红树林放线菌次级代谢产物的研究进展
2021-04-14贾舒涵郑红芸叶景静黄大林杨登峰黄庶识陈建宏蒋莲秀
贾舒涵,郑红芸,叶景静,黄大林*,杨登峰,黄庶识*,陈建宏,蒋莲秀
(1.桂林医学院基础医学院,广西 桂林 541004;2. 广西科学院 广西海洋天然产物与组合生物合成化学重点实验室,广西 南宁 530017)
0 引言
对于新的抗菌活性物质研制,可以通过化学和生物学等方法修饰结构,如氨基糖苷类(AG)修饰酶修饰、核糖体修饰、细胞膜的修饰等[3],来改善耐药性以及开发新的抗菌药物,或者发掘具有新型化学结构的天然生物抗菌活性物质,或者寻找药物作用的新靶位,或者通过细菌基因学的比对获得特异性靶标以此来增加特定微生物的抗菌活性[4]。
微生物资源是抗菌活性物质的主要来源之一。药用微生物主要包括放线菌、真菌和细菌,其中从放线菌中所发现的活性化合物最多( 约占70%),其次分别为真菌 ( 约占20%)和细菌(约占10%)[5]。但从放线菌得到的活性代谢产物,大部分都是来自于链霉菌属,主要原因是人们一直在陆地环境的范围内挖掘与筛选药用放线菌,使得研发新的活性物质以及抗生素出现了瓶颈。极端环境是未来发现未知放线菌资源以及新活性代谢产物的理想之地[6],极端环境包括高盐、高低温、高酸碱和高压之地,主要有冰川、沼泽、火山口、红树林等极端的特殊生境。而具有盐胁迫、高矿物组成、强还原性、强酸性、频繁的潮汐、强风、高温、强紫外辐射、缺氧污泥、寡营养等独特形态结构和生理生态的红树林生态系统,造就了性状独特、丰富和环境适应能力极强的放线菌种类,这些放线菌种类可以产生具有独特生物功能的次生代谢产物。
1 红树林放线菌
1.1 红树林概况
红树林主要生长于热带、亚热带海岸潮间带的滩涂浅滩上部,位于陆地与海洋的过渡带,长期受周期性海水浸淹,主要以真红树植物为主体的较为独特的木本生物群落,是最具特色的湿地生态系统,具有高湿度、高盐度、强酸和缺氧的特点[7]。
全球范围内的红树林大致分布在南、北回归线之间的区域,共分为两个群系:东方群系(东半球)和西方群系(西半球);我国红树林就属于东方群系。全球红树林以马来半岛及其附近的岛屿最为发达,但随纬度增加,红树植物的种类和数量会随之减少。根据世界粮农组织(FAO) 评估报告(2015),全球红树林面积 1.48×105 km2[8],分布于124 个国家和地区,其中亚洲拥有42%的红树林面积,非洲占20%,中北美洲占15%,大洋洲(澳火利亚、巴布亚新几内亚、新西兰、 南太平洋岛屿)占12%和南美洲占11%。迄今为止,印度尼西亚是最大的红树林国家,占全球红树林库存的26%~29%[9]。全球的红树林总体上呈现减少趋势,而在区域上中国红树林面积则有所恢复[10-12],我国红树林主要分布在包括广西、广东、海南、福建、浙江、台湾、香港和澳门等沿海省市,其中约92%位于广西、广东和海南三省区[13-14]。
全球红树植物种类属东半球最为丰富,含26 科32 属93 种,西半球较贫乏,仅8 科9 属17 种[15];我国红树植物种类为22科26 属38 种(不含外来种)[14],其中真红树植物13 科15 属27 种,半红树植物9 科11 属11 种[16]。
1.2 红树林放线菌
红树林是世界上生物多样性及基因多样性最丰富的生态环境之一,其特殊的生态系统,蕴藏着丰富的放线菌资源,也造就了红树林放线菌丰富的多样性。红树林放线菌广泛的分布于红树林湿地的动植物体表、体内、土壤及海底沉积物中,根据放线菌生长的生态环境,可将其分为三类:陆地放线菌、海洋放线菌及植物内生放线菌[17],这些放线菌主要有链霉菌属,小单孢菌属,马杜拉菌属,疣孢菌属,拟诺卡氏菌属等,以链霉菌属和小单胞菌属为优势菌群[18-20]。尽管近年来中国[21,22]、印度[23]、伊朗[24]、泰国[25]和马来西亚[26]等国家陆续报道从红树林特境中发现的新型放线菌,但从红树林分离到的放线菌种类仅为实际种类0.1-0.5%[18]。目前,文献报道放线菌种达2000 个,其中从红树林分离鉴定出放线菌纲下的11 科24 属[27],据不完全统计约有60 多种[27,28]。
2 红树林放线菌次级代谢产物及活性
2.1 红树林放线菌次级代谢产物
放线菌具有复杂的次级代谢系统,能够产生许多结构新颖、种类繁多的代谢产物[29]。目前发现大约10000 种天然抗生素中,约70%是由放线菌产生的[12-13],从放线菌发现的生物活性物质已经超过13700 余种,占已发现的33500 种天然活性物质的40%以上[30]。其中以链霉菌属产生的生物活性物质居多,产生了约 7600 种化合物,其余化合物由其它放线菌产生[15]。稀有放线菌也能代谢产生大量抗生素,包括利福平、西索米星、庆大霉素等,它们的分子量一般小于3000D,如紫色小单胞菌及其它小单胞菌产生的庆大霉素,糖多胞菌产生的红霉素,马杜拉放线菌产生的马杜拉霉素和洋红霉素,拟无枝酸菌产生的万古霉素,游动放线菌产生的游壁菌素,、诺卡氏菌产生的利福霉素等[31]。
放线菌产生的次级代谢产物具有丰富的结构多样性,造就它们多样的生物活性。放线菌产生的次级代谢产物主要包括抗生素类药物、抗肿瘤类药物、免疫抑制剂、降血脂类药物、抗虫药物。抗生素根据结构主要包括金霉素、土霉素、氧四环素等四环类;青霉素、头孢菌素等β-内酰氨类;庆大霉素、节那霉素、链霉素、新霉素等氨基糖苷类抗生素;红霉素、吉他霉素、螺旋霉素、麦迪霉素等大环内酯类抗生素;利福霉素等安莎类[32];万古霉素等糖肽类;制霉素、两性霉素B 等多烯类。抗肿瘤类包括阿霉素等蒽环类;苯丁亮氨酸等二肽类;紫杉醇等二萜生物碱类;博来霉素等糖肽类。免疫抑制剂类包括他克莫司大环内酯类免疫抑制剂[26];环孢菌素A 等环多肽类。降血脂类包括洛伐他汀等内酯类;普伐他汀等多元醇类。抗虫药物主要是阿维菌素、伊维菌素等。阿维菌素是从土壤中分离得到的链霉菌Streptomyces avermi-tilis MA -4680,从发酵菌丝中获得的天然产物,并命名为阿维菌素 (aver-mectins,AVM),阿维菌素为白色或微黄结晶粉末或淡黄色溶液,属大环内酯类抗生素;对阿维菌素结构修饰,获得伊维菌素[33]。
由于特殊的生态特性,红树林放线菌可以产生多种生物活性的次级代谢产物,具有抗菌等多种生物活性。Duddu MK 等人[34]从靠近印度东海岸的卡基纳达(Kakinada) 的辣木属(Koringa) 红树林沉积物中分离得到7 株放线菌,其中一种放线菌的分泌物对金黄色葡萄球菌(staphylococcus aureus)NCIM2079 和白色念珠菌(candida albicans) MTCC183 表现出优异的抑制活性。从马来西亚半岛丹戎马坡的红树林土壤中发现了三种新的稀有放线菌对NBRC 钙不动杆菌耐甲氧西林金黄色葡萄球菌和枯草芽孢杆菌都表现出抗菌活性[35];Ramesh等人从印度孟加拉湾不同地点采集的海洋样品中分离出放线菌208 株(链霉菌属占88%),其中111 株对人类致病菌有抗菌活性,151 株对两种植物病原菌有抗真菌活性[36];Suksaard 等[37]在泰国拉廊从水和沉积物中分离出 448 株放线菌,其中50 株具有促植物生长的放线菌中分别筛选出14 株对两种稻致病菌具有抑制作用和11 株具有氨基环丙烷羧酸盐(ACC)脱氨酶活性。Zhao 等[28]从广西北部湾茅尾海红树林放线菌分离出42 株放线菌中,以淀粉酶、蛋白酶、纤维素酶作为底物筛选出16 株具有酶活性,产淀粉酶活性菌株最多,其次产蛋白活性,产纤维素酶活性最少,其中的 GXN 151 号菌株 Micromonospora citrea 具有两种酶活性,其他的均只有一株酶活性。Liu[38]等从6 份不同来源分离出的96 株稀有放线菌中,有80 株菌对7 种靶标菌有抗菌活性,主要为马杜拉菌属和拟无枝酸菌属稀有放线菌;吴家法等[39]从红树林土壤样品中分离到 117 株放线菌中,有9 株链霉菌属,其的次级代谢产物对香蕉枯萎病病原菌有拮抗作用;Wang[40]等从广西北部湾的海泥样品和植物样品中分离的1 株链霉菌具有抑制香蕉枯萎病和金黄色葡萄球菌活性,并获得了星形孢菌素。
红树林放线菌产生的次级代谢产物有着广泛的抗菌、抗病毒、抗肿瘤、抗氧化等活性[27,29],表1 列出2015-2019 年期间,筛选出来源于红树林放线菌活性级代谢产物55 个,其中具有抗菌活性约22 个,细胞毒活性10 个,抗肿瘤活性16 个,抗炎活性4 个,抗关节炎和哮喘的各1 个,抗氧化活性1 个,酶抑制剂1 个,其中产生抗菌活性的次级代谢产物最多,其次是细胞毒活性。
产生于红树林放线菌的部分活性物质已在临床上得到应用,包括:四环素类抗生素兼具抗炎、抗凋亡和神经保护作用[41];泰乐菌素和尼达霉素等大环内酯类抗生素是世界上公认的畜禽支原体感染的预防和治疗的首选药物[42];唑烷酮类抗生素依哌唑胺 (eperzolid)和利奈唑酮(linezolid)、利奈唑酮的衍生物、四环素类抗生素的替加环素、喹诺酮类的奥泽沙星、β-内酰胺类的头孢菌素类的他唑巴坦等,具有较强的稳定性,同时还具有耐青霉素酶、过敏反应较少的特点,相比青霉素效果更好。
2.2 红树林放线菌次级代谢产物结构和功能
放线菌次级代谢产物从结构上可分为生物碱、聚酮类、环肽类、大环内酯类、肽类、黄酮类、香豆素类、哌嗪类和烯萜类[16]。大环内酯类是一类分子结构中具有12~16 碳内酯环,从功能上可分为抗菌、杀虫、植物生长调节、抗肿瘤活性及其他多种重要的活性类型,临床上主要治疗葡萄球菌和一些致病性流感嗜血杆菌、博代氏杆菌、棒状杆菌属、衣原体、支原体、立克次氏体和军团菌;肽类主要由糖基化的环状或多环非核糖体肽组成,其特征为存在一个与糖连接的独特长脂肪链,可作为抗生素、激素、抗癌药物、杀虫剂等应用于医药和农业等领域;黄酮类是一类存在于自然界的、具有2-苯基色原酮(flavone)结构的化合物,具有抗菌、抗病毒和抗肿瘤活性,临床上对HIV 病毒有较强的抑制作用、降低肿瘤细胞耐药性;香豆素类是邻羟基桂皮酸的内酯,香豆素的母核为苯骈α-吡喃酮,该类化合物的母核结构有简单香豆素类、呋喃香豆素类、吡喃香豆素类三种类型,具有抗菌、抗真菌、抗肿瘤活性;哌嗪类是化合物结构中含有哌嗪结构的一类化合物,具有抗菌、抗肿瘤活性,临床上具有酶抑制剂抗癌作用;烯萜类种类很多,放线菌主要是倍半萜类,分子中含三个异戊二烯单位的一类化合物,其中的环状倍半萜有青蒿素,穿心莲内酯,具有抗菌、抗病毒活性,临床上治疗肠道疾病,肝病,治疗疟疾。
近10 年来,红树林放线菌中产生次级代谢产物的研究上有较大突破性进展。李聆莉[43]等对我国南海红树林底泥分离出的链霉菌Streptomyces sp.H41-26 中获得放线菌素类化合物,即放线菌素D(1)、放线菌素X2(2)、放线菌素Xβ(3)和放线菌素Xα(4),它们的结构分别为actinomycinD:R1=R2=H;actinomycinX2:R1+R2=O;actinomycinXβ:R1=OH,R2=H 4:actinomycinXα:R1=H,R2=OH,对人致病性病源菌具较好的抑制活性,其中放线菌素D 和放线菌素X2对结核杆菌具有很强的抑制作用[44]。Maniceavelu Manimaran[45]等从印度泰米尔纳德邦Pichavaram 红树林土壤的分离筛选的链霉菌VITMK1中获得两个化合物,吡咯并[1,2-A] 吡嗪-1,4- 二酮和六氢-3-(2-甲基丙基),它们对变形杆菌(23mm)、副伤寒沙门氏菌(21mm)、金黄色葡萄球菌(20mm)和蜡样芽孢杆菌(20mm)几种病原菌的抑制作用最大,有较好的抗菌活性效果。Han等[46]在广东红树林芦苇根茎泥样来源的链霉菌Streptomyces sp.CHQ-64 突变株的次级代谢产物中得到杀粉蝶菌素类化合物piericidin F(60),该化合物对宫颈癌细胞(HeLa)、急性早幼粒细胞白血病细胞(NB4)、腺癌人类肺泡基底上皮细胞(A549)和非小细胞肺癌(H1975)等多种肿瘤细胞都表现出较好的活性。从红树林土壤中分离到的链霉菌属放线菌的次级代谢产物中分离出来的多肽类放线菌素D 是最早用于癌症治疗的抗生素[47],包括后来人们陆续发现的丝裂霉素、盐孢菌素等活性成分均具有抗肿瘤活性。王丽[48]从福建省红树植物根际的26 份土样中分离筛选到的136 株放线菌,NRPS 基因检测中有55 株放线菌具有合成非核糖体多肽合成酶能力,卤化酶基因检测中有18 株放线菌具有卤化酶能力,这73 株放线菌可能具有产生新的卤代化合物的能力,卤化酶具有潜在的抗癌能力。宋现芹[49]等从红树林沉积物中分离得到一株放线菌 Strepomyces costaricanus SCSIO ZS0073,从中获得3 个单体化合物分别鉴定为放线菌素D、放线菌素XOβ 和一个多烯类化合物制霉色基素(fungichromin);13CNMR 谱显示,放线菌素D 结构中的δ165~175 之间有12 个碳吸收,在δ34~40 之间有4 个-CH3 碳的吸收;放线菌素XOβ 有62 个碳,比放线菌素D 多一个氧原子,脯氨酸4 号位的氢被羟基取代;制霉色基素有35 个碳,其中包含10 个烯碳,1 个酯基碳,化合物的紫外吸收光谱显示在300~400nm 范围内有3 个连续的大峰,是多烯类抗生素的典型特征,具有抗真菌活性。Chen[50]等从中国红树林白骨壤根际土壤中筛选出链霉菌属的一个黑色链霉菌新种452T 菌株,提取得到二酮哌嗪类化合物,对人癌细胞系A549(肺癌)、HCT-116(结肠癌)和HepG2(肝癌)具有显著的抗肿瘤活性。陈名洪[51]等从福建三个红树林保护区样品中分离到的318 株放线菌,有46 株疣孢菌属菌株为潜在的新菌株,从有些菌株中筛选出潜在的烯二炔类(Enediyne)抗肿瘤抗生素,结构独特,抗肿瘤作用机制特异且强效,是迄今为止发现的抗肿瘤活性最高的天然化合物。Feng[52]等从海南三亚红树林土壤中分离的149 株放线菌中,发现一株稀有放线菌为马杜拉菌属的潜在新种,对白色念珠菌和黄瓜炭疽病菌都具有抗菌活性,进一步研究表明该菌株具有抗细菌、真菌、肿瘤等活性[53]。Chandrananimycins A~C 来源于海洋马杜拉属放线菌,具有抗结肠癌、黑瘤、肺癌、乳腺癌等肿瘤细胞的活性,它们都具有吩噁嗪-3 酮骨架,chandrananimycins C 还具有强抗细菌、真菌及藻类活性[54]。

表1 红树林放线菌次级代谢产物活性、来源以及成分(2015-2019 年)
红树林生态系统的多样性,其次级代谢产物在结构类型和功能方面表现出与陆生放线菌不同的特点。按结构主要包括生物碱、萜类、香豆素、异香豆素、色酮、内酯、蒽醌类、大环内酯类等。抗霉素A1a/b(1,2)、抗霉素A2a(3)、抗霉素A9(4)、放线菌素X2(5)、放线菌素D(6)、制霉色基素(fungichromin)(7)、Reedsmycins A(8)、 gephyyamycin(9)、 酚 醛 类Phenol, 2,4-bis (1,1-dimethylethyl)-(10)、 生 物 碱pyrazine-1,4-dione,hexahydro-3- (2-methylpropyl)-(11),7,8-Epoxylanostan-11-ol,3-acetoxy(12)、生 物 碱Dihydroergotamine Mesylate(13)、增 塑 剂1,2-Benzenedicarboxylic acid, diisooctyl ester(14)、增塑剂Phthalic acid, diisodecyl ester(15)等不同结构化合物具有抗菌活性。
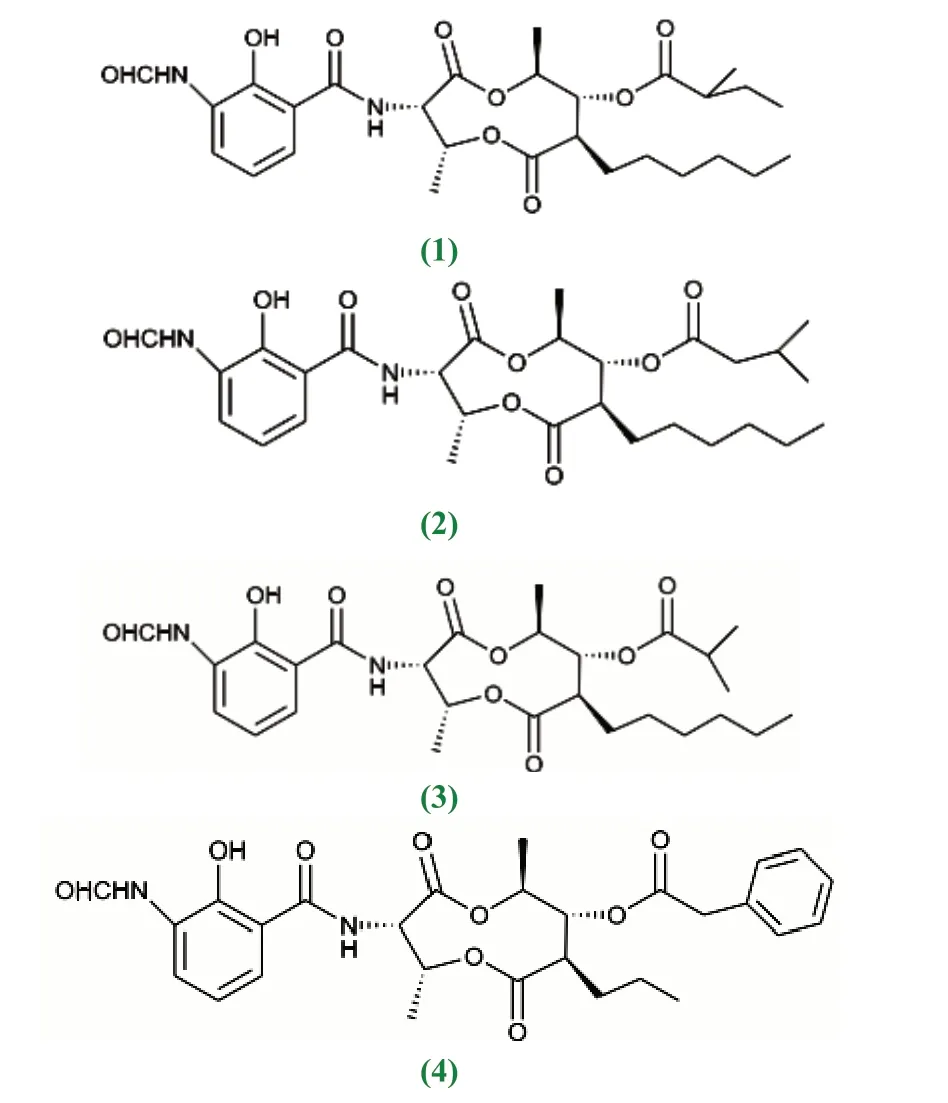

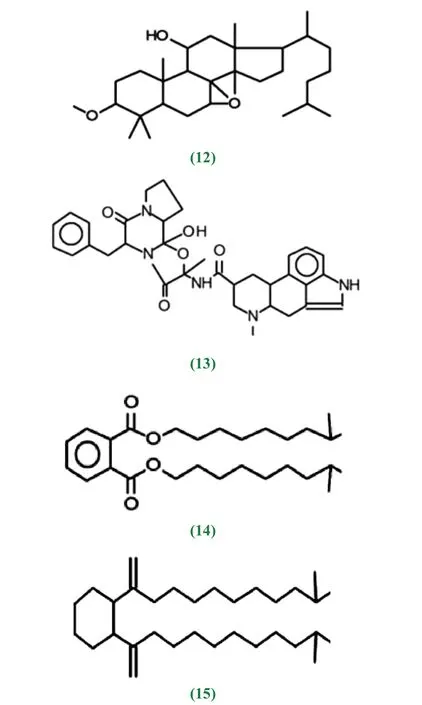
酚醛类Phenol, 2,4-bis (1,1-dimethylethyl)-(10)、生物碱pyrazine-1,4-dione, hexahydro-3- (2-methylpropyl)-(11)、生物碱Dihydroergotamine Mesylate(13)等化合物也具有抗炎活性。类固醇类7,8-Epoxylanostan-11-ol, 3-acetoxy(12)同时具有抗炎和抗哮喘活性。
新抗霉素A (16)、新抗霉素B (17)、(2S,3R)-N-[3-(formylamino)-2-hydroxybenzoyl]-l-threonine propyles-ter(18) 等化合物具有细胞毒活性。
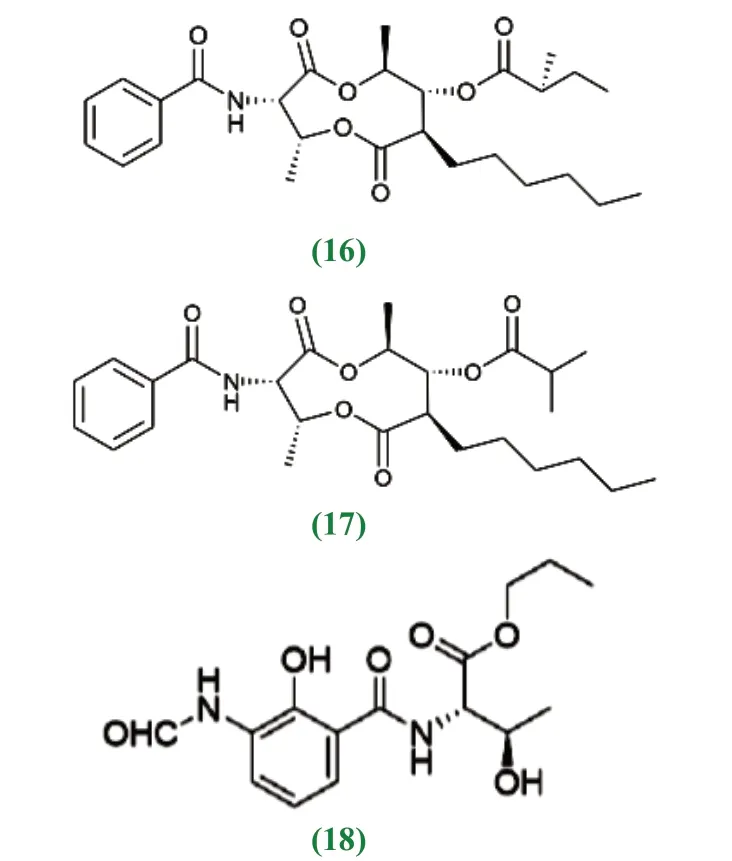
抗宫颈癌,腺癌和肺癌的杀粉蝶菌素类化合物piericidin F(19)、抗宫颈癌新生物碱类化合物(20)、Halichoblelide D(21)、Antimycins E~H(22)、抗 乳 腺 癌(2S,3R)-l-threonine, N-[3-(formylamino)-2-hydroxybenzoyl]-ethyl ester(23)、抗人前列腺癌醌类cysrabelomycin(24)、类固醇类化合物7,8-Epoxylanostan-11-ol, 3-acetoxy(12)等化合物具有抗肿瘤活性。
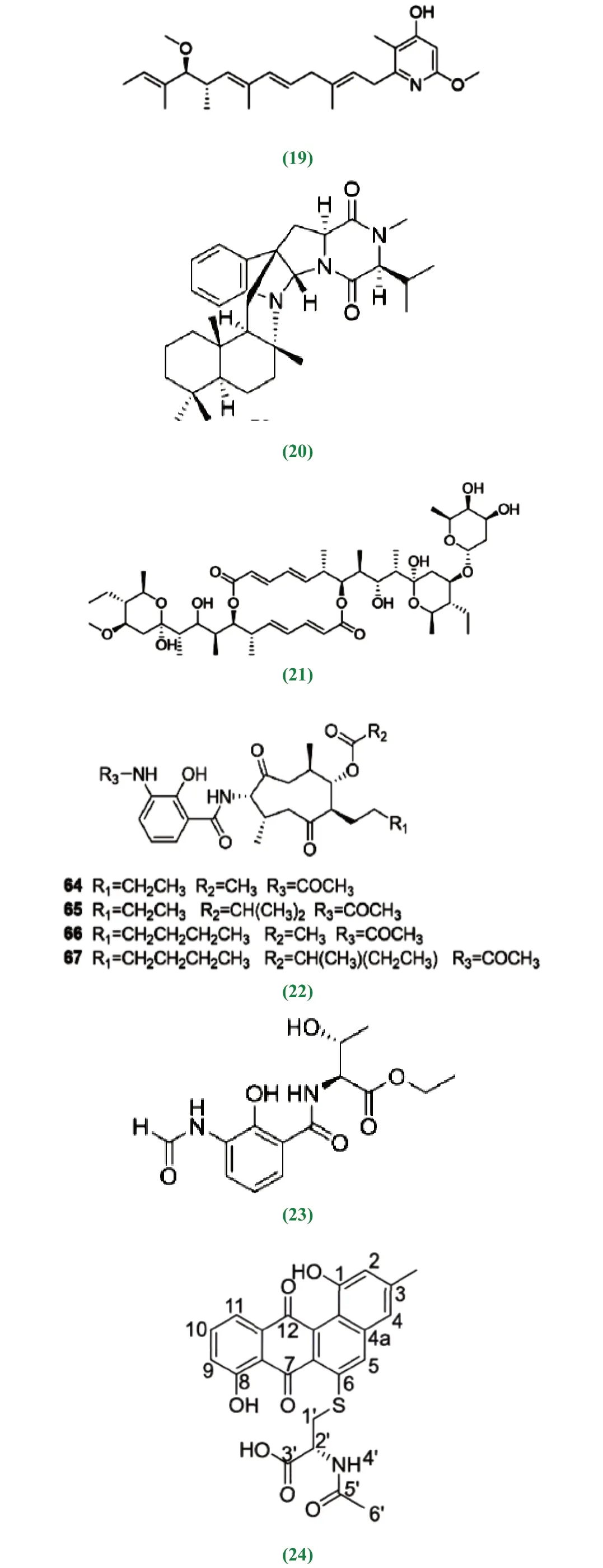
生物碱类化合物主要为含氮环状结构,有抗炎、抗癌等活性;类固醇类化合物不含结合的脂肪酸,主要由3 个六碳环己烷和一个五碳环结构组成,具有抗炎、抗哮喘、抗肿瘤等活性;抗霉素类化合物结构由独特的九元双内酯环母核与3-甲酰胺水杨酰胺通过酰胺键连接构成,具有抗真菌、抗虫、抗肿瘤及抗炎活性。酚醛类化合物主要结构是芳香烃环上的氢被(-OH)取代的一类芳香族化合物,具有抗炎活性。
3 展望
红树林具有丰富的放线菌资源,放线菌产生的次级代谢产物具有丰富的结构多样性和生物活性多样性。红树林放线菌也是发现新活性化合物的宝库,目前已在医药、农业等方面具有重要价值。尽管新的放线菌不断被发掘,但目前分离到的种类仍然很少,从红树林放线菌中重复发现产物,所以发现新化合物并筛选新型菌株是现代实验中一项非常棘手的问题。随着放线菌资源的深入研究,分离新菌株是发现新化合物的前提,通过筛选稀有放线菌菌种,能够发现更多新的生物活性物质和新的化合物。为了减少分菌重复率,提高新化合物的发现,针对筛选方法进行优化,利用微生物培养新技术以及高通量筛选技术等可以发现更多的稀有放线菌,提高药物筛选的有效性,相信未来稀有放线菌能在药物开发利用方面发挥更大作用。由于抗生素的不合理使用,多重耐药菌的出现,新药物的开发利用更为重要,但天然产物的有效化合物较低,筛选和纯化困难,可以通过放线菌发酵新技术和生物合成方法获得高产量的活性物质,并通过设计许多特异性的新药筛选模型,获得新颖、新结构活性物质。一些活性强的新骨架化合物,可以通过进一步修饰或者通过化学合成新技术方法,活动活性更强成本更低的化合物。
