EFNA5 基因生理功能的研究进展
2021-04-21薛佳佳张国梁
薛佳佳 ,曹 阳,高 一,吴 健,李 旭,肖 成,张国梁,3,4,5*
(1.吉林农业大学动物科学技术学院,吉林长春 130000;2.吉林省农业科学院,吉林公主岭 130033;3.吉林坤成牧业科技发展有限公司,吉林公主岭 136100;4.农业农村部肉牛遗传育种重点实验室,吉林公主岭 136100;5.吉林省肉牛繁育及养殖技术科技创新中心,吉林公主岭 136100)
随着基因组学、分子生物学及生物信息学的发展,国内外学者对肝配蛋白A5(Ephrin A5,EFNA5)基因结构、调控机制以及生理功能方面进行了大量研究,主要围绕着人类癌症和神经性疾病方面展开。近年来EFNA5基因的研究开始向其他领域深入,多种新的生理功能被报道,哺乳动物EFNA5基因在细胞中调控多种细胞反应,如增殖、凋亡、分化、黏附及迁移等[1-3],参与晶状体发育及维持[4-5],作用机体胰岛素[6],调控雌性哺乳动物生殖过程[7]。最新研究发现,EFNA5基因能够参与脂肪产热,调控脂代谢[8],可能是肥胖症的治疗靶点,并且与经济动物肉品质性状相关[9]。因此,EFNA5基因在治疗人类肥胖症、胰岛素抵抗以及畜牧业提升畜禽肉品质性状等方面具有潜在的价值。本文综述了EFNA5基因结构、生物学功能、与癌症的关系及畜牧相关研究等方面,为挖掘EFNA5基因在生物机体上的重要功能提供思路。
1 肝配蛋白A5 所属家族概述
EFNA5基因是Ephrin 配体基因A 类的成员之一。该家族成员分为Ephrin 配体和Eph 受体两大类,由22个家族成员组成[10-12]。有研究人员在癌症和脑细胞中发现Ephrin 配体[13],科学家们利用统一命名法[10],根据序列的同源性以及在细胞膜上的结构排列顺序[11,14],将哺乳动物中的Ephrin 配体分为A 类和B 类,分别是EphrinA1-A5、EphrinB1-B3。与Ephrin 配体结合的促红细胞生成素产生肝细胞(Eph)受体于1987 年被首次克隆并鉴定[15]。研究人员根据Ephrin 配体与Eph 受体的结合偏好,将哺乳动物中的Eph 受体分为A 型与B 型,分别是(EphA1-A8)、(EphB1-B6)[10]。
如图1 所示,Ephrin基因在结构上,其A 类通过糖基磷脂酰肌醇(GPI)连接到膜上[16]。而B 类基因在胞内部分的结构是由PDZ 结构域以及相关的保守酪氨酸残基组成,而胞外部分则是由酪氨酸激酶结构域(TK)和受体结合结构域(RBD)组成[11]。
Eph 受体在结构上由细胞外成分和细胞内成分组成。Eph 受体的胞外成分包括富含半胱氨酸的结构域(Cys-rich domain),包括细胞因子结构域(Sushi)和表皮生长因子结构域(EGF)、配体结合结构域(LBD)以及纤维连接蛋白结构域(FN)[11]。Eph 受体的胞内部分由含有近膜的细胞质区域组成[17],该区域含有多个保守分子,如跨膜区(TM)、酪氨酸激酶结构域(TK)、无菌α基序(SAM)和PDZ 结构域组成[11]。SAM 结构域是Eph 受体结构中保守的分子域之一,但有趣的是它的破坏并不影响Eph 受体的生物学作用[18]。Eph受体通常利用位于蛋白C-末端的PDZ 结合序列与配体PDZ 结构域结合,形成聚类信号复合物发挥作用[11]。
2 EFNA5 基因表达与定位
EFNA5基因主要分布于细胞膜、细胞骨架和胞外的位置,主要在皮肤、脑、脾、肾脏、心脏和脂肪中表达,在其他组织中表达较低。
EFNA5基因位于人类5 号染色体,含有5 个外显子,编码序列长度为687 bp,可编码228 个氨基酸,EFNA5 还存在2 个转录本,分别编码188、201 个氨基酸。不同的转录本编码不同的氨基酸,它们在功能上也存在差异,在其他物种中也可能会出现新的转录本。在结构上,Ephrin 配体家族非常保守,EFNA5基因同其他家族成员一样通过一种被称为GPI 的蛋白质结构域整合到细胞膜中[19]。
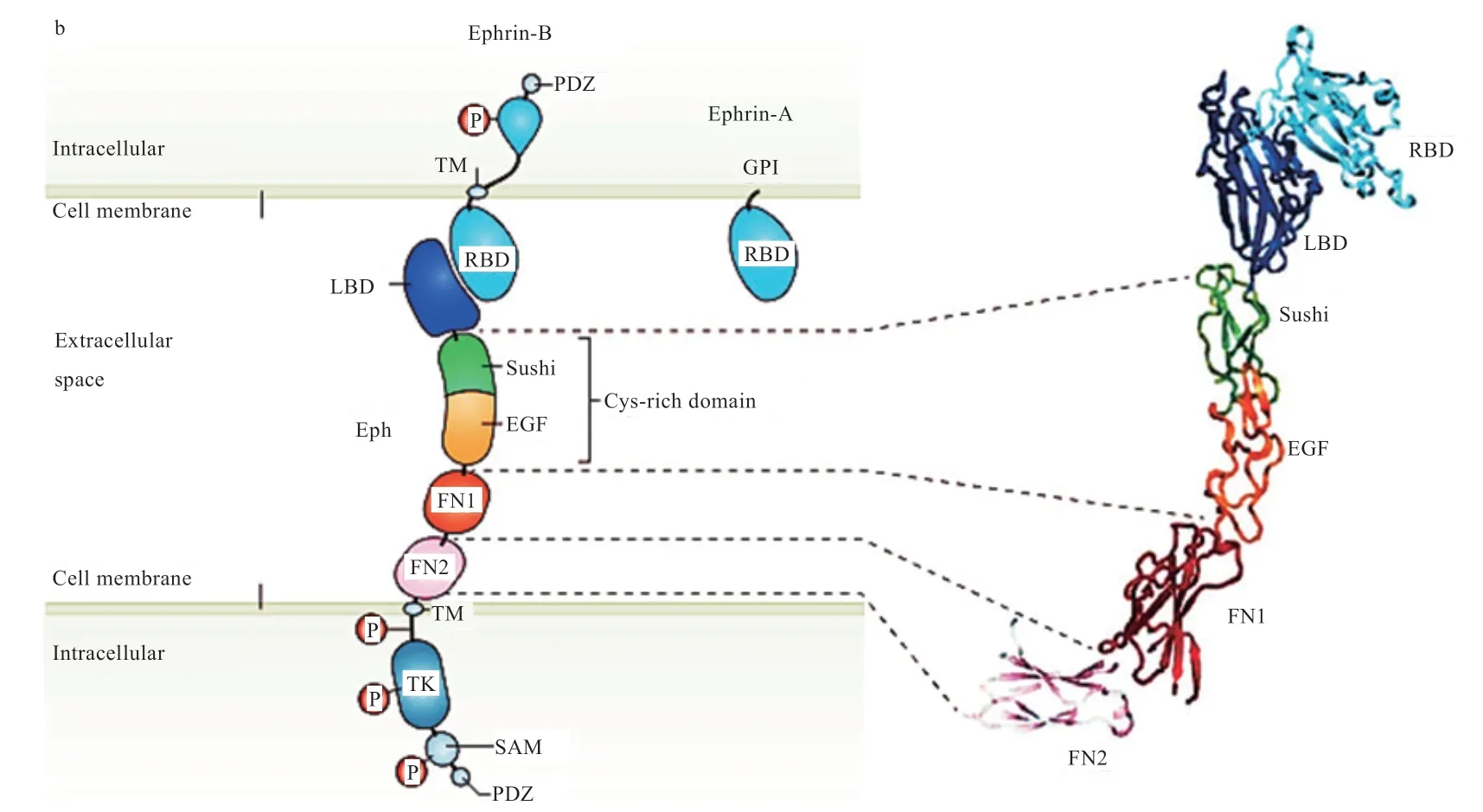
图1 肝配蛋白配体及肝配蛋白促红细胞生成素产生肝细胞受体的结构[9]
3 Ephrin-Eph 结合的信号传导
Ephrin 配体和Eph 受体蛋白均位于细胞膜上,其功能的发挥是通过细胞间接触来实现的[20]。Ephrin-Eph结合后能够发出多种信号模式来调控正常或病理的细胞生理过程。例如正向信号(Ephrin:Eph),Ephrin 充当信号的配体和发起者,Eph 是信号的接收者;反向信号(Eph:Ephrin),Eph 充当配体并将信号诱导到接收者Ephrin;双向信号传导,Ephrin 和Eph 同时充当配体和受体,这种信号传导模式很少发生[11]。
Ephrin-Eph 的信号转导通常被称为正向信号转导。在正向信号转导中,Eph 受体能够影响酪氨酸激酶受体(RTK)家族下游的一些接头分子及效应分子[21]。Eph受体的激活会导致一些酪氨酸家族成员的自磷酸化,包括SH 结构域(下游信号结构域)和其他激酶(如酪氨酸激酶Src 和Abl),以及没有受体角色的接头分子,如Nck 和富含半胱氨酸类受体蛋白激酶(Crk)[22]。Pasquale[23]研究表明,Eph 受体能够通过调节肌动蛋白影响Rho GTPase 成员进而来调节细胞形态改变细胞骨架并调控细胞黏附以及迁移,这一过程主要是将鸟苷二磷酸交换为受Eph 受体调节的鸟苷三磷酸来完成的。
Ehrin-Eph 家族通过反向信号转导途径调节多种病理和正常的细胞功能,在反向信号传递中,一些分子起关键作用,例如Src 家族激酶能够诱导EphrinB 磷酸化,从而为包括Grb4 在内的信号蛋白的SH2 结构域形成结合位点[24]。在反向信号中,另一个起关键作用的是PDZ 结构域,PDZ-RGS3 复合物将EphrinB 连接到G 蛋白偶联受体分子,并调节神经细胞迁移和干细胞更新,这表明PDZ 蛋白能够募集到EphrinB 的C-末端[25]。EphrinA5 在与其下游的Eph 受体结合时可与质膜相连,导致细胞室的形成,从而改变细胞的结构。这种反向信号被认为是由Fyn 蛋白激酶招募的,然后激活Sry 激酶家族蛋白,证实在大脑发育过程中Ehrin-Eph 的信号传递主要是反向的[26]。
4 EFNA5 基因的生理功能
4.1EFNA5参与神经发生、血管生成 EphrinA5 是已报道的能够调节成年脑细胞增殖的A 亚类Ephrin 成员之一。有学者研究表明,在颞叶癫病(TLE)小鼠模型中EFNA5和EphA4 之间的相互作用可激活ERK 和Akt 信号通路发挥调节作用,当ephrin/Eph 相互作用被干预后,能够抑制TLE 模型新生神经元生成以及微血管重塑[27]。在EFNA5功能丧失的研究中显示,在小鼠模型的海马齿状回中,神经元细胞增殖显著减少[28]。这些研究证明EFNA5基因在神经元生成以及血管生成中发挥至关重要的作用。
4.2EFNA5参与多种细胞反应 有研究发现,缺乏EFNA5的小鼠胚胎神经上皮细胞(NEC)凋亡减少;在平行实验中,EFNA5的异位过表达能够通过Caspase-3依赖性方式在体内引起NEC 大量凋亡,从而导致胚胎脑发育异常,表明EFNA5过度表达会引起细胞凋亡[1]。Worku 等[2]以小鼠卵巢颗粒细胞(GC)为研究对象,观察EFNA5对GC 凋亡、增殖、细胞周期进程及相关信号转导通路的影响,发现EFNA5是GC 中的促凋亡因子,能够调节雌性小鼠卵泡中GC 的增殖以及类固醇的生成,在卵泡的发生中起着重要的作用。Nguyen 等[3]研究显示,EFNA5配体在人原代骨髓基质细胞(BMSCs)中高表达,而造血干细胞(HSPC)的增殖、分化、黏附和迁移又依赖于BMSCs,当EFNA5-Fc 分子存在的情况下,HSPC 的黏附和迁移显著增加。这些研究表明,EFNA5在细胞增殖、分化、黏附、迁移以及凋亡等细胞反应中发挥重要作用。
4.3EFNA5调控雌性生殖 有学者研究发现,EFNA5在卵巢中表达,且缺少EFNA5的雌性小鼠不能生育,出现了许多卵巢缺陷,包括卵丘卵母细胞复合体在卵巢内释放,卵母细胞滞留在黄素化卵泡内的发生率增加,颗粒细胞和卵泡液血栓,纤维蛋白血栓和间质出血,表现出对促黄体生成素的不良反应,在超排后出现异常的卵巢组织学,表明EFNA5基因在雌性生殖中发挥着重要作用[29]。另有学者利用EFNA5的siRNA 处理小鼠颗粒细胞,然后进行转录组测序,探索EFNA5干扰后颗粒细胞的全基因组图谱,结果表明,与阴性对照相比,siRNA 处理的细胞中有包括EFNA5基因在内的208 个基因失调,它们主要富集在细胞外过程、钙结合和细胞黏附,参与了PI3K-Akt 信号转导途径、细胞因子-细胞因子受体相互作用、局灶性黏附和Rap1信号转导途径等过程,进一步干扰EFNA5基因表达能够影响参与卵泡发生的十多种基因的转录水平、调控网络以及生物学过程[7]。因此,EFNA5在雌性生殖中的调控作用可以为畜禽繁殖研究提供新的线索和研究思路。
4.4EFNA5维持葡萄糖稳态 Szepietowska 等[30]研究发现,EFNA5 能够激活邻近的β细胞中EphA5 受体,减少低血糖时的胰岛素分泌,在基础状态以及低血糖时,过表达EFNA5 能够刺激下丘脑腹内侧部(VMH)中的EPHA5 受体增加,能够暂时增加局部的谷氨酸浓度;敲除EFNA5则对VMH 间质液体谷氨酰胺浓度产生明显的抑制作用;在急性低血糖时,大脑葡萄糖敏感区VMH 内EFNA5 与EPHA5 的相互作用能够改变与胰岛素协同作用以恢复葡萄糖稳态,其调节作用可能是通过改变谷氨酸/谷氨酰胺循环来实现的。此外,在另一项研究中,EFNA5 在VMH 中的表达会受到前期反复低血糖的影响,而EFNA5受体在VMH 中的表达无明显变化,但通过靶向微量注射EFNA5 激活VMH 中EPHA5 受体,会使反复低血糖的非糖尿病大鼠的葡萄糖输注率降低[6]。据上述结果分析,EFNA5在脑内关键的葡萄糖敏感区VMH 内的作用可能是反复低血糖引起的胰高血糖素分泌障碍的关键原因之一。
4.5EFNA5动员脂肪产热 目前有研究表明EFNA5基因可能参与脂肪分化,并具有动员脂肪产热的作用。在徐露[8]的研究中,以野生型(WT)小鼠及ALDH1A1(-/-)型小鼠内脏脂肪(IAB)为对象,比较2 种不同细胞系间有关脂肪动员、产热及神经生长相关基因的表达差异,结果发现,ALDH1A1(-/-)脂肪细胞较WT细胞表这较高水平的PPARγ、PPARα、Ucp2 和EFNA5等同产热及神经生长相关的基因;在5 月龄的WT 及ALDH1A1(-/-)小鼠的饲养实验中发现ALDH1A1(-/-)小鼠较WT 小鼠血清中EFNA5含量高,且分化的ALDH1A1(-/-)脂肪细胞较分化的WT 细胞中EFNA5表达量更高;将喂养高脂饲料140 d 的小鼠随机分为PBS 注射组(n=6,100 μL)及EFNA5组(n=6,45 ng/mL,100 μL),隔天IAB 注射1 个月,冷刺激(4℃环境6 h)后比较2 组小鼠的代谢率,结果表明,注射EFNA5组小鼠的基础代谢率较PBS 组有明显升高;RT-PCR 检测10 例白人女性(5 例BMI<30,5 例BMI ≥40)的皮下脂肪组织中EFNA5的表达,结果显示同正常人群相比,肥胖女性体内EFNA5的表达较正常人群低[8]。这些结果证明,EFNA5在脂肪动员和产热方面发挥作用,这为人类肥胖症的治疗和畜禽肉质选育提供了新途径。
4.6EFNA5影响晶状体发育与维持 Noh[31]研究表明,在胚胎中使用α-Cre 转基因技术来诱导EFNA5在神经视网膜远端区域过表达会导致鼻部和颞叶视网膜细胞大量凋亡;除双极神经元外,分化的视网膜神经元的数量显著减少,整个视网膜大小,特别是鼻腔和颞叶区明显减小。这说明EFNA5的表达水平在调节神经视网膜中视网膜前体细胞池的大小中起关键作用。
Lin[4]对不同发育阶段的野生型和EFNA5(-/-)小鼠眼组织进行切片,H&E 染色观察,免疫组织化学检测蛋白表达和定位,结果发现EFNA5(-/-)小鼠出现永存性原始玻璃体增生症(PHPV),最明显的是在晶状体后方存在一个巨大的增生性肿块,并伴随小鼠一生,此外,EFNA5(-/-)小鼠眼睛内的肿块在胚胎和出生后阶段是有丝分裂活跃的。这说明EFNA5是调节原发性玻璃体退变的关键因子。
晶状体中的纤维细胞通过相互作用形成高度有序的结构以保持透明度。而Cooper 等[5]研究发现,与正常晶状体中的六边形外观形成对比,缺乏EFNA5基因的小鼠其晶状体纤维细胞在横截面上呈现圆形和不规则,且白内障在87%的缺乏EFNA5基因的小鼠中发生并发展。这一结果证明EFNA5是晶状体发育和维持的关键调节因子。
5 EFNA5 基因在癌症方面的研究进展
EFNA5基因与许多癌症的发生与发展相关。有相关研究显示,EFNA5在肝癌细胞中表达下调,并作为肿瘤抑制因子发挥作用[32]。Li 等[33]鉴定出EFNA5是miR-645 的直接靶基因,MiR-645 特异地靶向EFNA5mRNA 的3'UTR 并阻碍其表达,下调EFNA5抑制了结肠癌细胞迁移和侵袭的能力。同样的,在肝癌组织中miR-96 和miR-182 的表达与EFNA5蛋白水平呈负相关,miR-96 和miR-182 直接作用于EFNA5基因的3´UTR 区域,抑制mRNA 翻译,miR-96 和miR-182 通过下调EFNA5抑制肝癌细胞的增殖和迁移[34]。这些结果证明EFNA5基因与癌症密切相关,是人类癌症治疗的一个潜在靶点。
6 EFNA5 基因在畜牧研究方面的研究进展
目前,有报道发现EFNA5基因能够调控畜禽的经济性状。Yun 等[35]为深入研究在延黄牛脂肪细胞分化过程中的转录信息,在前体脂肪细胞(Day-0)、中分化期(Day-4)和成熟脂肪细胞(Day-9)上进行了RNA-Seq,结果发现EFNA5在成脂分化的第0~4 天差异表达,从分子生物学的角度为肉牛的选育提供了潜在的遗传标记。有研究人员在西门塔尔牛的3 个生长阶段(出生后6、12、18 个月)对西门塔尔牛的身高、体长、臀高、心脏大小、腹部大小和管骨大小6 个生长性状进行了全基因组相关联分析,发现包括EFNA5在内的多个候选基因,这些发现为进一步研究西门塔尔牛生长性状的潜在遗传机制提供了科学依据[9]。另外,EFNA5也被鉴定为影响肉鸡生长性状的候选基因[36]。另有研究发现,EFNA5在家禽卵巢中表达,且与低产个体相比,EFNA5在高产个体中具有上调趋势,表明该基因可能与家禽生殖过程有关[37]。因此,EFNA5基因有价值在畜牧领域深入研究。
7 小结与展望
综上所述,EFNA5基因在哺乳动物中具有非常重要的生理功能。它能够调控多种细胞反应,影响晶状体发育及维持,调控机体胰岛素代谢,参与雌性哺乳动物生殖过程。深入研究EFNA5基因的生理功能及其作用的分子机制,将进一步拓展人们对它的了解。目前EFNA5基因在畜禽经济性状上的研究只停留在初级阶段,但已有研究证明EFNA5基因可能是影响畜禽肉品质性状和生长性状的候选基因,从分子生物学的角度为畜禽选育提供了潜在的遗传标记。此外,EFNA5基因具有动员脂肪产热、影响脂肪代谢的功能,可能成为治疗肥胖症的潜在靶点。这为探究EFNA5基因在人类疾病方面的治疗以及家畜生产方面的选育提供了一定的理论依据和新的研究思路。
