犬瘟热病毒N 和H 蛋白拮抗IFN-信号通路的研究
2021-04-13龚成燕李连峰陈杰胡博赵建军
龚成燕,李连峰,陈杰,胡博,赵建军
(1.中国农业科学院特产研究所农业农村部经济动物疫病重点实验室,吉林 长春 130112;2.中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室,黑龙江 哈尔滨 150069;3.西北农林科技大学,陕西 杨凌 712100;4.黑龙江八一农垦大学,黑龙江 大庆 163319)
犬瘟热(Canine distemper, CD)是由犬瘟热病毒(Canine distemper virus, CDV)感染引起的一种热性、高度接触性传染病[1]。CDV 宿主范围广泛,可感染食肉目多个物种,包括犬科、猫科、鼬科、鬣狗科和熊科等,引起肉食动物多系统疾病,并伴有严重的免疫抑制[4],甚至对非人灵长类具有感染致死的能力[2,3]。CDV 属于副粘病毒科麻疹病毒属成员,该属成员还包括麻疹病毒(measles virus,MV)、小反刍兽疫病毒(peste despetits ruminants virus,PPRV)和被根除的牛瘟病毒(rinderpest virus, RPV)[5]。病毒基因组为单链负股RNA,长度为15 690 nt,共编码8 个蛋白。包括核衣壳蛋白(N)、血凝素(H)、基质膜蛋白(M)、磷蛋白(P)、病毒聚合酶(L)、融合蛋白(F)以及两个非结构蛋白C 和V[6]。尽管CDV 疫苗接种极大地限制了CDV的传播,但仍有免疫动物发生CDV 感染的情况[7]。因此,深入了解CDV 逃逸宿主的分子机制,将有助于通过识别药理抑制靶点来设计新的治疗策略,从而在某些情况下增强或取代疫苗。
麻疹病毒属成员具备多种策略来逃避宿主先天免疫系统。MV、PPRV、RPV 和CDVV 蛋白均可干扰酪氨酸激酶2(Tyrosine kinase 2,Tyk2)的磷酸化,并阻止信号转导与转录激活因子(signal transducers and activators of transcription,STAT)STAT 1 和STAT 2 激活,最终导致对I型干扰素(interferon,IFN)信号通路的阻断[8];Fontana 等[9]通过研究MV 弱毒疫苗株和野生毒株C、V 蛋白对干扰素信号的调控,发现V 蛋白比C 蛋白对IFN 的表达具有更强的抑制作用;通过研究PPRV野毒株和疫苗株对IFN- 的表达的影响,证实PPRV V蛋白分别通过与黑色素瘤分化相关抗原5(melanoma differentiation-associated gene-5, MDA5)和维甲酸诱导基因(Retinoic acid-inducible gene I,RIG-I)相互作用抑制IFN 通路[10]。
N 蛋白在CDV 蛋白中的含量最高,同时也是核蛋白核心的主要组成元素,具有很高的保守性,在病毒复制和转录中起主要作用,与CDV毒力密切相关[11-14]。H 蛋白是构成CDV 粒子囊膜表面的纤突,具有血凝素功能,与细胞受体结合,是病毒入侵宿主的重要调节蛋白[13,15]。H 蛋白易变异,从而引起CDV 毒力的变异[16]。表达MV Edmonston 疫苗株H 蛋白的重组MV 野毒株感染猕猴后,病毒在体内的复制水平减弱[17]。进一步研究发现,在感染表达疫苗株H 蛋白重组MV 的猕猴体内,IFN-被诱导表达,而在感染MV 野毒株的猕猴中,IFN-表达受到抑制[18]。干扰素调节因子3(interferon regulatory factor,IRF3)是诱导I 型IFN 所需的RIG-I 样受体(RIG-Ⅰ-like receptors,RLRs)通路中的关键分子。TANK 结合激酶1(TANK-binding kinase 1,TBK1)与IRF3 相互作用可促进IRF3 的磷酸化,进而活化IRF3 诱导Ⅰ型IFN 的表达[19]。PPRV N 蛋白可抑制I 型IFN 和干扰素刺激基因(ISG)的表达,其分子机制为N 蛋白结合IRF3 后抑制TBK1 与其相互作用,进而抑制IRF3 的磷酸化,阻断Ⅰ型IFN信号通路[20]。进一步证实PPRV N蛋白106-210 氨基酸区域是其抑制IFN 和ISG 的关键区域,而IRF3 的140-400 氨基酸区域是IRF3 与PPRV N 蛋白互作的关键区域[20]。与其相反,MV N 蛋白可通过诱导IRF3 的磷酸化,进而激活IRF3 诱导I 型IFN 的表达,且这种相互作用发生在N 蛋白的376-523 氨基酸区域[21]。
目前,关于CDV 疫苗株和野毒株N、H 蛋白调控干扰素表达的研究鲜有报道。因此,本研究通过构建表达CDV 疫苗株和野毒株的N、H 蛋白真核表达载体,初步探索不同毒力CDV 株N、H 蛋白对IFN- 表达的影响,可为后期研究CDV N、H 蛋白与宿主天然免疫分子的互作奠定基础。
1 材料方法
1.1 细胞、病毒和质粒
人胚胎肾细胞293T(HEK293T)、犬肾小管上皮细胞(MDCK);CDV 野毒株LN(10)1[22]、SD(14)7[23]和CDV 疫苗株CDV3[24];真核表达载体pCI 和融合表达Flag 抗原标签的H 蛋白重组质粒pCI-LN(10)1-H 和pCI-SD(14)7-H[22]均由中国农业科学院特产研究所和黑龙江八一农垦大学保存;Trans5感受态细胞购自上海生物工程有限公司;pGEM-T 载体购自Promega 公司;荧光素酶报告基因质粒pGL3-IFN- 和海参荧光内参质粒pRL-TK 由吉林农业大学曹鑫教授惠赠。
1.2 主要试剂
病毒RNA 提取试剂盒(Viral RNA Mini kit)购自QIAgen 公司;PrimeScript Ⅱ1st Strand cDNA Synthesis Kit反转录试剂盒、In-Fusion同源重组试剂盒和Premix Taq 购自TaKaRa 公司;胶回收试剂盒、中提质粒试剂盒购自Omega 公司;Cpo I 核酸内切酶、高保真DNA聚合酶、蛋白Marker 和ECL 底物发光试剂盒均购自Thermo 公司;小提质粒试剂盒购自全式金公司;Lipofectamine 3 000 转染试剂购自Invitrogen 公司;兔抗Flag 抗体购自Sigma 公司;CDV N 蛋白特异性单克隆抗体购自VMRD 公司;山羊抗兔二抗、山羊抗鼠二抗购自生工生物工程(上海)股份有限公司;Dual-Luciferase 双荧光素酶报告基因检测系统购自Promega 公司;RIPA 裂解液购于碧云天公司;Poly(I:C)购自InvivoGen 公司;Tris-MOPS-SDS Running Buffer Powder购自金斯瑞生物科技公司;转膜液购自Biosharp 公司。
1.3 CDV 感染MDCK 细胞荧光素酶表达检测
将MDCK 细胞铺一个24 孔细胞培养板,每孔约1 105个细胞,待其长至70%左右时,每孔转染pGL3-IFN-报告基因质粒(100 ng)和pRL-TK 内参质粒(10 ng),18 h后,分别将LN(10)1、SD(14)7 和CDV3 株按感染复数(MOI)0.1 感染MDCK 细胞,并设置病毒未感染组(Mock)对照,分别在感染0、8、12、24 h 后,弃掉细胞培养板的液体,加入100L/孔1 Passive Lysis Buffer裂解细胞20 mi n,转移裂解液至1.5 mL 离心管,12 000 r/min离心1 min。在1.5 mL离心管中加入20L 离心后的上清液,按照双荧光素酶检测试剂盒(名称)说明书,应用Promega Glo Max 发光检测仪测定荧光值[10]。
1.4 引物设计与合成
根据GenBank中CDV 野毒株LN(10)1(登录号:KP765764.1)、SD(14)7(登录号:KP765763.1)和CDV疫苗株CDV3(登录号:EU726268.1)基因序列,使用Oligo 软件设计扩增N 蛋白基因编码区的特异性引物[25];以疫苗株CDV 3 的基因序列为模板,根据In-Fusion 同源重组试剂盒说明书,设计一对扩增CDV3 H蛋白基因序列的特异性引物。具体引物信息见表1。引物由生物工程(上海)股份有限公司合成。

表1 扩增CDV N 蛋白和H 蛋白基因引物序列Table 1 N and H gene Primer Sequence
1.5 CDV N、H蛋白基因克隆
使用Viral RNA Mini kit 提取病毒RNA,具体操作步骤按照试剂盒说明书进行。以提取的病毒RNA 为模板,按照TaKaRa 公司的Prime ScriptTMⅡ1st Strand cDNA Synthesis kit 反转录试剂盒说明书,应用随机引物进行RNA 反转录。以反转录得到的cDNA 为模板,应用表1 特异性引物进行N 和H 基因PCR 扩增,PCR 反应体系按照Roche 公司Expand High Fidelity PCR kit 说明书进行,取全部PCR 反应液进行琼脂糖凝胶电泳检测,并回收阳性目的片段。
1.6 CDV N、H蛋白真核表达载体的构建
将LN(10)1、SD(14)7 和CDV3 株N 基因PCR回收产物与pGEM-T 载体连接,经酶切鉴定和测序,将克隆到pGEM-T 载体的目的片段与pCI 表达载体相连。重组阳性质粒pCI-LN(10)1-N、pCI-SD(14)7-N和pCI-CDV3-N 经酶切确认后送测序;将CDV3 H 基因PCR 回收产物,根据In-Fusion 试剂盒说明书,将目的基因与pCI 载体连接并转化感受态细菌,通过菌液PCR 鉴定,将阳性菌液测序确认阳性重组质粒pCICDV3-Hflag 基因序列正确。
1.7 CDV N、H蛋白真核表达载体的表达
待6 孔板中的HEK293T 细胞长至80%左右,根据Lipofectamine 3 000 转染试剂说明书,将成功构建的表达不同毒株N 蛋白和疫苗株H 蛋白的质粒转入细胞,转染量2g/孔,24 h 后,获取细胞裂解上清液,SDS-PAGE 电泳后,转印至PVDF 膜,转膜结束后用5%的脱脂奶粉封闭2 h,分别孵育抗N 蛋白单克隆抗体和兔抗Flag抗体,常温摇床2 h或4℃过夜,用PBST洗3 次后,分别孵育山羊抗小鼠二抗和山羊抗兔二抗,常温摇床1 h,用PBST洗3 次,每次15 min,按照ECL超敏显色液说明书,利用Tanon 化学发光成像仪曝光自显影检目的条带。
1.8 CDV N、H蛋白对IFN- 表达的影响
消化HEK293T 细胞,铺1 个24 孔细胞培养板,每孔约1 105个细胞,待其密度长至70% 左右,每孔转染pGL3-IFN- 报告基因质粒(100 ng)和pRL-TK内参质粒(10 ng),在此基础上,每孔分别转染500 ng不同毒株的N 或H 蛋白真核表达质粒或空载体表达质粒(pCI),转染12 h 后,参照Lipofectamine 3 000 说明书转染Poly(I:C)刺激细胞,其中未受刺激的pCI 组作为阴性对照,受刺激的pCI 组作为阳性对照,12 h后,弃掉细胞培养板的液体,加入100L/孔1 Passive Lysis Buffer 裂解细胞20 min,转移裂解液至1.5 mL离心管,12 000 r/min 离心1 min。在1.5 mL 离心管中加入20L 离心后的上清液,按照双荧光素酶检测试剂盒说明书,利用Promega Glo Max 发光检测仪测定荧光值[10]。相对荧光素酶活性值是萤火虫荧光素酶活性与海肾荧光素酶活性的比值。
1.9 统计学分析
使用GraphPad Prism 8.0 统计软件,采用单因素方差分析法对数据进行统计学分析,P<0.05 代表差异显著(*),P <0.01 代表差异极显著(**),P >0.05代表差异不显著(NS)。
2 结果
2.1 CDV 感染MDCK 细胞诱导IFN- 启动子的活性结果
转染pGL3-IFN- 报告基因质粒和pRL-TK 内参质粒,12 h 后,将LN(10)1、SD(14)7 和CDV3 分别感染MDCK 细胞,在感染后不同时间点检测荧光素酶的相对活性。结果显示疫苗株CDV3 在感染MDCK 后8 h,IFN- 启动子的活性有所上升,但随后又降低到背景水平,而野毒株LN(10)1 和SD(14)7 均未能诱导IFN- 启动子的活性,结果如图1。

图1 不同毒力CDV 感染MDCK 后不同时间IFN- 表达结果Fig.1 IFN- induction during the infection with different strains of CDV
2.2 CDV N、H 基因分子克隆结果
分别使用N 和H 基因的特异性引物扩增出目的片段,琼脂糖凝胶成像系统显示结果如图2 所示,扩增产物N 和H 大小分别为1 572 bp 和1 824 bp,与预期大小一致。将LN(10)1、SD(14)7 和CDV3 株N和H 基因克隆到pCI载体,通过酶切和测序证实pCI-N和pCI-H 重组质粒构建成功。

图2 N 和H 基因目的片段的扩增Fig.2 PCR amplification products of the CDV N and H gene
2.3 CDV N、H蛋白的表达结果
将表达CDV N 和H 基因的重组质粒pCI-N 和pCI-H 分别转染HEK293T 细胞,24 h 后获取细胞裂解液上清,利用Western blot 对其表达情况进一步验证,结果如图3 所示,N 蛋白大小约为56 kDa,融合表达Flag 抗原标签的H 蛋白大小约为110 kDa,表明各真核表达载体在HEK293T 细胞中正确表达,其中pCI-LN(10)1-H 及pCI-SD(14)7-H 真核表达载体之前已由本实验室构建完成并可正常表达[22]。

图3 N 和H 重组质粒的表达Fig.3 The expression of N and H recombinant plasmids
2.4 CDV N、H蛋白对-干扰素表达的影响
在HEK293T 细胞中过表达不同毒株CDV N 蛋白,12 h 后用Poly(I:C)刺激细胞,利用双荧光素酶报告系统检测试剂盒检测IFN- 启动子的活性,结果如图4A 所示,阳性对照组IFN- 启动子的相对活性为20.89,极显著高于阴性对照组(P <0.01),表明Poly(I:C)成功激活IFN- 信号通路;而这种激活作用在过表达不同毒力的N 蛋白时极显著受到抑制(P <0.01),pCI-LN(10)1-N、pCI-SD(14)7-N 和pCI-CDV3-N 实验组IFN- 启动子的相对活性分别为3.98、4.10 和8.32。同样方法在HEK293T 细胞中过表达不同毒株CDV H 蛋白,结果如图4B 所示,Poly(I:C)成功激活IFN信号通路,阳性对照组IFN- 启动子的相对活性为26.22,pCI-LN(10)1-H、pCI-SD(14)7-H 和pCI-CDV3-H实验组的IFN- 启动子的相对活性分别为5.28、5.70和6.79,极显著低于阳性对照组,说明不同毒力H 蛋白的过表达极显著抑制IFN- 通路的激活作用。
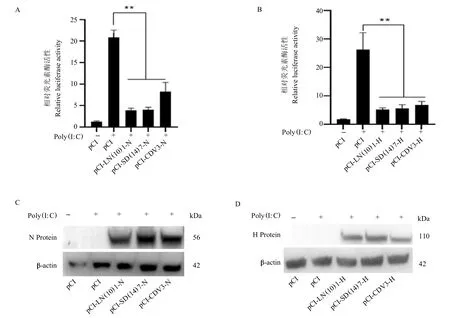
图4 不同CDV 毒株N、H 蛋白抑制IFN- 启动子的激活Fig.4 N and H protein of different strains suppresses IFN- promoter activation
3 讨论
在病原微生物入侵以后,宿主通过多种细胞模式识别受体(Pattern Recognition Receptors, PRRs)识别病原代谢过程中产生的保守组分病原相关分子模式(Pathogen-associated Molecular Pattern,PAMPs),经由不同的接头分子,活化多条信号通路,诱导炎性因子及IFN 的产生,以对抗病原微生物的感染,RLRs 是经典的PRRs[13]。然而,大多数麻疹病毒属成员进化出多种对抗宿主免疫系统的策略:一是病毒蛋白通过干扰RLRs等介导的IFN 上游信号通路的激活来抑制IFN 的表达,大部分RPV、MV、CDV 和PPRV 都能拮抗I型IFN的产生[26-28];二是病毒蛋白通过干扰IFN 下游信号通路的激活来抑制众多抗病毒蛋白的产生[27-29],如MVV蛋白通过与STAT1/2 结合,阻止STAT 蛋白向细胞核迁移,抑制IFN 诱导抗病毒因子的表达[8];同样,CDV V蛋白的两个结构域(N 端和C 端)选择性地抑制了STAT1 和STAT2 入核[30],进而阻断了IFN 诱导宿主产生抗病毒蛋白的信号通路。
为进一步在分子水平理解CDV 诱导宿主的免疫抑制机制,本研究通过不同病毒株感染MDCK 细胞和表达不同CDV 毒株H 和N 蛋白,研究其对IFN- 信号通路的调控作用。CDV 野毒株SD(14)7 和LN(10)1分离自2014 年和2010 年CD发病死亡狐狸。与LN(10)1株相比,SD(14)7 株H 蛋白多处氨基酸发生变异,引起CDV 对水貂、狐狸的毒力和免疫抑制增强[22]。CDV3 疫苗株为我国毛皮动物养殖场使用的CDV 疫苗株, 能较好地预防CD 的暴发流行。MDCK 细胞为CDV 易感宿主细胞,在病原微生物入侵时,可迅速产生包括IFN 在内的抗病毒因子[31],因此,本研究选择MDCK 细胞作为CDV 体外感染靶细胞。研究结果证实,疫苗株和野毒株CDV 感染MDCK 细胞后,只有疫苗株能引起IFN- 启动子的短暂激活,而野毒株在感染MDCK 细胞24 h 内,IFN- 启动子未激活,说明CDV 在感染MDCK 细胞后,可能是抑制了IFN- 的表达,与后续实验结果符合。Takayama 等[32]研究发现MV N 蛋白既不抑制STAT 和Jak 磷酸化也不引起STAT 降解,但N 蛋白的核移位干扰了STATs 入核,从而阻断IFN 的信号通路。Takayama 等[32]进一步研究发现在IFN-和IFN-刺激下,CDV N 蛋白同样具有抑制IFN-和IFN-下游信号通路激活,即抑制IFN诱导抗病毒蛋白产生。本研究结果显示,CDV N 和H蛋白均可下调Poly(I:C)诱导IFN- 启动子的活性,说明CDV N 和H 蛋白可通过干扰IFN- 信号通路来抑制IFN- 表达。Fontana 等[9]通过研究MV 疫苗株和野毒株C 和V 蛋白对IFN 调控的影响发现,V 蛋白比C 蛋白对IFN 信号转导具有更强的抑制作用,且这种抑制作用与MV 的毒力无关。本试验结果也显示CDV N 或H 蛋白下调IFN- 的表达与CDV 毒力无关,说明不同毒力的CDV N 或H 蛋白均可入侵宿主并可能与IFN- 信号通路中多个蛋白互作,但具体分子通路还有待进一步研究。
本实验通过研究疫苗株和野毒株N 和H 蛋白对IFN- 表达的影响,发现N 和H 蛋白抑制宿主天然免疫与CDV 毒力无关。未来将构建表达N 和H 蛋白截短体,进一步研究病毒蛋白抑制IFN 表达的关键氨基酸区域,有助于进一步理解CDV 基于分子水平的致病机制,为开发新型抗CDV 药物提供一定的理论依据。
