Larger floods reduce soil CO2 efflux during the post-flooding phase in seasonally flooded forests of Western Amazonia
2021-04-14EmmaDOCHERTYandAndrewTHOMAS
Emma M.DOCHERTY and Andrew D.THOMAS
1Department of Geography and Earth Sciences,Aberystwyth University,Aberystwyth SY23 3DB(UK)
2School of Geography,University of Leeds,Leeds LS2 9JT(UK)
ABSTRACT Seasonally flooded várzea forests of Western Amazonia are one of the most productive and biodiverse wetland forests in the world.However,data on their soil CO2 emissions,soil organic matter decomposition rates,and soil C stocks are scarce.This is a concern because hydrological changes are predicted to lead to increases in the height,extent,and duration of seasonal floods,which are likely to have a significant effect on soil C stocks and fluxes.However,with no empirical data,the impact of altered flood regimes on várzea soil C cycles remains uncertain.This study quantified the effects of maximum annual flood height and soil moisture on soil CO2 efflux rate(Rs)and soil organic matter decomposition rate(k)in the várzea forests of Pacaya Samiria National Reserve,Peru.The study was conducted between May and August 2017.The results showed that Rs (10.6—182.7 mg C m−2 h−1)and k(0.016—0.078)varied between and within sites,and were considerably lower than the values reported for other tropical forests.In addition,Rs was negatively affected by flood height(P <0.01)and soil moisture(P <0.001),and it decreased with decreasing river levels post flooding(P <0.001).In contrast,k was not affected by any of the above-mentioned factors.Soil moisture was the dominant factor influencing Rs,and it was significantly affected by maximum flood height,even after the floods had subsided(P <0.001).Consequently,we concluded that larger floods will likely lead to reducedRs,whilst k could remain unchanged but with decomposition processes becoming more anaerobic.
Key Words: C cycle,climate change,greenhouse gas,soil CO2 emissions,soil moisture,soil organic matter decomposition rate,tropical forest
INTRODUCTION
Containing around 40%of the Earth’s terrestrial carbon(C)stocks and having the world’s highest soil organic matter(SOM)decomposition rates,tropical forests play a significant role in the global C cycle(Carvalhaiset al.,2014).Changes in temperature and moisture will affect SOM decomposition and soil CO2emissionsviatheir influencing effects on soil gas diffusion,microbial and root activity,productivity,nutrient availability,and redox dynamics(Davidsonet al.,1998;Neheret al.,2003;Sottaet al.,2006;Clevelandet al.,2010).Because temperature is relatively constant in tropical ecosystems,soil moisture is one of the most important factors influencing soil microbial activity and SOM decomposition,and is a critical variable in models that predict changes in soil C stocks on site specific and global scales(Cook and Orchard,2008;Moyanoet al.,2013).The general relationship between soil moisture and microbial activity can be depicted using a parabolic curve with reduced activity at both extremes and highest activity at an intermediate soil moisture content,where the balance of gas diffusion and water availability is optimal (Chamberset al., 2004; Vincentet al., 2006;Moyanoet al.,2013).Despite a broad understanding of the effect of moisture on soil microbial processes,decomposition and soil respiration rates remain poorly quantified (Cook and Orchard,2008),and in most C cycle models,they are represented as simple empirical functions (Tayloret al.,2017).
In general,it is thought that the decomposition rates will slow down with episodic soil saturation as a consequence of anoxia,leading to reduced soil biological activity(Schuur,2001).However,this may not be the case in highly weathered tropical soils where weakly crystalline and reactive iron minerals allow dissimilatory iron reducing bacteria to oxidize SOM,contributing to high rates of anaerobic C oxidation under hypoxic/anoxic conditions(Dubinskyet al.,2010).The reduction of iron can also lead to enhanced soil phosphorous availability by reducing the affinity of iron for phosphorous.As phosphorous is frequently the limiting nutrient in tropical ecosystems(Clevelandet al.,2010),any increase in its availability might encourage microbial activity(Chaconet al.,2006;Cleveland and Townsend,2006;Liptzinet al.,2011).Additionally,leaching of dissolved organic matter(DOM)can significantly contribute to decomposition in tropical forests that experience frequent heavy rainfall (Cleveland and Townsend,2006).Leaching of DOM has been shown to sustain microbial activity as it moves through soils(Neffand Asner,2001).It is an important factor in negating nutrient limitations of decomposition and affected by organic matter solubility. Owing to the extraordinary diversity of biota in the tropics,significant variations in litter chemical traits exist,affecting DOM leaching,microbial activity,and SOM decomposition(Townsendet al.,2008;Wiederet al.,2009).Thus,DOM leaching is likely to be an important,but poorly understood,process affecting the C cycle in tropical ecosystems.
As the largest of the world’s tropical forest regions,Amazonia is recognized for the significant role it plays in the global C cycle,and as such has received a considerable amount of scientific attention(Malhiet al.,2014).However,the vast majority of previous research effort has concentrated on Eastern and Central Amazonia, on areas with infertile soils and areas where more frequent and intense droughts are expected over this century(Chamberset al.,2004;Quesadaet al.,2010;Gattiet al.,2014).On the other hand,Western Amazonia has distinctive climatic, ecological, and pedological characteristics compared to the rest of Amazonia.Wood productivity (Malhiet al., 2004), nutrient turnover(Phillipset al., 2004), and total net primary productivity(Aragãoet al., 2009)are all higher in the Western forests compared to those in the typical Eastern Amazonian forests.Lowland várzea forests occupy a large proportion of the Western Amazon,typically becoming flooded as a consequence of a surge in rivers of Andean origin for around six months of the year,during which the soils are submerged and nutrient-rich sediments are deposited(Junket al.,2012).River hydrological regimes in várzea forests influence several ecosystem processes,including the C cycle(Bodmeret al.,2014).Western Amazonia is predicted to experience a 3%—8%rise in total precipitation,a 9%/18%increase in the extent of mean/maximum flood events,and lengthening in the duration of flooding by up to three months by the end of the 21st century(Langerwischet al.,2013;Sorribaset al., 2016; Zulkafliet al., 2016). Despite the potential implications of these changes for soil C stocks and fluxes,to the best of our knowledge,no studies have investigated soil C cycle dynamics in the várzea forests of Western Amazonia.
The aim of the present study was to determine the impact of flooding and soil moisture changes on soil CO2efflux rate(Rs)and SOM decomposition rate(k)in the várzea forests of Western Amazonia.The underlying hypothesis was that in terms of both depth and duration,the areas experiencing more flooding would have lowerkandRsas a consequence of soil waterlogging(Moyanoet al.,2013).The relationship between volumetric soil moisture,k, andRsdepends on numerous interacting processes affecting microbial activity,gas diffusion, and solute transport through the soil pore space matrix(Cook and Orchard, 2008). In flooded soils,oxygen is rapidly depleted,thereby severely restricting aerobic heterotrophic microbial activity and gas diffusion.As soil moisture declines(with the decrease in river levels and increase in soil drainage), soil gas diffusivity and oxygen concentrations increase, facilitating microbial respiration and the movement of CO2to the soil surface,resulting in increasedRs(Moyanoet al.,2013).Therefore,peak values ofRstypically occur at intermediate soil moisture contents before declining as soils progressively become drier(Fig.1).However,kandRsare not necessarily affected by soil moisture levels in the same way because while gas diffusion under waterlogged conditions limitsRs,decomposition may still occur,particularly in soils with high levels of ferrous compounds,through microbial anaerobic metabolism(Liesacket al.,2000;Dubinskyet al.,2010).
MATERIALS AND METHODS
Study area
The study was carried out at Tacshacocha (74◦21′E,4◦52′N, 117 m above sea level), a 200-km2area located on the Samiria River within the Pacaya Samiria National Reserve(PSNR),Loreto,Peru(Bodmeret al.,2018).Extending over 2.08×106ha, the reserve forms a part of the largest expanse of várzea flooded forests in Western Amazonia.The reserve has relatively stable air temperatures(21—31◦C) with monthly mean temperatures varying no more than 1◦C across the year.Humidity(80%—100%)and annual precipitation(1 500—3 000 mm)are also consistently high with unpronounced seasonality and zero months when precipitation is below 100 mm(University of East Anglia Climatic Research Unitet al.,2017;Bodmeret al.,2018).However, seasonal rainfall in the Andes causes an annual surge in sediment-rich waters travelling downstream,leading to an increase in river levels within the reserve by 9.17±1.19 m between October(low water level)and April(high water level).In extreme years,the rise in river water levels can flood up to 95% of the reserve by up toca. 4 m for several months of the year(Bodmeret al.,2014).Because flooding is directly associated with river level fluctuations,it is important to note that a higher level of flooding also denotes longer flood duration.
Vegetation cover within várzea forests varies according to fluvial dynamics,affecting both forest structure and species richness.Vegetation types establish in accordance with flood tolerance and geomorphological history of the substrate and are in constant succession depending on recent levels of exposure to flooding, sedimentation, and erosion from riverbanks(Wittmannet al.,2004).

Fig.1 Schematic illustration of the hypothesized effects of hydrological dynamics on soil moisture conditions and resultant heterotrophic respiration:predicted control factor(a),subsequent soil conditions(b),and resultant soil C process rates(c).The relationship between volumetric soil moisture and heterotrophic respiration is the overall result of various interacting effects whose influences are in differing directions,such as those of diffusion limitations and solute transport.Consequently,a peak tends to occur at intermediate values,indicating optimal soil moisture(adapted with permission from Moyano et al.(2013)).Rs is the soil CO2 efflux rate.
Gleysols are typical soils in the region(Quesadaet al.,2011),owing to long periods of saturation during which high levels of ferric iron are reduced to mobile ferrous compounds(Driessenet al.,2001;Buolet al.,2003).The reduction and subsequent oxidation of iron has been shown to accelerate SOM decomposition and contribute to rapid C cycling in tropical soils,even under periodic anaerobic conditions(Hall and Silver, 2013), but there have been no studies on this process in the investigated region. Some peat and clayey peat soils containing high mean organic C concentrations of 33%have also recently been uncovered within palm swamp areas at Tacshacocha (Lähteenojaet al., 2012). However,soils within the investigated area generally remain largely unstudied and relatively little is known about them.
Experimental design
Data collection took place between May 29 and August 1,2017,during the transition between high and low water once the flooding had subsided. Nine 10-m2plots were established,with three replicates of non-flooded,low,and high flood height categories dispersed across the research area.Flood height categories were established based on clear watermarks left on tree trunks from flooding.Non-flooded sites did not flood in 2017,whereas low and high flood height categories flooded on average 8 and 63 cm above the mineral soil,respectively.Additionally,using watermarks left by a record flood in 2012,it was determined that each category experiences a mean difference in annual high water level of 54 cm.
Determination of k and Rs
Determination ofkwithin each plot followed the Tea Bag Index(TBI)protocol(Keuskampet al.,2013).Three pairs of Lipton rooibos and green tea bags were buried 8-cm deep in a triangle formation within each plot. Incubation periods lasted 43—62 d,after whichkwas calculated using the method described in Keuskampet al.(2013).
Soil CO2efflux was measured in each plot every five days between June 14 and August 1 using static closed respiration chambers(Davidsonet al.,2002;Pumpanenet al.,2004).Full details of the chambers and the method can be found in Thomas(2012)and Thomaset al.(2018).Within each plot,three chambers were placed approximately 5 m apart in a triangle formation(within 1 m of the tea bag pairs where possible), avoiding placement within 1.5 m of any trees with diameter at breast height(DBH)>5 cm(DBH was measured at approximately 1.3 m above the ground).Chambers enclosed 83 cm2of soil and ranged 454—695 mL in volume, depending on insertion depth. Two-way vent valves on the chamber lids ensured that any pressure differences between the chamber and atmosphere were minimal and rapidly equilibrated. Heat sinks mounted through the chamber walls ensured that the internal air temperatures were not elevated above the ambient temperature.In order to minimize the impacts of chamber placement onRs, in preliminary experiments, we determined the appropriate length of resting time between chamber placement and measurements. Accordingly, the chambers were placed in the ground in early morning between 7:00 a.m.—9:00 a.m.and left for at least two hours before taking the readings between 10:00 a.m.—1:00 p.m.In order to avoid differences because of spatial variations within the plots,the chambers were consistently placed in the same location within each plot.Each measurement cycle involved creating an artificial breeze above the chamber to remove any accumulated CO2,placing the lid onto the chamber,and extracting three air samples of 10 mL using a gas syringe immediately and after one and two minutes.Prior to gas extraction,the air within the chamber was mixed by gently pumping the syringe.Air sample CO2concentrations were determined using an EGM-4 infrared gas analyzer (PP Systems, USA). In order to correct the effect of any unnatural retardation in diffusion due to the accumulation of CO2inside the chamber, a diffusion correction factor was applied(for details see Thomas(2012)).Equations described in Kutzbachet al.(2007)were used to determine mass C flux from the diffusion-corrected changes in CO2concentration normalized to mean temperature and pressure during measurement.At the time of sampling,air temperature was recorded using a USB502 logger(Adept Science, UK), while soil temperature and volumetric soil moisture within the top 5 cm of mineral soil were measured using a multifunction thermometer probe and an SM150 soil moisture probe(Delta-T Devices Ltd.,UK),respectively.
Meteorological data collection
A static USB502 data logger at Tacshacocha logged the ambient temperature and humidity under the canopy every half hour throughout the data collection period.Precipitation measurements using a rain gauge were collected at 7:00 a.m.daily from June 16 to August 1,and measurements of river level(relative to sea level)were obtained from the nearest hydrological station at San Regis,located further down the Marañon River(SEHINAV,2017).
Vegetation and soil assessment
Vegetation inventories of all trees with DBH ≥10 cm were produced,whilst the number of saplings taller than 1.5 m but with DBH ≤5 cm was counted in all plots.Additionally,at five points within each plot,understory density(using a 3-m touch pole with 30-cm intervals)and canopy openness(Brownet al., 2000) were measured. The decomposition layer was defined as the dark decaying matter above the mineral soil.Thicknesses of leaf litter layers(consisting of recently fallen leaves)were determined using a ruler against a section cut into the soil.Flood heights were measured from markings on at least three trees within each plot in 2017 and 2012 when record water levels were recorded.
Triplicate soil samples were collected from each plot four times throughout the fieldwork at 5—8 cm depth from the surface of mineral soil next to the teabag sites using a small plastic tube equating to a known volume of 18.85 cm3.Samples were air-dried and their masses were determined for bulk density calculation(Rowell,2014).The triplicate samples were composited and pH was determined at soil:water ratio of 1:2.5 using a digital pH probe after shaking for 15 min(Rowell,2014).Total C,nitrogen(N),and C:N ratio were also determined using the composited sample in the laboratory using a Leco TruSpec total element analyzer(Leco Instruments Ltd.,UK).Lastly,total C stocks for the 8-cm layer were calculated for each triplicate sample using previously determined bulk densities and C contents.
Statistical analysis
Normality tests were carried out on all dependent variables using the QQ-norm plot and Shapiro Wilks test.Vegetation and soil characteristics were compared across flood height categories by one-way analysis of variance(ANOVA)andpost-hocTukey test if significant. Repeated measures ANOVA was conducted to test for differences inkandRsacross flood height categories.The repeated measures function was applied to resolve non-independencies and account for the variance due to repeats within each plot (teabag pairs and respiration chambers)and over time(forRsdetermination only).Residual plots were visually inspected to make sure that model assumptions were satisfied.When the analysis was significant, a Tukeypost-hoccomparison of means was ran to further evaluate the relationship between different flood heights.
In order to test the effect of variable hydrological controls onRs,we analyzed river level,precipitation,and soil moisture data.Firstly,linear regressions were used to test for relationships between soil moisturevs. flood height, river level, and precipitation separately. Flood height was the only variable found to interact with soil moisture,which is why subsequently,both bivariate and multivariate models for river level, precipitation, and soil moisture were fitted againstRsusing linear regression for the whole dataset,and then for each flood height category individually.
In order to determine the effect of other variables potentially influencingkandRs,linear regressions were run individually on each of the following variables:soil temperature, air temperature, pH, bulk density, total C, total N,C:N ratio,leaf litter thickness,decomposition layer thickness,soil moisture,river level,and flood height.Exhaustive linear regression was then undertaken on significant variables(excluding air temperature and flood height because of their interactions with soil temperature and soil moisture,respectively) in order to test all subset combinations and find the best explanatory model.Selection of the best model was based on the highestR2value with least variables for bothkandRsseparately.Models were then cross-checked through stepwise elimination by comparing maximal with reduced models in order to confirm that each variable was significantly informative for the model.Definitive models were checked to ensure that model assumptions were met.Finally,relative weights tests were applied to reveal the relative ability of each variable in explaining variance inkand Rs.All analyses were carried out in R(R Core Team,2016)using the lme4(Bateset al.,2015),lmertest(Kuznetsovaet al.,2016),multcomp(Hothornet al.,2008),leaps(Lumley,2017),car(Fox and Weisberg,2011),relaimpo(Gröemping,2006),and ggplot2(Wickham,2009)packages.
RESULTS
Climate conditions and vegetation and soil characteristics
Air temperature and relative humidity were relatively constant,averaging 24.7±2.1◦C and 96%±4.4%,respectively,from May 31 to August 1.Rainfall occurred on 13 of the 48 recorded days,with the mean daily rainfall of 4.4 mm and the largest single daily event of 45 mm(Fig.2).River levels dropped nearly 5 m during the study period(Fig.2).Soil moisture was not significantly(P >0.05)correlated with river level or precipitation,whereas flood height significantly(adjustedR2= 0.33,P <0.001) affected soil moisture,with the high flood height plots consistently maintaining significantly higher soil moisture compared to the other two flood height categories(Fig.2,Table I).
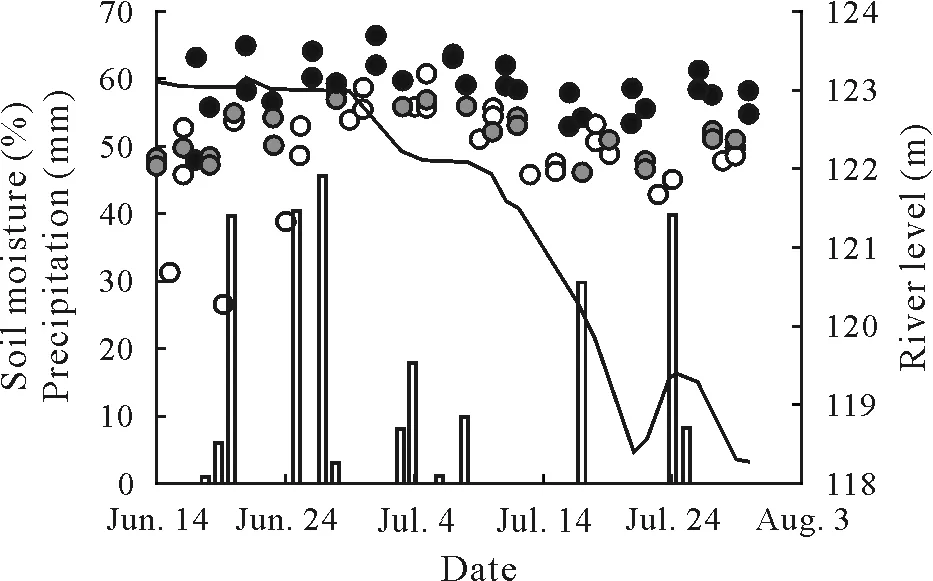
Fig. 2 Changes in Marañon river level (line), precipitation events (columns),and volumetric soil moisture(circles)within the top 5 cm of mineral soil across the three flood height categories of high flood(black circles),low flood(gray circles),and non-flooded(white circles)at Tacshacocha,Peru during the experiment period from June 14 to August 1,2017.
Accumulated DBH significantly increased as flood height decreased,whereas leaf litter layer thickness,decomposition layer thickness,and soil moisture at both 5—8 and 0—5 cm depths increased significantly with the increase in flood height. Additionally, soil pH and C:N ratio decreased as flood height increased, although neither were significant(Tables I and II).
Influence of flood height on Rs and k
There was no significant difference (P >0.05) inkacross flood heights(Fig.3a).However,flood height significantly(F2,26=6.79,P <0.01)affectedRs,with meanRsincreasing atca.10 mg C m−2h−1as flood height decreased(Fig.3b).Post-hoccomparisons revealed significant differences (P <0.001) inRsbetween the non-flooded and high flood categories,but not(P >0.05)between the non-flooded and low flood ones or the low and high flood ones.
When tested against flood height categories separately,river level was better at explainingRsthan soil moisture for both sites that were flooded (high and low flood heights)(Fig.4a,b,e,f).Conversely,for non-flooded sites,Rswas best explained by soil moisture(Fig.4c,g).When all flood height categories were combined,Rsshowed a significant positive relationship with river level but a significant negative relationship with soil moisture(Fig.4d,h).Overall,a combination of river level and soil moisture provided the best explanatory models forRsin terms of the adjustedR2value for all the flood height categories.
No significant relationships were found between any other variables tested(listed above in the statistical analysis section) andk. However,Rsshowed significant positive relationships with soil temperature,air temperature,and C:Nratio,and significant negative relationships with leaf litter layer thickness and decomposition layer thickness(Table III).

TABLE I Soil characteristics across flood height categories at Tacshacocha,Peru during the experiment period from June 14 to August 1,2017
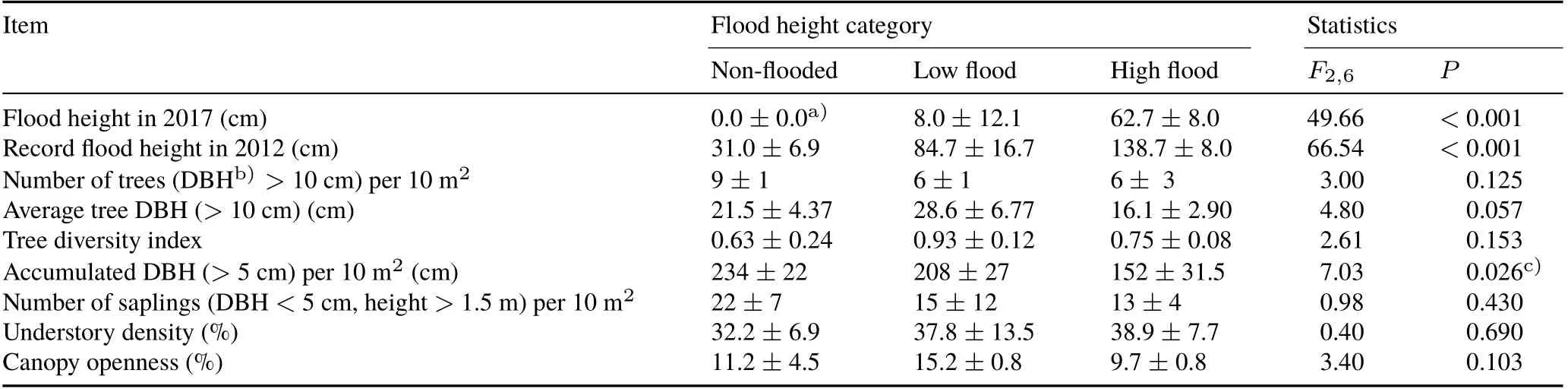
TABLE II Vegetation characteristics across flood height categories at Tacshacocha,Peru during the experiment period from June 14 to August 1,2017

Fig.3 Boxplots showing soil organic matter decomposition rates(k)(a)and soil CO2 efflux rates(Rs)(b)across the three flood height categories at Tacshacocha,Peru during the experiment period from June 14 to August 1,2017.Boxes show 25—75 percentiles,vertical lines show 10—90 percentiles,horizontal lines within boxes are medians(n=90),and points outside the boxes represent outliers.
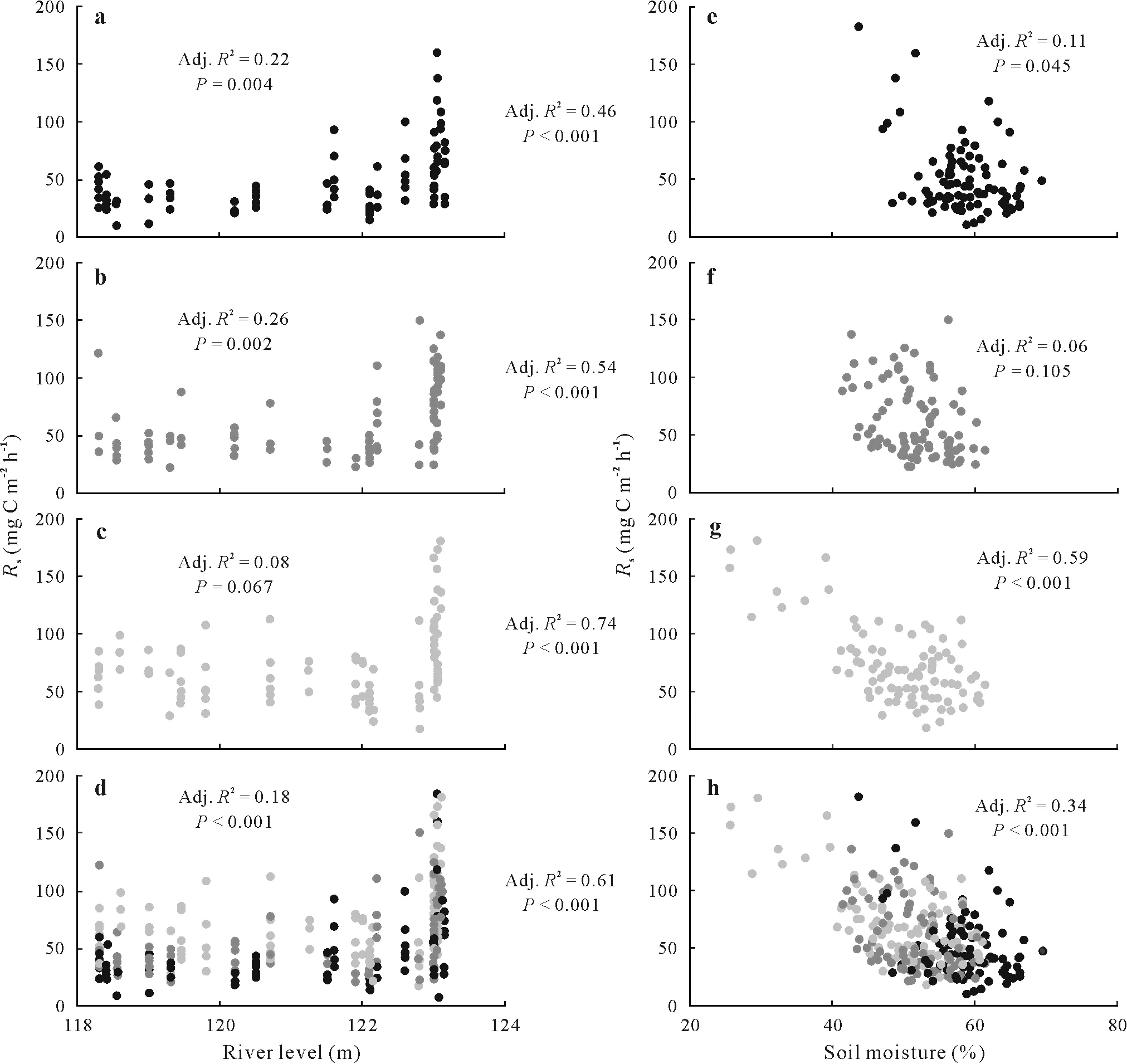
Fig.4 Relationships between soil CO2 efflux rate(Rs)and river level(a—d)and volumetric soil moisture(e—h)for the high flood(a and e),low flood(b and f),non-flood(c and g),and all the sites together(d and h).Adjusted R2 (Adj.R2)and P values are shown for individual bivariate analysis within each graph,while multivariate analyses combining the effects of river level and soil moisture on Rs are between the two graphs of each flood category.

TABLE III Linear regression outputs for significant influencing parameters vs. soil CO2 efflux rate(Rs)at Tacshacocha,Peru
Exhaustive linear regressions of significant variables revealed that a five-variable model(soil moisture,river level,soil temperature, decomposition layer thickness, and C:N ratio)provided the best explanation ofRs(adjustedR2=0.622,P <0.001).However,the comparison of maximal and reduced models showed that the soil temperature,decomposition layer thickness, and C:N ratio did not make significant contributions(P >0.05),leaving the definitive model(Eq.1)with soil moisture and river level as predictor variables(F2,87=69.25,adjustedR2=0.605,P <0.001):
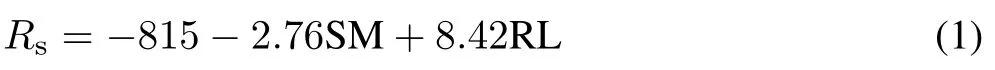
where SM is the soil moisture(%)and RL is the river level(m).Relative weights tests of the definitive model(Eq.1)showed adjustedR2of 0.388 and 0.226 for soil moisture and river level,respectively,indicating that the former was the most important variable when predictingRs.
DISCUSSION
TheRsat Tacshacocha varied greatly both between and within sites. Although the highestRsvalues in the present study were within the previously reported range of 146±76 mg C m−2h−1for tropical forests across the globe(Bond-Lamberty and Thompson,2010),the averageRswas consistently lower. However, theRsvalues in the present study were higher than previously reported rates of 12.52—20.75 mg C m−2h−1from Western Amazonia(del Aguila-Pasquelet al.,2014;Malhiet al.,2014).These comparisons suggest that there are large spatial and temporal variations inRsboth within Western Amazonia and between Western Amazonia and other tropical forests,implying that existing calculations could be overestimating the currentRsfor global tropical forests.Thekvalues were similarly variable but within the lower range observed in tropical forests with comparable mean annual temperature and precipitation(Tayloret al.,2017).
Changes in Rs and k with flood height
The significantly higherRsat the non-flooded sites than those at the flooded sites can likely be explained by a combination of interacting factors. The most important of them is the impedance of gas transport within the soil both during and after waterlogging through a combination of replacement of air within soil pores by water, damage of soil structure (Vincentet al., 2006), and clogging of pore spaces with fine sediment deposits. High levels of water at the soil surface could also create a soil-atmosphere barrier, further inhibiting the diffusion of CO2from the soil to the atmosphere (Kursar, 1989; Sottaet al., 2004).In turn, reduced gas movement limits oxygen availability for biological activity and consequently lowersRs(Dreyer,1994).Furthermore,an increase in vascular plant coverage associated with areas that tend to flood less could increaseRsas a consequence of increased root respiration(Malhiet al.,2014).There was a significant increase in accumulated DBH from non-flooded to high flood height categories,suggesting that root respiration could have been a contributing factor to the observed increase inRs;however,no other vegetation variables followed this pattern,indicating that root exclusion experiments would be necessary to demonstrate any influence onRsfrom root respiration at these sites.
In contrast,the lack of significant difference inkacross flood height categories could indicate that decomposition is continuing anaerobically under saturated conditions.Further analysis would be necessary to confirm this, and we suggest that this could be possible because of the likely high abundance of ferrous compounds indicated by the rust colour running through the soils (personal observations),accompanied by iron-reducing and oxidizing bacteria,which have been found to account for large proportions of SOM decomposition in similar soils with fluctuating redox conditions(Dubinskyet al.,2010;Hall and Silver,2013).Smaller dataset in comparison toRsin this study,and the variable nature of decomposition even within small distances(Prescottet al.,2000),could be contributing to the lack of observable differences inkacross flood height categories, but nevertheless it appears probable that decomposition processes and soil CO2emissions are decoupled,at least temporarily.Importantly,if a greater proportion of microbial decomposition occurs anaerobically as flood heights increase,CO2production could decrease and the production of other greenhouse gases, such as CH4, could increase (Liesacket al.,2000;Megonigalet al.,2004).
Rs and other hydrological controls
Firstly,it is important to note that flood height appeared to be the dominant influence on soil moisture,which is likely because of the lasting effect that frequent flooding has on soil structure and water-holding capacity (Vincentet al.,2006). The decrease inRswith increasing soil moisture was expected because of the well documented effect of saturation on both biological activity and the movement of gas through soils(Cook and Orchard,2008;Moyanoet al., 2013). However, this relationship was not as clear in areas that had undergone flooding during high water season.This can be explained,at least in part,because soil moisture varied much less in the plots that had been flooded than in those that had not, limiting the range over which the relationship was observable. TheRsat the flooded sites was,however,more strongly correlated with river level as compared with non-flooded sites.The fact thatRsdecreased as river levels receded at the sites that had flooded was unexpected and contradicted previous findings on seasonally flooded soils (Vincentet al., 2006). The most plausible explanation for this is that as river levels drop, DOM and dissolved minerals within the soil solution are leached out and become inaccessible for microbial activity (Neffand Asner, 2001).Furthermore, the initially highRscould be a consequence of the availability of nutrients in freshly deposited sediments,which would diminish throughout the terrestrial phase(Cook and Orchard,2008).It should also be mentioned that sediment replenishment from flooding might also be influencingRsthrough changing soil nutrient and mineral status or blocking soil pores (Cook and Orchard,2008);however,a detailed physio-chemical analysis of this soil was beyond the scope of the present study.
These results imply that a combination of hydrological factors, working in opposing ways, is influencing the variation inRsin várzea forests, at least in the transition from high to low water level.However,considering that a relationship between river level andRshas not previously been observed in this way,the complicated nature of these findings warrants further investigation over a longer period and across several seasons in order to unravel the complex interactions between flooding,soil moisture,and C cycling processes.
CONCLUSIONS
Soil moisture was the most important factor influencingRsand that it was dominantly influenced by maximum annual flood height, with areas that experienced higher flooding depth consistently maintaining significantly higher soil moisture even after the floods receded. Considering that the flood height can vary up to 4 m in várzea forests and that the difference in annual water level between flood height categories in this study was onlyca. 0.54 m, this highlights the significant effect that a small change in flood intensities could have on the release of CO2from these soils in the future.If,as predicted,flooding does increase in height and last longer,várzea forest soils may emit less CO2overall throughout the terrestrial phase.Decomposition of SOM,however, did not change significantly across the same categories, indicating the occurrence of anaerobic microbial activity.Anaerobic processes can produce other volatile C and N metabolites, which would contribute to the overall global warming potential of várzea soil gas emissions.Accordingly,future research should incorporate measurements of other greenhouse gases in order to gain a more in-depth understanding of how changes in flooding intensities might affect future soil emissions from várzea forests.
ACKNOWLEDGEMENTS
This research was supported by Department of Geography and Earth Sciences(DGES)and Institute of Biological,Environmental and Rural Sciences(IBERS)at Aberystwyth University, Wales of UK. Thanks go to them in particular for the provision of equipment and analysis facilities.We thank the Peruvian Ministry of the Environment and the Peruvian Protected Area Authority(SERNAP)for permits and collaborations. Special thanks are also extended to FundAmazonia,Amazon Eco,Operation Wallacea,the personnel of the Pacaya Samiria National Reserve,and the local Cocama guides of the Samiria River, Peru for their collaboration and support throughout our fieldwork.
杂志排行

Pedosphere的其它文章
- Notes to Authors
- Mitigating greenhouse gas emissions from croplands and pasturelands—climate-smart agriculture
- Effects of warming,wetting and nitrogen addition on substrate-induced respiration and temperature sensitivity of heterotrophic respiration in a temperate forest soil
- Long-term(42 years)effect of thinning on soil CO2 emission in a mixed broadleaved-Korean pine(Pinus koraiensis)forest in Northeast China
- Characteristics of greenhouse gas emissions from rice paddy fields in South Korea under climate change scenario RCP-8.5 using the DNDC model
- Effects of the nitrification inhibitor nitrapyrin and the plant growth regulator gibberellic acid on yield-scale nitrous oxide emission in maize fields under hot climatic conditions