裂壶藻营养价值及其在动物生产中的应用进展
2021-04-08张新明胡晓文
张新明,赵 璐,胡晓文
(1.日照职业技术学院 海洋工程学院,山东 日照 276826; 2.日照市海洋生物工程技术研究中心,山东 日照 276826; 3.中国海洋大学 海洋生命学院,山东 青岛 266003)
裂壶藻(Schizochytriumlimacinum)是一种海洋微藻,在分类上属于真核生物域,囊泡藻界,异鞭藻类,双环门。它易于规模化生产,适合大范围推广和使用[1]。裂壶藻为较小的球形,适于被幼仔水生动物捕食,具有较高的饵料效率和开发利用价值[2]。裂壶藻含有丰富的高度不饱和脂肪酸二十二碳六烯酸(Docosahexaenoic acid,DHA)和蛋白质、矿物质等营养物质,其营养价值与碳源、氮源、pH值、外源性添加物等环境条件有关[3-7],同时裂壶藻含有多糖、活性肽、角鲨烯等物质[2,8-10],在动物生产中具有非常好的应用前景。裂壶藻作为饲料添加剂直接在饲料中添加,或替代部分蛋白源和油脂在鱼类、虾类等水生动物以及蛋鸡中广泛应用,具有提高存活率,提高产品DHA含量,增强体色,促进脂肪代谢,提高非特异性免疫能力等功效[11-13],其效果与添加量、添加方式、裂壶藻来源等因素有关。对裂壶藻的营养效用、活性物质、对环境的响应机制和在动物生产中的应用情况进行综述,以期为裂壶藻开发利用提供参考。
1 裂壶藻的营养价值
1.1 裂壶藻的基本营养
裂壶藻营养丰富,不仅含有蛋白质、脂肪、糖类、灰分等常规营养物质,而且含有丰富的必需氨基酸和DHA等高不饱和脂肪酸,是一种具有重要应用价值的微藻。
裂壶藻的营养成分因种类不同而有所差异,研究人员对不同种类的裂壶藻营养物含量的测量值差异比较大(表1)。
从表1可以看出,裂壶藻蛋白质含量为9.35%~42.51%,脂肪含量为18.98%~56%,糖类含量为5.27%~29.26%,灰分含量为4.85%~12%。裂壶藻还含有丰富的Mg、Ca、Na、Fe等元素,含有少量的Cu、Mn、K、Zn、Al,含有微量的Pb[16]。
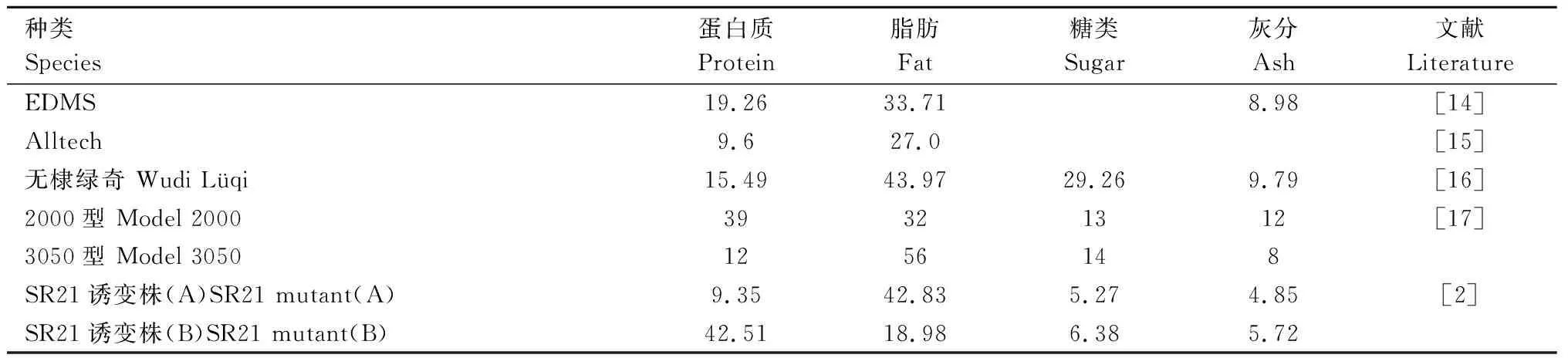
表1 裂壶藻基本营养成分Tab.1 Basic nutrients of Schizochytrium limacinum %
裂壶藻主要营养成分为蛋白质和脂类,同时含有少量的糖类和灰分。裂壶藻蛋白质和脂肪含量因培养条件不同产生很大的差异,而糖类和灰分的含量相对稳定。以豆粕水解物为氮源时,主要成分是脂类(含量为42.83%),其次是蛋白质(含量为9.35%);以酵母提取物为氮源时,主要成分是蛋白质(含量为42.51%),其次是脂类(含量为18.98%)。2种条件下总糖和灰分含量变化不大(含量分别为5.27%~6.38%、4.85%~5.72%)[2]。
裂壶藻含有16种氨基酸(10种必需氨基酸和6种非必需氨基酸),谷氨酸、天冬氨酸、脯氨酸、亮氨酸含量较高,组氨酸、蛋氨酸、酪氨酸、异亮氨酸含量最低。不同的培养基对氨基酸的含量也会产生影响。分别以豆粕水解物和酵母提取物为氮源时,氨基酸组成种类相同,总氨基酸含量分别为7.38%和39.7%,必需氨基酸均占总氨基酸总量的40%左右。从种类上来看,2种氮源条件下分别以谷氨酸、天冬氨酸和谷氨酸、精氨酸含量较高,精氨酸、组氨酸和蛋氨酸、组氨酸含量最低[2,18]。
裂壶藻含有丰富的不饱和脂肪酸,尤其是DHA含量非常高。裂壶藻脂肪酸含量占干质量的18.3%~25.2%,总脂肪酸主要由DHA(C22∶6n-3)、二十二碳五烯酸(Docosapentaenoic acid,DPA)(C22∶5n-6)和棕榈酸(C16∶0) 组成,其中多不饱和脂肪酸(Polyunsaturated fatty acid,PUFA)DHA 含量占总脂肪酸含量的39.62%~49.84%,DPA含量约为12.5%,棕榈酸约为41.8%~46.04%,二十碳五烯酸(Eicosapentaenoic acid,EPA)和n-6PUFA含量较少,EPA含量占总脂肪酸含量的1.25%[16,19-22]。裂壶藻含有17种脂肪酸,其中饱和脂肪酸(Saturated fatty acid,SFA)9种,PUFA 8种,脂肪酸含量最丰富的主要为C16∶0和C22∶6n-3(DHA)。脂肪酸含量与培养条件有关,分别以豆粕水解物和酵母提取物为氮源时,总脂肪酸中DHA含量分别为36.84%和28.63%[2]。
1.2 裂壶藻油脂的主要成分
裂壶藻油脂主要包括甘油酯、磷脂和不皂化物3类物质。甘油酯以甘油三酯为主,磷脂以磷脂酰胆碱为主,不皂化物包括烷烃、二丁基羟基甲苯、角鲨烯、胆固醇、麦角固醇、豆甾醇、β-谷甾醇、环阿屯醇,角鲨烯和胆固醇的含量分别为19.99 mg/g和40.93 mg/g。裂壶藻总油脂由73.60%~87.86%中性脂类和11.32%~26.40%极性脂类组成,极性脂中DHA质量分数(52.01%~76.10%)显著高于中性脂(34.80%~35.51%)。裂壶藻合成的色素主要有虾青素、β-胡萝卜素、角黄素等,β-胡萝卜素含量为34.91 μg/g[8-9,21]。
1.3 裂壶藻的生物活性物质
裂壶藻除含有常规营养物外,还含有多糖、活性肽等具有免疫活性或抗菌活性的物质。宋泽等[16]研究发现,裂壶藻可溶性多糖提取率为2.13%,其多糖含量为49.81%,多糖的单糖主要以半乳糖为主(56.84%),并含有少量的甘露糖、鼠李糖,另外还含有少量的葡萄糖、木糖;硫酸基含量为4.76%,多糖的硫酸基团主要位于半乳糖残基的C6位。曹欢等[23]从裂壶藻中获得2种纯品多糖,相对分子质量分别为39.8 ku和103.8 ku,主要为硫酸半乳聚糖,其硫酸酯基主要在半乳糖残基的C6位。汪少芸等[24]从裂壶藻中分离得到酸性多糖、弱碱性多糖和碱性多糖等3个主要组分,结果表明,弱碱性多糖具有较高的抗氧化活性;酸性多糖、弱碱性多糖和碱性多糖对苹果轮纹病菌、瓜果腐霉病菌、葡萄灰霉病菌和甜瓜枯萎病菌有一定强度的抗菌性,其中对葡萄灰霉病菌抑菌效果较为明显。梁双振[10]从裂壶藻藻粕酶解提取物得到活性物质为分子质量不足3 000 u的蛋白肽及酶解肽混合物。蛋白肽对小鼠体外炎症具有一定的抵抗作用。酶解肽对金黄色葡萄球菌抑菌性最强,大肠杆菌次之,沙门菌最弱。
裂壶藻DHA具有较强的免疫增强活性,在一定条件下,可显著提升吞噬细胞的增殖能力及吞噬活性、细胞酸性磷酸酶(Acid phosphatase,AKP)和溶菌酶的分泌及活性[25]。裂壶藻藻粕胰蛋白酶酶解生物活性肽添加质量浓度为0.1~0.001 g/L时,能明显促进成纤维细胞的增殖。活性肽能够维持小鼠肠道黏膜相对较好的吸收代谢能力,具有抗炎的生物活性,可以治疗肠道炎症,功效优于诺氟沙星[26]。
2 裂壶藻DHA对环境的响应机制
无机盐类对裂壶藻生长和发酵产物的积累具有非常重要的影响,在培养基中适量添加无机盐能显著提高裂壶藻中DHA产量。NaCl与细胞内相关酶系和酶活性的改变有关,在培养过程中,裂壶藻油脂产量、DHA产量随着NaCl质量浓度的增大而增加,质量浓度为3 g/L时,DHA产量最大。镁是裂壶藻代谢关键酶的激活剂。随着Mg2+质量浓度的增加,油脂产量也增加,裂壶藻的培养过程中,MgSO4质量浓度为6 g/L时,油脂产量和DHA产量最大。磷在细胞的生理代谢过程中发挥着重要作用,是机体重要化合物(如核苷酸、磷脂、三磷酸腺苷酶等)的组成成分,磷酸盐质量浓度会影响脂肪酸代谢中某一关键酶的活性,裂壶藻培养过程中油脂产量和DHA产量最大时,KH2PO4最佳质量浓度为4.5 g/L[27]。
碳源、氮源种类和质量浓度同样会影响裂壶藻的生长和脂肪物质的积累。3%~6%葡萄糖作为碳源能明显促进裂壶藻生长和DHA积累;氮源质量浓度提高能促进机体生长,对DHA含量无显著影响[28]。碳源、氮源质量浓度对裂壶藻生长和脂肪酸组成影响不同,分别以葡萄糖和豆粕水解物为碳源和氮源,质量浓度高时细胞生物量较高,而质量浓度较低时,细胞总脂含量以及总脂中的DHA含量较高[3]。裂壶藻产油量和DHA产量均随着葡萄糖质量浓度增加(20~80 g/L)而升高,质量浓度再增加会发生抑制作用。较高的初始氮源质量浓度对机体生长有利,但对油脂合成和DHA的积累会产生负面影响[4]。甘油作为碳源时,虽然利用率高,但是其油脂产量却比葡萄糖作为碳源时低[8]。裂壶藻高产油脂的最优碳氮比约为5∶1,而高产DHA的最佳碳氮比约为2.5∶1,高油脂产量与最优DHA产量并不同步,高碳氮比有利于脂质的积累,较低碳氮比有利于DHA的合成[21]。而康晶等[5]研究发现,0.4 g/L氮源最适合裂壶藻油脂和DHA积累,最佳C∶N为80∶1。
温度、溶解氧和pH值同样会影响裂壶藻生长和产油量。机体生物量和产油量随着温度的升高(18~28 ℃)而增加,而DHA产量超过24 ℃会降低[4]。低溶氧不利于机体生长但有利于DHA合成,氮源限制时溶氧水平为5%条件下利于DHA发酵,DHA产量达11.5 g/L,占总脂肪酸含量的百分比为44.41%[5]。培养液pH值对裂壶藻生长及其油脂产量、DHA产量都有很大的影响,在pH值3~9,生物量随pH值升高而降低[28];pH值5~7有利于机体生长;pH值3~4有利于油脂和DHA的积累。发酵培养前4 d pH值为3,后2 d pH值为5条件下,其总脂和DHA的产量较大,分别为9.92 g/L和4.14 g/L[6]。
添加外源添加剂能够提高裂壶藻合成不饱和脂肪酸的含量。添加苹果酸和柠檬酸,能提高DHA比例、油脂产量以及生物量,并促进碳源的利用,促进产油[8]。同时添加0.3 mg/L生物素、1 g/L苹果酸、0.4 mg/L洛伐他汀时DHA的产量(11.55 g/L)比对照提高71.87%[7]。添加150 mg/L乙醇胺、10 mg/L萘氧乙酸和1 mg/L水杨酸,DHA产量达到6.18 g/L,比对照提高12.77%[29]。添加生物素可促进油脂积累,添加浅蓝菌素有利于DHA及不饱和脂肪酸含量的提高。添加0.3 mg/L的生物素时,产油量增加27.9%,DHA占细胞干质量的比例增加13%;每克干细胞中含角鲨烯0.57 mg,相比对照提高了30%。添加浅蓝菌素时,会抑制饱和脂的合成,有利于不饱和脂的合成,DHA含量提高5%,DHA占细胞干质量的比例可达12%[30]。糠基腺嘌呤(KT)、6-苄基腺嘌呤(BA)、赤霉素(GA)、萘乙酸(NAA)、吲哚乙酸(IAA)、吲哚丁酸(IBA)6种激素能不同程度提高裂壶藻生长速度和DHA含量,但质量浓度过高会产生抑制作用。6种激素适宜质量浓度分别是BA 3 mg/L、KT 10 mg/L、GA 1 mg/L、IBA 3 mg/L、NAA 6 mg/L、IAA 3 mg/L;BA添加量为3 mg/L时,裂壶藻生物量和DHA含量分别比对照组提高27.1%和30.3%[31]。
二水平主斜井装备两台GDS-100型钢丝绳牵引带式输送机,设计运输能力400吨/小时,带宽1m,全数字直流调速,最大运行速度1.8 m/s,运输长度1490m,井筒平均倾角17°,电机功率为2×400 kW。主要担负井下戊一岩石带式输送机系统原煤运输任务。
维生素B1(VB1)、维生素B3(VB3)、维生素B6(VB6)、维生素B12(VB12)和生物素5种B族维生素均可提高DHA产量,最适质量浓度分别为0.100 mg/L、0.005 mg/L、0.005 mg/L、0.300 mg/L和0.050 mg/L,VB12对DHA产量提高效果最好,VB6和VB3次之,生物素和VB1最差。5种维生素以最适质量浓度添加到培养基中,DHA产量提高了53.8%[32]。VB1、VB12、生物素、叶酸和硫辛酸5种维生素能够促进裂壶藻机体生长,提高DHA 的比例,但是对于油脂产量和生物量的促进效果不明显[8]。
3 裂壶藻在生产中的应用
3.1 在水产动物生产中的应用
3.1.1 在促进水产养殖动物生长和存活方面的应用 裂壶藻作为添加剂添加到饲料中或者替代部分蛋白源能够促进动物存活或生长。在鲍苗配合饲料中添加30%裂壶藻粕对鲍苗成活率影响不明显,但能提高鲍苗生长速度,壳长和体质量比对照组分别增加了3.4%和16%[18]。随着饲料中裂壶藻添加量(1.25%~2.00%)的升高,刺参特定生长率显著提升[33]。
以裂壶藻粕作为配合饲料的部分营养源,可以发挥互补作用,提高饲料质量和养殖效果。以裂壶藻粕(25%)代替混合饲料中部分鱼浆,赤点石斑鱼(Epinephelusakaara)体长提高1.2%,体质量增加9.6%[34]。微藻中的DHA等高不饱和脂肪酸可以被牙鲆稚鱼充分消化和吸收,满足仔稚鱼的生长和发育,以微藻替代鱼油在牙鲆稚鱼的培育中是可行的,在饲料中添加裂壶藻可以显著提高牙鲆鱼的成活率[15]。添加裂壶藻粉能显著提高星斑川鲽增重率、特定生长率和脏体比,但对存活率、饲料系数、摄食率、肥满度、肝体比、蛋白质效率没有显著影响[35]。
裂壶藻粉在饲料中的添加量为1.0%~1.5%时,显著增加了斑点叉尾鮰(Ictaluruspunctatus)的体质量,添加0.5%~2.0%的裂壶藻其饲料效率显著提高[36]。饲料中添加8%螺旋藻、2%裂壶藻和8%大型溞粉可提高红草金鱼(Carassiusauratusred var)的生长性能和饲料利用率,其蛋白质效率、特定生长率、肥满度与增重率显著提高,饲料系数显著降低[37]。裂壶藻可显著影响津新鲤(Cyprinuscarpiovar.Jian)的生长性能,在0.80%裂壶藻水平组中,增重率和特定生长率均显著提高,肥满度则在1.20%水平组中达到最大值[11]。在饲料中添加1%裂壶藻能有效提高非洲王子鱼(Labidochromiscaeruleus)的增重率、特定生长率及饲料效率[38]。在基础饲料中添加6.0%盐藻、1.5%裂壶藻、6.0%玉米蛋白粉可以显著提高红草金鱼体蛋白质效率、特定生长率、肥满度和增重率,降低饵料系数[39]。投喂经过裂壶藻、维生素C(VC) 及海水鱼多维强化的生物饵料,可以显著降低大菱鲆仔稚鱼的白化率和畸形率,提高仔鱼的存活率[40]。乳酸链球菌和裂壶藻联合使用显著提高了驼背鲈(Cromileptesaltivelis)终体质量、增重率和生长速率,提高了对哈维氏弧菌的抵抗力[41]。饲料中添加3%裂壶藻,能够显著提高斑马鱼感染创伤弧菌后的存活率,相对保护率为42.8%;显著提高斑马鱼肾脏中溶菌酶的信使RNA(mRNA)水平,显著降低肿瘤坏死因子的mRNA水平,从而抵抗细菌的感染,提高存活率[19]。
裂壶藻粕中赖氨酸、蛋氨酸和精氨酸分别占蛋白质的4.8%、2.5%和4.3%,非常接近南美白对虾(Penaeusvannamei)的氨基酸需求,可以作为南美白对虾配合饲料新的蛋白源[42]。添加0.5%的裂壶藻发酵粉,能有效提高南美白对虾个体质量,降低饲料系数,但对对虾增重率、成活率和饲料系数影响并不显著,过量增加并不会进一步促进虾的生长,甚至会影响虾的生长[12]。杨帆[43]的研究结果表明,饲料中添加3%的破壁裂壶藻粉,可显著提高凡纳滨对虾(Litopenaeusvannamei)的生长性能,显著提高对虾末均质量、存活率和增重率,显著降低饲料系数。在基础饲料中添加10%的裂壶藻,可提高中国对虾(Fenneropenaeuschinensis)体长、体质量和存活率,影响并不显著,但与维生素E(VE)联合使用对中国对虾影响显著[44-45]。饲料中添加4.5%破壁裂壶藻粉时,对虾生长性能反而下降。饲料中添加未破壁裂壶藻粉时,对虾生长性能没有显著变化。可能由于脂质存在于由细胞壁包裹的体细胞内,其含有的DHA不能完全被吸收利用[43]。饲料中添加200 mg/kg VE和1%裂壶藻均能促进斑节对虾(Penaeusmonodon)精荚的再生,两者有协同作用,还能显著增加精子团的数量,会影响丝裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK)信号通路的相关基因在斑节对虾精荚组织的相对表达量,激活细胞膜上的信号传递因子,并将促生长信号传递到精子核内,使核转录因子c-Myc等活性加强,促进细胞增殖分化,从而促进精荚成熟[46]。
3.1.2 在提高水产养殖动物免疫机能方面的应用 添加裂壶藻粉能显著提高对虾脂肪代谢能力和非特异性免疫机能。饲料中添加0.5%~2.0%裂壶藻粉,凡纳滨对虾肌肉和肝胰脏中高不饱和脂肪酸(Highly unsaturated fatty acids,HUFA)含量、血清总甘油三酯(TG)含量显著升高,总胆固醇(TC)含量保持稳定,1.5%组高/低密度脂蛋白(HDL/LDL)显著高于其他各组,高质量浓度组超氧化物歧化酶(SOD)显著提升。饲料中添加裂壶藻促进了凡纳滨对虾肌肉中n-3HUFA的沉积,同时也提高了虾体脂质过氧化压力[14]。饲料中添加3%的破壁裂壶藻粉能增强脂肪分解代谢能力,显著提高对虾血清TG以及HDL/LDL,说明裂壶藻能促进血液向肝脏输送胆固醇,避免胆固醇在血管中的堆积,从而提高了胆固醇的利用效率。添加裂壶藻粉后,凡纳滨对虾肝胰腺总抗氧化能力显著增加,肝胰腺丙二醛(MDA)含量显著降低。肝胰腺SOD和谷胱甘肽过氧化物酶(GSH-PX)基因的表达水平显著提高,肠道胰蛋白酶、糜蛋白酶、胰脂酶基因的表达水平显著提高[43]。
VE对不饱和脂肪酸有保护作用,可以防止裂壶藻中的DHA氧化,进而减少体内过氧化物的产生,并且促进机体免疫功能的发挥,对抗刺激及应激的能力增强。添加裂壶藻对中国对虾血清、肝脏、肌肉和鳃中总蛋白质含量起到促进作用,VE与裂壶藻联用溶菌酶活性升高,血清、肝胰腺和鳃SOD活性显著增强,MDA含量显著降低。VE和裂壶藻联合使用对免疫基因的调控能力更加显著,可显著降低血液、肝胰腺和肌肉中Toll样受体(TLR)、核因子κB(NF-κB)基因的表达水平,提高机体抗氧化能力,从而增强中国对虾免疫力[44-45]。投喂不同质量浓度虾青素和裂壶藻后,中国对虾肝胰腺、肌肉、血清、鳃中的总蛋白质、SOD和过氧化氢酶(CAT)活性均有所上升,而对MDA均有不同程度的抑制作用,说明虾青素和裂壶藻联合作用可以增强中国对虾的免疫能力[47]。
裂壶藻具有较好的降脂和提高机体抗氧化与非特异性免疫能力的作用,显著提高红草金鱼血清和各组织中SOD、过氧化氢酶活性与一氧化氮含量,降低MDA含量及血清中TC含量[37]。投喂经裂壶藻强化的卤虫可以提高半滑舌蹋稚鱼体质量、碱性磷酸酶(ACP)、碱性蛋白酶活性和甲状腺素、三碘甲状腺原氨酸水平,说明裂壶藻营养强化增加了卤虫体内脂肪酸尤其是必需脂肪酸的含量,所含有的营养物质更能满足稚鱼的营养需求[48]。裂壶藻粉可在一定程度上提高星斑川鲽营养价值,提高机体的脂肪代谢能力,改善鱼体的抗氧化能力和非特异性免疫能力。肝脂酶、脂蛋白脂酶、总脂酶活性都显著升高,提升了鱼体血清总抗氧化能力、溶菌酶含量、补体C4含量,迟缓爱德华氏菌攻毒存活率显著提高[35]。饲料中添加0.80%~1.20%的裂壶藻,可显著提高津新鲤免疫能力,提高CAT、SOD、溶菌酶、一氧化氮合酶活性和谷胱甘肽的含量,降低MDA含量和谷草转氨酶(GOT)/谷丙转氨酶 (GPT)比值。随着裂壶藻质量浓度的增加,津新鲤血细胞呼吸爆发活性显著提高,感染嗜水气单胞菌后的累计死亡率显著降低[11]。饲料中添加1%裂壶藻,非洲王子鱼血清中GPT、GOT活性显著降低,肠道中AKP、ACP活性显著提高[38]。饲料中添加1%的裂壶藻,对血艳红慈鲷(Copadichromistrimaculatus)肝脏和血液中的AKP、ACP、GPT、GOT的活性均无显著影响[49]。基础饲料中添加6.0%盐藻、1.5%裂壶藻、6.0%玉米蛋白粉,可以显著降低红草金鱼TG与TC含量,提高组织中SOD及CAT活性,降低MDA含量[39]。乳酸链球菌和裂壶藻联合使用显著提高了驼背鲈巨噬细胞的呼吸爆发活性,血清SOD、碱性磷酸酶、溶菌酶活性[41]。
饲料中添加裂壶藻后,刺参体腔液溶菌酶、SOD、碱性磷酸酶、肠道淀粉酶、肠道蛋白酶活性及吞噬细胞吞噬活性和呼吸爆发活性显著高于对照组,随着添加量的增加,这些酶的活性都呈现上升趋势[33]。
3.1.3 在提高水产养殖动物产品品质方面的应用 饲料中添加裂壶藻(0.5%~2.0%),凡纳滨对虾肌肉中粗脂肪和粗蛋白含量均未显著改变,但灰分含量随着添加量的增加呈现上升趋势[14]。用裂壶藻强化的轮虫投喂凡纳滨对虾幼体后,提高了幼体的脂肪酸含量,显著提高幼体的DHA、ω-3HUFA、DHA/EPA 含量和变态率[50]。裂壶藻发酵粉对南美白对虾肌肉灰分和粗脂肪含量无显著影响,但能显著影响肌肉中蛋白质的含量和粗脂肪中DHA的含量,1.0%组蛋白质含量最高,0.5%组粗脂肪中DHA的含量最高。南美白对虾能够通过饲料营养通道来富集 DHA,说明裂壶藻发酵产物应用于南美白对虾饲料中能改善肌肉的品质[12]。裂壶藻粉对星斑川鲽全鱼和肌肉中粗蛋白、粗脂肪、粗灰分没有显著影响,肌肉和肝脏中的DHA含量和DHA/EPA值显著提高,EPA含量降低[35]。裂壶藻具有较好的增红提亮效果,红草金鱼体表亮度与红度值明显增加[37]。饲料中添加1%的裂壶藻,可以显著提高非洲王子鱼体表或血艳红慈鲷鳍条中总类胡萝卜素含量和体表红度值(b值),对体色有着明显的改善作用[38,49]。基础饲料中添加6.0%盐藻、1.5%裂壶藻、6.0%玉米蛋白粉,可以显著提高红草金鱼皮肤、鳞片和鳍条中类胡萝卜素含量[39]。
3.1.4 在水产养殖饵料生物上的应用 用裂壶藻作为营养强化剂能促进轮虫和卤虫的生长发育,提高体内DHA等多不饱和脂肪酸的含量,显著提高轮虫和卤虫的营养价值。用50 mg/L和80 mg/L裂壶藻强化的轮虫活力和摄食状况明显加强,轮虫增长速率提高5倍以上,怀卵率提高80%以上,DHA含量达到7.85~8.14 mg/g[51]。用裂壶藻粉对褶皱臂尾轮虫进行营养强化后,轮虫密度和怀卵率较油脂组显著增加,体内的EPA、DHA、PUFA、HUFA含量及DHA/EPA比值明显增加[50,52]。卤虫无节幼体经裂壶藻营养强化24 h后,营养价值显著提高,EPA+DHA的总量达到无节幼体干质量的1.3%[17]。
3.2 在畜禽和其他动物生产中的应用
3.2.1 在蛋鸡生产中的应用 饲粮中添加不同含量(0.5%~3.0%)裂壶藻粉对海兰褐蛋鸡产蛋率、平均蛋质量、料蛋比、蛋形指数、蛋壳厚度、蛋壳强度、哈氏单位、蛋黄水分和冻干蛋黄质量影响不显著。添加0.5%的裂壶藻粉,脂蛋白酯酶和肝酯酶活性显著提高。添加1.0%~1.5%裂壶藻粉,其血清TG、TC和低密度脂蛋白胆固醇含量显著低于对照组;但添加量增加到3.0%时,略有升高趋势。添加裂壶藻粉可以线性增加鸡蛋中ω-3 PUFA和DHA含量,降低ω-6/ω-3,对蛋黄α-亚麻酸含量无影响。添加第15天时,蛋黄中DHA和EPA含量最高,28 d后达到稳定。当裂壶藻粉添加量大于2%时,蛋黄中DHA含量不再增加[13,22,53]。
日粮中分别加入1%~3% DHA含量为15%的裂壶藻粉,对18~20周龄杏花鸡鸡蛋油脂总量没有明显影响,花生四希酸(Arachidic acid,ARA)在油脂中的含量、蛋黄胆固醇含量有所下降,其他脂肪酸含量无显著变化。鸡蛋的DHA含量随添加量增加和饲养时间延长呈上升趋势,DHA含量从1.5%提高至5.25%;对料蛋比、日采食量及日产蛋量、蛋壳厚度、蛋黄相对质量、平均蛋质量无明显影响,但当添加量增大至3%时,产蛋率与对照组相比下降22.1%[54]。
饲粮中添加胆碱与裂壶藻油对26周龄京红蛋鸡鸡蛋卵黄中粗脂肪、TC及TG含量均无显著影响。胆碱和裂壶藻有交互作用,1 000 mg/kg胆碱+0.5%裂壶藻粉组鸡蛋卵黄总磷脂含量和DHA含量显著提高,n-6PUFA含量显著降低,而对蛋鸡生产性能和蛋品质无显著影响[55]。
3.2.2 在奶牛生产中的应用 日粮中添加裂壶藻粉对奶牛产奶量无影响,对牛奶中DHA含量的富集具有极显著的影响,随着裂壶藻粉用量的增加,牛奶中DHA含量和不饱和脂肪酸水平显著提升,适宜添加量为150~250 g/(头·d),牛奶中DHA含量达到0.1~0.2 g/L。添加量为250 g时,牛奶中DHA的相对含量达到52.82%,与母乳含量接近。超过250 g后,牛奶中DHA含量的转化率降低,牛奶的乳脂含量显著降低[56]。
3.2.3 在小鼠中的应用 通过高脂饮食建立小鼠肥胖模型,裂壶藻DHA油可有效预防高脂饮食诱导的小鼠体质量增加和附睾脂肪、肠系膜脂肪、肾周脂肪和腹股沟脂肪等腹部脂肪积累,TC和TG含量显著降低,脂肪细胞体积和肝组织异位脂肪堆积明显减少,明显缓解了肝组织内脂滴数量,脂肪组织中激素敏感脂酶基因的mRNA表达水平显著提高,能够抑制高脂饮食诱导的肝组织脂肪变性[57-58]。饮食中添加6%的裂壶藻油,小鼠的体质量降低至对照组的86%,血糖降低至对照组的85%,EPA和DHA在小鼠的附睾脂肪组织中的沉积浓度升高,小鼠肝脏中过氧化物酶体增殖物激活的受体γ编码基因的表达水平显著提高,抗炎细胞因子TGF-β1的血浆水平显著升高[59]。
昆明孕鼠从受孕到仔鼠断乳连续灌胃裂壶藻提取物,能增加仔鼠的体质量、身长,对孕鼠分娩总数、活胎数及胎仔总数无明显影响,对神经发育有明显的促进作用,仔鼠的平面翻正时间缩短,前肢握力时间延长,但对仔鼠的悬崖回避时间、空中翻正阳性率、耳廓分离时间、上门牙萌出时间、下门牙萌出时间、开眼时间、阴道开放时间和睾丸下降时间等指标没有显著影响[60]。
3.2.4 在其他动物中的应用 用含有10%裂壶藻藻渣的日粮饲喂8日龄黑水虻幼虫,其体质量及体长、粗蛋白与粗脂肪含量没有显著变化。黑水虻幼虫可以富集裂壶藻藻渣中的n-3PUFA,且随着裂壶藻添加量的增加,幼虫体内n-3PUFA含量显著提升[61]。
4 展望
裂壶藻含有丰富的营养价值,尤其含有动物生长所需的DHA等必需脂肪酸,在饲料中添加一定量的裂壶藻能够提高动物的生长性能和存活率,提高产品的营养价值,增强体色,增加机体对DHA等物质的积累,并且能在一定程度上提高机体对脂肪的代谢能力和非特异性免疫能力,降低血清TC和TG含量,降低MDA含量,提高酸性磷酸酶、碱性磷酸酶、SOD活性。因此,裂壶藻在动物生产中有非常好的应用前景。
裂壶藻属于高脂低蛋白的营养物质,不宜作为仔稚鱼的开口饵料单独使用。研究发现,作为出膜5 d后多鳞白甲鱼的饵料,在生长方面,裂壶藻效果不及商品饲料,但对存活率的提高效果较为明显[62]。用裂壶藻替代50%的鱼油对海鲈(Lateolabraxjaponicus)生长没有明显影响,但完全替代则会抑制生长,导致增重率下降,裂壶藻替代鱼油所引起的EPA和ARA含量降低也是影响鱼体生长的重要因素[63]。豆粕、大豆浓缩蛋白和裂壶藻粉混合物可以替代约40%的鱼粉等海洋来源蛋白质,并且以裂壶藻粉作为饲料主要脂质来源,不会对鞍带石斑鱼(Epinepheluslanceolatus)的生长产生显著影响,但是替代过多就会产生不利影响[64]。
今后对裂壶藻的研究开发可以从以下几个方面开展:
(1)筛选优良藻种和进一步加强裂壶藻培养条件的优化研究。裂壶藻的营养水平,尤其是蛋白质和不饱和脂肪酸、氨基酸的水平受诸多因素的影响,通过控制氮源、碳源种类或比例,改变温度、pH值等培养条件,添加适当的无机盐、维生素、生物素、苹果酸、柠檬酸、乙醇胺等添加剂可以改变蛋白质、氨基酸、脂肪含量,尤其是提高DHA含量。可以采用诱变的方式定向筛选高产油藻株、高蛋白藻株,实现裂壶藻产物规模化稳定高效生产。
(2)进行裂壶藻作为饲料蛋白源和油脂替代物的研究。这对于降低饲料生产成本,提高养殖效益具有非常重要的意义。裂壶藻中的DHA等不饱和脂肪酸存在于细胞内,不易被氧化,使用比较方便,相对鱼油来说,没有鱼腥味,可全年发酵生产,生长周期短,不受季节影响,裂壶藻有望成为生产多不饱和脂肪酸的良好来源[58]。但是由于裂壶藻油脂主要存在于由细胞壁包裹的细胞内,在饲粮中添加量过多,动物很难将其完全消化分解,影响养殖动物对其有效利用,可以使用破壁的藻类,膜过滤高压均质法破壁效果最好,破壁率可达到100%[22,65]。VE具有抗氧化功能,可以进一步开展破壁裂壶藻和VE等抗氧化物的联合应用研究,从而提高裂壶藻的利用率。
(3)进行裂壶藻体内角鲨烯、虾青素、β-胡萝卜素等生物活性物质的开发研究。虽然其在裂壶藻中的含量很少,但可以通过控制发酵条件、筛选诱变株等方式,提高这些物质的产量,扩大裂壶藻的应用价值。
(4)进行裂壶藻在改善动物产品品质方面的研究。裂壶藻可以抑制小鼠高脂饮食引起的脂肪堆积,因此在功能性添加剂开发方面具有潜在的应用价值。
5 小结
裂壶藻是一种具有重要应用价值的微藻,含有蛋白质、脂肪、糖类、灰分等多种营养物,必需氨基酸种类齐全,尤其是DHA等不饱和脂肪酸含量丰富,另外含有多糖、活性肽等具有免疫活性物质。无机盐类、碳源、氮源、温度、溶解氧、pH值、添加剂等对裂壶藻生长和发酵产物的积累具有非常重要的影响。裂壶藻添加到饲料中能够促进动物存活或生长,提高机体脂肪代谢能力,提高机体非特异性免疫能力,增加机体对DHA营养物的积累,提高营养价值。随着研究的不断深入,裂壶藻在动物生产中将得到越来越广泛的应用。
