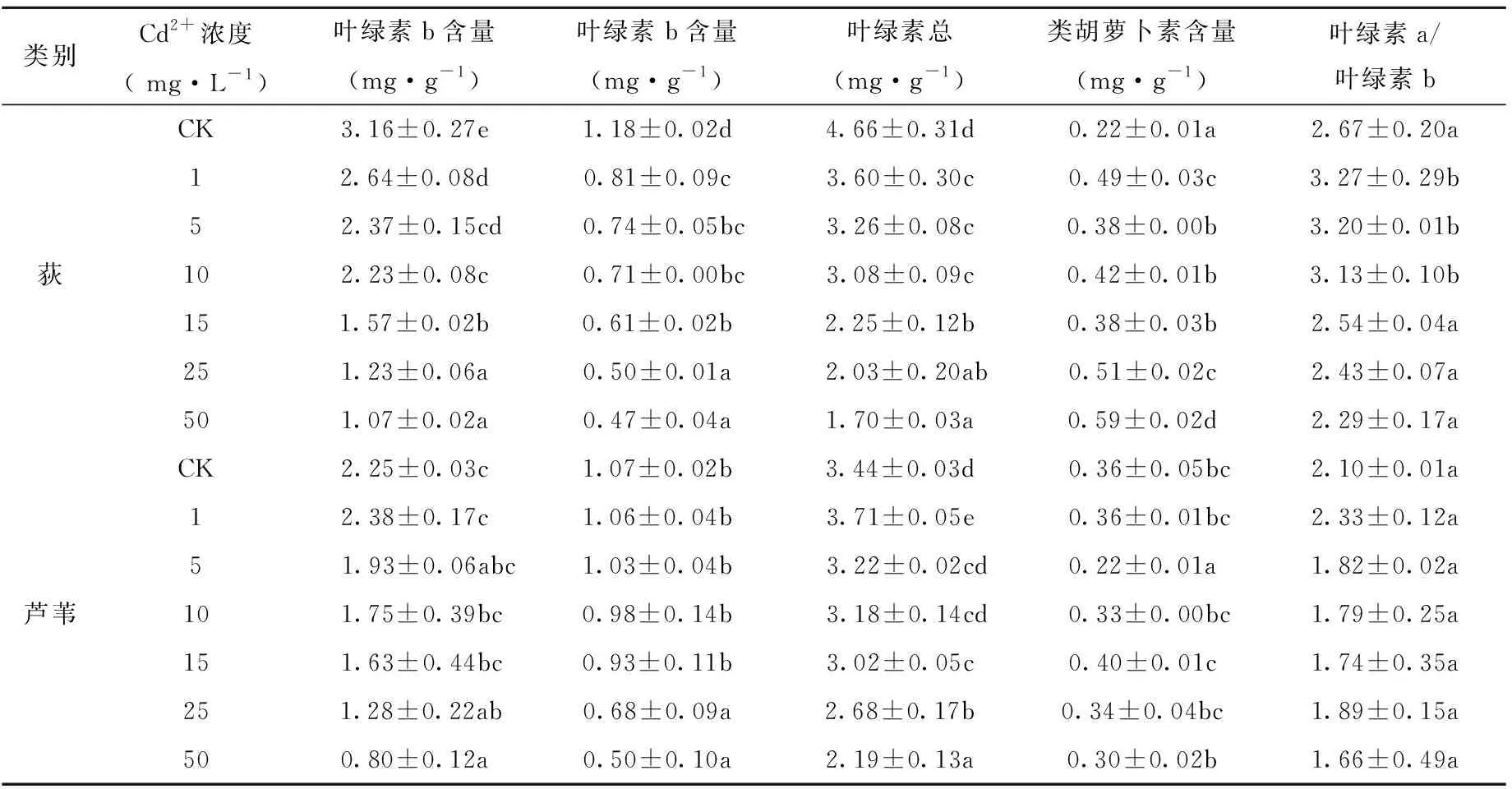
表2 Cd2+对荻和芦苇幼苗光合色素的影响
2.7 Cd对荻和芦苇幼苗叶片组织电导率的影响
随着Cd胁迫浓度的增大,荻和芦苇幼苗叶片组织相对电导率具有相似的变化规律,均呈显著增大趋势(P<0.01)。当Cd2+≥1mg·L-1时,荻和芦苇幼苗的电导率分别为对照组的1.09、1.67、1.91、2.14、2.37、2.39倍和1.55、2.12、2.59、2.90、3.32、3.42倍。结果表明,重金属Cd2+胁迫浓度越高,植物叶片受伤害的程度越大。当Cd2+浓度≥25mg·L-1时,2种植物叶片组织的电导率的变化不显著,说明此时重金属对植物叶片已经具有较高程度的破坏。
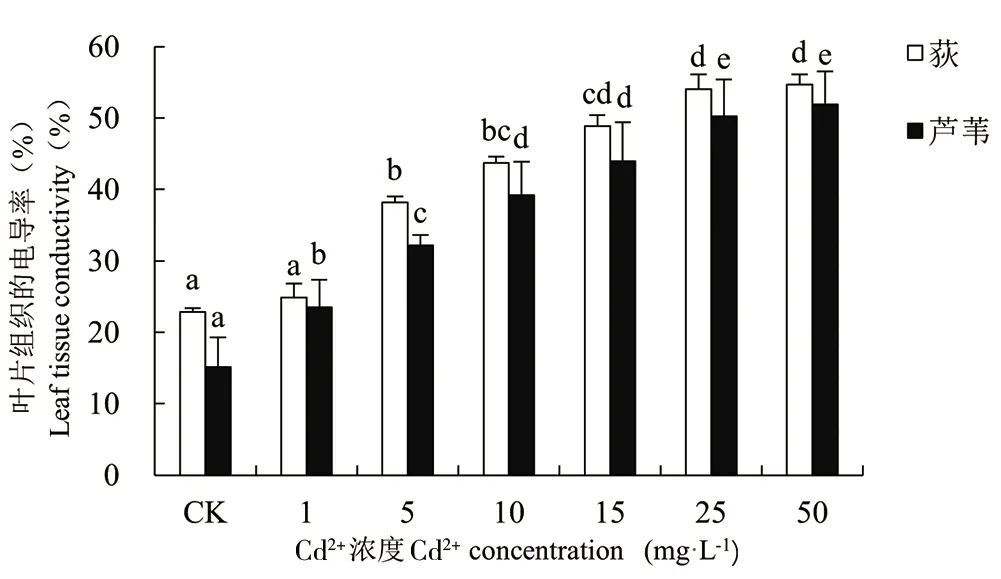
图8 Cd2+对荻、芦苇幼苗叶片组织电导率的影响Fig.8 Effect of Cd2+ on the tissue electrical conductivity of Miscanthus sacchariflorus and Phragmites australis
3 讨论
萌发期是植物生长发育过程中受外界非生物因素影响最敏感的时期之一,幼苗期对Cd胁迫产生的毒害反应尤为显著[23]。研究结果表明,重金属Cd对荻和芦苇种子萌发和幼苗生长均产生了一定的影响。种子的发芽指数、活力指数、根长、苗高、鲜重均随Cd2+浓度的增加而降低,这一结果与岑画梦、田如男等的研究结果一致[20,23]。当Cd2+为1mg·L-1时,荻和芦苇的发芽率最高,说明低浓度Cd2+(1~5mg·L-1)处理对种子萌发有促进作用,Cd2+≤25mg·L-1时,荻和芦苇种子能够正常萌发,说明两种植物对Cd2+都表现出一定耐性。高浓度Cd处理下,2种种子萌发受阻,部分原因是Cd影响了淀粉酶、蛋白酶、脂肪酶等的活性,从而抑制种子内贮藏淀粉和蛋白质的分解,降低了体内营养物质的利用率,影响了种子的萌发和幼苗生长[24]。Khadijah等研究Cd2+对豌豆种子萌发期胚胎生长的影响,发现Cd2+可以直接附着在蛋白水解酶的催化位点上,使种子内主要的贮藏蛋白的水解过程明显被破坏,游离氨基酸的释放量减少,导致胚胎缺乏必需元素氮的供应,最终影响种子的萌发[8]。本研究中,Cd胁迫下2种幼苗α-淀粉酶活性、总淀粉酶活性变化规律基本一致,对发芽率与总淀粉酶活性之间进行Pearson相关性分析表明,荻和芦苇的发芽率与淀粉酶活性之间存在显著的正相关关系,相关系数分别为0.561和0.759(P<0.01)。荻幼苗体内脂肪酶活性在Cd2+浓度为10mg·L-1时出现最低点,之后呈增大趋势;芦苇体内蛋白酶活性随Cd2+浓度的变化先降低后增加再降低。根据实验结果推测,高浓度的Cd2+并没有直接致种子死亡,外界的毒害延长了种子的萌发期,耐受力强的种子存活继续萌发,因此高浓度下脂肪酶、蛋白酶活性增加,耐受力弱的种子则直接死亡。除上述因素外,重金属Cd对两种植物的抑制作用还可能与两种植物自身的遗传特性及种子结构有关。
重金属进入植物体之后会结合到蛋白质的巯基上,干扰细胞中的酶活性、蛋白质的功能及细胞渗透调节系统,使某些必需离子无法被细胞吸收,对幼苗生长产生抑制[25-26]。本研究发现,随胁迫浓度增大,对2种植物的根长、苗高和鲜重抑制作用增强,当Cd2+浓度为50mg·L-1时,幼根被完全抑制,对苗高的抑制率分别达86.79%和67.44%,说明Cd胁迫对幼根的抑制作用大于对幼苗的抑制作用,与前人研究结果一致[15,21,26,27],处于萌发期的种子,胚根率先突破种皮,吸收周围环境中的水分及营养,胚根吸收大量Cd2+,受胁迫时间长,毒害作用大[28]。我们分析了荻和芦苇鲜重与叶绿素总量间存在极显著正相关关系(P<0.01),相关系数分别为0.924和0.669,说明Cd2+胁迫下叶绿素含量的高低与幼苗鲜重之间存在关联性。通过对荻和芦苇幼苗生长状况分析表明,荻较芦苇更易受Cd2+的毒害。
植物受到重金属或其他形式的胁迫时,体内产生大量的活性氧自由基(ROS)使细胞膜结构受损破坏细胞膜系统的完整性及通透性,造成大量胞内离子和有机物外渗,外界有毒物质流入细胞,扰乱了细胞内环境的稳定。细胞膜通透性的变化,是衡量植物抗逆性的生理指标之一,膜的通透性越大,相对电导率越高[29-31]。本研究中,随Cd胁迫浓度增大,荻和芦苇叶片组织相对电导率不断增大,表明Cd2+胁迫的浓度决定了细胞膜遭受破坏的程度。CK与Cd胁迫处理下,荻幼苗叶片组织的电导率始终高于芦苇幼苗叶片组织的电导率,可能是荻和芦苇细胞膜结构的自身差异性造成的。当Cd2+浓度≥25mg·L-1时,2种幼苗相对电导率的变化均不显著,高浓度Cd2+胁迫下细胞膜系统受损严重,继续增大胁迫浓度对相对电导率影响较小。
光合色素是植物进行光合作用的物质基础,其含量变化直接影响光合作用的强弱[32]。研究表明,重金属Cd能通过降低叶绿素的含量、改变叶绿体的超微结构、抑制与光合作用有关酶的活性等途径影响光合作用[33-34]。重金属进入细胞后,与酶蛋白的-SH基结合,干扰了有关叶绿素合成酶和CO2固定酶的活性,破坏了叶绿素的结构与功能,导致光合作用受阻[35]。也有研究指出,Cd胁迫下的叶绿体中有热休克蛋白(Hsp)的产生,这一蛋白对减少光合作用的损伤有一定的作用[36]。本研究中,芦苇在Cd2+为1mg·L-1时叶绿素含量达最大值,说明Cd胁迫促进了芦苇的光合作用。分别对2种幼苗叶绿素a、叶绿素b与Cd2+浓度之间进行相关性分析结果表明,叶绿素a与叶绿素b含量与Cd2+浓度之间均存在极显著的正相关关系(P<0.01),相关性系数分别为0.933和0.716,说明Cd胁迫对叶绿素a和叶绿素b影响相似。由前面实验结果可知,随胁迫浓度增大,膜通透性增加,经相关性分析表明,荻和芦苇叶绿素总量与相对电导率间存在显著负相关关系,相关系数分别为-0.792和-0.651(P<0.01)。由此推测,重金属Cd在改变细胞膜通透性的同时,可能会对叶绿体的膜结构也产生一定的影响,通过干扰叶绿素的合成,影响光合作用的过程;同时推测Cd对线粒体膜也会产生一定作用,影响植物的呼吸过程。Cd2+胁迫对2种幼苗呼吸作用的影响机制有待进一步探明。
4 结论
(1)高浓度的Cd(Cd2+≥25mg·L-1)胁迫显著抑制了荻和芦苇的种子萌发与幼苗生长,在Cd2+≤15mg·L-1时,对荻和芦苇种子萌发与幼苗生长的影响较小,无显著差异(P>0.01),说明2种植物对重金属Cd均有一定的耐受能力。
(2)通过对2种植物的萌发指标、生长指标、光合色素含量、酶活性和叶片组织相对电导率等试验结果进行分析,结果显示荻更易受到Cd2+的伤害,芦苇的耐受性更强。这为能源植物修复Cd污染土壤的治理提供了一定的参考依据。本试验仅针对萌发期的种子和幼苗进行分析,2种植物生长在土壤环境中对重金属Cd的抗性机理及修复效率还有待进一步研究。
