小胶质细胞参与新生儿缺氧缺血性脑损伤所致突触异常的研究*
2021-04-06闵颖俊赵俊雄曾祥菲华海蓉毛榕榕金会艳
闵颖俊, 赵俊雄, 彭 行, 曾祥菲, 颜 琳, 华海蓉,毛榕榕, 金会艳, 李 凡△
(昆明医科大学 1病理学与病理生理学系,2临床医学系,3基础医学实验教学中心机能学实验室,云南昆明650500)
新生儿缺氧缺血性脑损伤(hypoxic-ischemic brain damage,HIBD)是导致儿童神经系统伤残的常见原因之一。近年来新生儿重症医学发展迅速,HIBD 患儿存活率显著提高,但幸存患儿因HIBD 所致脑损伤却依然存在,包括脑瘫、癫痫、认知功能障碍等[1-3],严重影响患者的生活质量,给家庭和社会带来极为沉重的负担。脑损伤发生后,可直接因缺氧缺血导致神经元受损、突触异常增多或者减少,也可因缺氧缺血激活免疫系统,间接导致神经系统出现异常的突触修剪,进而导致患者出现认知障碍[4]。
小胶质细胞(microglia,MG)作为脑内主要的免疫细胞,在生理情况下通过释放活性氧和细胞因子,清除凋亡的细胞碎片,同时受到补体CR3 及C1q 的调控,在发育脑内可以影响突触数量,发挥突触修剪作用,对维持中枢神经系统内环境稳定及形成神经环路尤为重要[5-8]。MG 活化是HIBD 的核心发病机制之一,当脑内由于缺氧、缺血、炎症等原因发生病理改变时,MG 将被激活,表现为数量增多和功能活跃,过度激活的MG 在清除死亡细胞碎片的同时,也可产生大量炎症因子,造成神经毒性作用,损伤中枢神经系统的正常细胞,造成神经元损伤及突触结构和功能破坏。目前的研究认为发育脑内MG 与外周血单核细胞源性巨噬细胞(monocyte-derived macrophage,MDMs)共存,二者可共享传统意义上MG 的标志物CD11b[9],这提示脑内CD11b+细胞的变化并不能完全代表脑内驻留MG 的变化。然而目前对于这两种细胞是否均参与了HIBD 后脑内的吞噬活动,是否均影响HIBD 后突触蛋白清除的过程尚不清楚。有研究报导CX3CR1 在脑内仅表达于MG[10],常被用于脑内驻留MG 的特异性标志物[11]。为研究脑内驻留巨噬细胞-MG 在HIBD 后对突触的作用,本研究利用CX3CR1GFP小鼠构建HIBD模型,试图揭示MG通过吞噬参与HIBD后突触异常可能的作用机制。
材料和方法
1 实验材料
1.1 实验动物 用于本实验的C57BL/6J小鼠,动物许可证编号为SCXK(滇)2015-0002,由昆明医科大学动物实验学部提供及饲养。CX3CR1GFP小鼠购自Jackson Laboratory(No.005582),幼鼠6~10只与母鼠同笼饲养,保证食物及水供应,光照时间为7:00~19:00。对实验动物的饲养及处理,均遵守国际医学科学组织理事会制定的国际动物研究指导原则(1985),旨在减少实验动物的数量并减轻其痛苦。
1.2 实验材料 吸入用七氟烷(喜保福宁,进口药品注册证号为H20150020,日本丸石制药株式会社);水合氯醛(Solarbio,Cat:T8590);氯化三苯基四氮唑(2,3,5-triphenyltetrazolium chloride,TTC;Sigma,Cat:93140);多聚甲醛(天津市瑞金特化学品有限公司);溶酶体相关膜蛋白1(lysosomal-associated membrane protein 1,LAMP1;Novus,Cat:NB120-19294);髓系细胞触发受体2(triggering receptor expressed on myeloid cells 2,TREM2;Millipore,Cat:MABN755);神经元核抗原(neuronal nuclear antigen,NeuN;Millipore,Cat:ABN90);4′,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI;Dojindo,Cat:D212);突触后致密蛋白95(postsynaptic density protein 95,PSD95;CST,Cat:36233);突触蛋白(synapsin,SYP;CST,Cat:9020);荧光II 抗购自Life;Fluoro-Jade B(FJB)染液(Merck Millipore,Cat:AG310);Western blot 用II 抗购自博奥森公司。石蜡切片机(Leica,型号:RM2235);正置显微镜(Olympus,型号:BX53);冰冻切片机(Leica,型号:CM1950);荧光共聚焦显微镜(Nikon,型号:A1)。
2 实验方法
2.1 动物模型的建立 本研究采用同窝出生的9~11日龄C57BL/6J及CX3CR1GFP小鼠幼崽,通过改良的Rice-Vannucci 法建立HIBD 模型[12],操作步骤如下。实验动物随机分为缺氧缺血(hypoxia-ischemia,HI)组和同龄对照(control,Con)组。HI组通过七氟烷吸入麻醉(每只30 μL)仰卧位固定四肢,在一种小型实验动物手术箱(专利号:ZL 201520214776.3)中进行左侧颈总动脉结扎手术,术后将动物放回母鼠身边休息1.5 h,休息结束,放入一种改进的恒温缺氧模型实验装置(专利号:ZL 201320101770.6)中,保持温度为34.0℃~34.5℃,在8% O2+92% N2下暴露45 min。Con组无缺氧处理,其余条件与HI组一致。
2.2 TTC 染色 取造模后3 d 小鼠及同龄对照组小鼠,腹腔注射10%水合氯醛(3 mL/kg)麻醉后取脑组织,脑组织置于-20℃冰箱中冰冻10~15 min,待脑组织变硬后进行切片,脑组织由前额至小脑切成6 片,切片厚度为1 mm,切好的脑片放入提前预热到37℃的1%TTC 染液中,避光水浴约15 min,染色结束后,将脑片置于4%多聚甲醛溶液中固定过夜后拍照。
2.3 体重称量 于小鼠结扎颈总动脉造模当天(术前)至HI 后3 d,每天晚上19 点称量各组别小鼠体重,记录每天体重变化情况,并绘制体重增长曲线,观察C组及HI组的体重增长情况。
2.4 抓握实验[13]将HI 后1~3 d 的小鼠及同龄对照小鼠的前爪搭置在细铁丝上,观察小鼠运动功能协调性。评分标准:跌落为0 分;一只或两只前爪紧紧抓住细铁丝为1 分;试图爬上铁丝为2 分;一只或两只前爪和一只或两只后爪紧紧抓住细铁丝为3分;前爪和后爪紧紧抓住细铁丝和尾巴缠绕铁丝为4分;逃跑为5分。
2.5 悬吊实验[14]将HI 后1~3 d 的小鼠及同龄对照小鼠的两只前爪紧紧抓住离地面高度为45 cm 的细铁丝,记录小鼠悬吊在细铁丝上的时间。评分标准:1 分≤10 s;2 分≤11~30 s;3 分:31 s~2 min;4分:3~5 min;5分>5 min。
2.6 HE 染色及Fluoro-Jade B(FJB)染色 取HI 后3 d的小鼠及同龄对照组小鼠,10%水合氯醛腹腔注射麻醉后,经4%多聚甲醛溶液灌流后取脑组织,浸入10%甲醛溶液中固定7 d,经梯度乙醇脱水、浸蜡包埋成蜡块,切片后进行常规HE 染色(晾干后正置显微镜下观察)或FJB 染色(晾干后荧光显微镜下观察)。
2.7 Western blot 检测目标蛋白水平 取HI 后3 d的小鼠及同龄对照组小鼠,10%水合氯醛腹腔注射麻醉后,迅速分离脑组织,取出皮层,放入预冷的1.5 mL EP 管内,根据组织重量,加入裂解液,进行研磨,并4℃离心,收集蛋白上清液,经BCA 测定浓度后,每组取处理好的蛋白样品20 μg(10 μL)进行电泳,并将蛋白质转移到聚偏二氟乙烯膜上。待转膜结束后,使用5%脱脂牛奶封闭1 h 后,加入相应的I抗,室温孵育1 h,4℃过夜。第2 天加入含有辣根过氧化物酶的II 抗,室温孵育2 h。经Bio-Rad Chemi-Doc XRS+显影系统扫描后,用ImageJ 6.0 软件测量目标蛋白及内参照的灰度值。
2.8 免疫荧光染色 取HI 后3 d 的小鼠及同龄对照组小鼠,10%水合氯醛腹腔注射麻醉,经2%多聚甲醛溶液灌流后取脑组织,浸入2%多聚甲醛溶液中固定,4℃过夜,先后经15%及30%蔗糖脱水沉底,用液氮速冻组织样本,保存在-20℃冰箱备用,利用冰冻切片机切片后进行免疫荧光化学染色,具体步骤如下:-20℃拿出玻片,室温晾干→免疫组化笔画圈→PBS-T 洗10 min×3 次→5%驴血清室温封闭2 h→滴加Ⅰ抗,4℃过夜→PBS 洗10 min×3 次→滴加相应的Ⅱ抗,室温1 h→PBS 洗10 min×3次→滴加DAPI工作液,室温3 min→PBS 洗10 min×3 次→荧光抗淬灭剂封片,4℃保存,激光共聚焦显微镜观察。
3 统计学处理
使用GraphPad Prism 6 软件进行统计分析。所有数据表示为均数±标准误(mean±SEM)。对各组数据进行正态性和方差齐性检验,若数据呈正态分布,则采用两独立样本t检验;若数据不符合正态分布,则采用Mann-WhitneyU检验。以P<0.05 为差异有统计学意义。
结果
1 HIBD后小鼠脑组织病理形态学、体重及行为学的变化
造模后3 d,取小鼠脑组织进行TTC 染色,对照组脑组织切片均匀红染,HI 组脑组织于损伤同侧出现明显液化坏死灶(图1A,黄色虚线条所示区域)。小鼠用4%多聚甲醛经左心室穿刺灌流后,观察大脑大体形态,对照组可见两侧大脑半球大小均等,呈淡黄色;HI 组可见损伤同侧大脑半球有呈白色的坏死灶(图1B,黄色虚线条所示区域)。HE染色结果则显示,HIBD 后3 d,脑皮层及海马CA2 区质地疏松、染色变浅,神经元胞体变小,核深染,出现核固缩、核碎裂、核溶解等现象(图1C 中绿色箭头所示),提示复制模型成功。随后我们观察了HIBD术前至HIBD后3 d小鼠及同龄对照组小鼠的体重变化:造模当天HI组小鼠及同龄对照组小鼠术前体重差异无统计学显著性,但HIBD 后1~3 d 的HI 组小鼠体重显著低于同龄对照组小鼠(图1D),其抓握实验评分也显著低于同龄对照组小鼠(图1E)。在悬吊实验中,HIBD 后1 d 的HI 组小鼠悬吊实验评分显著低于同龄对照组小鼠,HIBD 后2 d 及3 d 的HI 组小鼠悬吊实验评分与同龄对照组相比虽有所降低,但差异无统计学显著性(图1F)。
2 HIBD后神经元损伤及突触蛋白异常表达
造模后3 d,取小鼠脑组织进行FJB染色,对照组脑组织切片未见阳性细胞,HI 组脑组织于损伤同侧皮层及海马出现阳性细胞,表明HIBD 后皮层及海马存在坏死神经元(图2A,黄色箭头所示)。小鼠脑皮层的Western blot 结果显示,HIBD 后3 d 脑皮层神经元的NeuN 表达较同龄对照组明显下降,这进一步证实HIBD 后脑内神经元受损(图2B)。免疫荧光染色也显示,HIBD 后3 d 脑皮层神经元细胞核的标志蛋白NeuN 表达下降(图2C,白色箭头所示),均提示HIBD 后神经元损伤。神经元受损后往往伴随有突触蛋白的表达异常,因此我们进一步研究了HIBD 后脑内突触蛋白的表达情况,发现HIBD 后3 d,与同龄对照组小鼠相比,脑皮层区域,突触后膜蛋白PSD95的表达出现增加,并且聚集在梗死灶边缘区域(图2D,白色虚线为梗死区域,白色箭头所示为增多的突触后膜蛋白),提示HIBD 后脑内突触后膜蛋白异常增加;Western blot的结果也显示,HIBD 后脑皮层内突触后膜蛋白PSD95的表达增多(图2E)。

Figure 1. Pathomorphological changes of brain tissue,body weight and behavioral changes of the mice after HIBD. A:TTC staining of the brain tissue at 3 d after HIBD;B:the gross morphology of the brain at 3 d after HIBD;C:HE staining of the brain tissue at 3 d after HIBD;D:the body weight of the mice after HIBD;E:the grip test score of the mice after HIBD;F:the hanging test score of the mice after HIBD. Con:control mice of the same age;HI:hypoxic-ischemic mice. Mean±SEM. n=3-26. *P<0.05,**P<0.01 vs Con group.图1 HIBD后小鼠脑组织病理形态学、体重及行为学变化
3 HIBD后脑内皮层吞噬蛋白增加
本研究进一步通过对HIBD 后3 d的小鼠脑组织进行Western blot 检测,结果显示,与同龄对照组小鼠相比,HIBD 后3 d 小鼠脑皮层吞噬相关蛋白TREM2 和LAMP1 表达均增加(P<0.05 或P<0.01),提示HIBD后脑皮层内吞噬蛋白表达增加,见图3。
4 HIBD 后脑内髓系细胞——CD11b+细胞吞噬功能的变化
神经元损伤及突触蛋白的异常表达必然破坏脑内环境的稳定,激活免疫系统,MG 作为脑内重要的免疫细胞,在生理及病理情况下都有强大的吞噬能力。为了进一步确定HIBD 后脑内髓系细胞——CD11b+细胞的吞噬能力变化,我们通过免疫荧光多重标记染色发现,HIBD 后3 d,HI 组梗死灶边缘及中心区CD11b+细胞(红色)数量明显增多,部分与神经元标志物NeuN(绿色)及LAMP1(青色)共染(图4,白色箭头所示),部分未与LAMP1 共染(图4,黄色箭头所示),提示HIBD 后脑内部分CD11b+细胞出现吞噬神经元的现象。
5 HIBD 后脑内驻留巨噬细胞——MG 参与吞噬突触
为进一步确认MG是否参与了吞噬突触的过程,本研究利用MG 被特异性标记了绿色荧光的小鼠——CX3CR1GFP转基因小鼠建立HIBD 模型,通过免疫荧光共聚焦扫描发现,HIBD 后3 d,脑内MG 数量增多,胞体变圆,且TREM2 表达增加(图5A,白色箭头所示),提示HIBD 后MG 活跃,吞噬能力增加。为进一步确认MG是否参与了吞噬突触,本研究进一步观察了HIBD 后MG 胞内SYP 的变化,结果表明,HIBD 后3 d,MG 胞内出现了SYP 的表达(图5B,黄色箭头所示),提示HIBD后MG参与了突触吞噬。

Figure 2. Neuronal injury and increase in postsynaptic membrane protein after HIBD. A:Fluoro-Jade B(FJB)staining of cerebral cortex and hippocampal CA2 region in mice at 3 d after HIBD;B:the expression of NeuN in the cerebral cortex of mice detected by Western blot at 3 d after HIBD;C:the NeuN immunofluorescence staining results in the brain at 3 d after HIBD;D:the immunofluorescence staining results of brain postsynaptic membrane protein PSD95 at 3 d after HIBD;H:the expression of PSD95 in the cerebral cortex of mice detected by Western blot at 3 d after HIBD. Con:control mice of the same age;HI:hypoxic-ischemic mice. Mean±SEM. n=3-5. *P<0.05 vs Con group.图2 HIBD后神经元损伤及突触后膜蛋白增多

Figure 3. Increased expression of cortical phagocytic protein after HIBD. A:the expression of phagocytosis-related molecule TREM2 in cortex of mice detected by Western blot at 3 d after HIBD;B:the expression of phagocytosis-related molecule LAMP1 in cortex of mice detected by Western blot at 3 d after HIBD. Con:control mice of the same age;HI:hypoxic-ischemic mice.Mean±SEM. n=5. *P<0.05,**P<0.01 vs C group.图3 HIBD后脑皮层吞噬蛋白表达增多
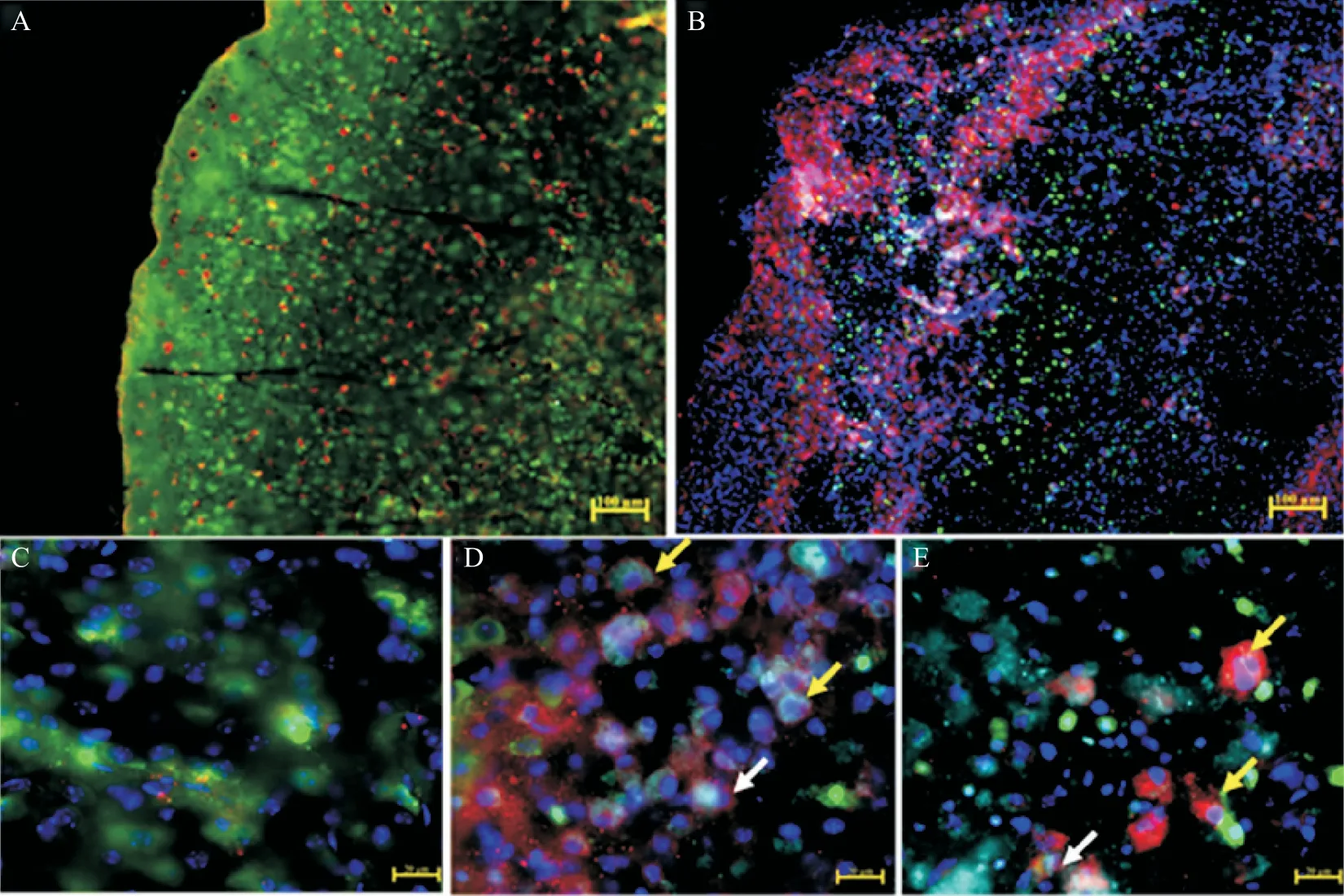
Figure 4. Changes of macrophage phagocytosis in the brain after HIBD. The immunofluorescence staining results of neurons,macrophages and phagocytosis-related proteins in the brain of mice at 3 d after HIBD were shown(scale bar=100 μm in A and B;scale bar=20 μm in C,D and E). A and C:control group;B,D and E:HIBD group. Part of CD11b+cells(red)were costained with neurons(green)and LAMP1(cyan),as shown by the white arrows in D and E. The yellow arrow showed that some CD11b+cells(red)were not co-stained with LAMP1(cyan). Blue represented DAPI.图4 HIBD后脑内巨噬细胞吞噬情况变化
讨论
HIBD 是导致新生儿死亡以及患儿远期行为出现异常的主要原因之一,远期行为异常与神经元丢失、突触数量与结构变化密切相关。MG 作为脑内最为重要的固有免疫细胞[15],除通过释放炎症因子参与脑损伤后的神经炎症反应外,还可通过吞噬作用清除脑内的细胞碎片及轴突碎片参与维持内环境稳定。HIBD后突触损伤的方式及MG对突触蛋白的吞噬作用尚未见报道。本模型的受损区域主要为皮层及海马神经元,伴随着MG 的活化,其吞噬功能的改变,清除脑内细胞碎片与轴突碎片的同时,过度活化的MG 也有可能损伤正常神经元。突触作为神经元的基本结构单位,是学习和记忆的基础,神经元的损伤往往会导致突触数量和结构的变化,引起患儿出现学习障碍和行为异常。因此深入研究HIBD 后神经元和突触的变化,了解MG 的吞噬功能改变,尤其是对其吞噬突触的作用,对于了解HI 后MG 在脑损伤过程中发挥的作用尤为重要,并可为临床治疗HIBD患儿提供潜在的治疗靶点。
神经元及其突触是维系行为正常的必要基础,本研究首先证实了HIBD 造成的行为异常及突触丢失情况。结果揭示HIBD 后小鼠体重出现明显下降,HIBD 后1~3 d 小鼠抓握实验评分出现明显下降,而悬吊实验则仅在HIBD 后1 d 出现评分下降,提示小鼠在HIBD 后出现了行为异常。HE染色发现,HIBD后小鼠脑皮层及海马区域神经元出现坏死典型的病理改变,与多数人的研究一致[12,16]。我们进而通过Western blot、FJB 染色及免疫荧光染色揭示,HIBD后坏死神经元数量明显增多,正常神经元数量明显低于同龄对照组,表明HIBD 后3 d 小鼠脑内出现了明显的神经损伤。神经元胞体的受损往往伴随有其突起(轴突及树突)的损伤[13],轴突及树突是形成神经突触的重要结构,突触又是信息传递的基础,对于构建大脑内精确的神经环路尤为重要,有研究报道低氧可以诱导海马CA1神经元树突棘形态结构发生变化,最终影响其学习和记忆[14]。这些结果表明,HIBD后出现了突触的丢失。
构成突触的蛋白主要包括突触前膜及突触后膜蛋白,突触蛋白的表达与突触损伤未必一致。2019年Griffiths 等[17]的研究发现外伤性脑损伤后,突触后膜蛋白表达增多,也与癫痫模型海马损伤后模型相似[18]。还有研究报道在低冲击爆炸所致的脑损伤模型中,脑皮层突触数量的减少伴随有突触蛋白厚度增加,而海马则相反,提示损伤后脑内发生了潜在的突触重构[13]。本研究的结果表明HIBD 后大脑皮层内突触后膜蛋白PSD95 的表达明显增多,增多的PSD95 蓄积在梗死灶的边缘区域,可见HIBD 后突触的损伤形式为突触的直接丢失及突触后膜蛋白PSD95 的过表达,其如何参与突触重构的机制还有待进一步研究。
我们还探讨了HIBD 后脑内驻留巨噬细胞MG的吞噬作用是否参与HIBD 后突触损伤及其可能的作用机制。LAMP1表达增加往往意味着细胞吞噬活动增强[19]。TREM2 则是参与调控MG 吞噬的主要分子,但其在HIBD 中的作用未见报道。本研究的结果显示,HIBD 后3 d 皮层吞噬相关蛋白LAMP1 及TREM2 表达明显增多;免疫荧光共定位结果显示,HIBD 后3 d 梗死灶边缘区域LAMP1 的表达增加,但并不完全表达在CD11b+细胞内,且此区域NeuN的表达仍呈现下降,同时部分溶酶体活跃的CD11b+细胞出现吞噬神经元的现象。这些结果提示,HIBD 后,尤其重度损伤组脑内CD11b+细胞具有活跃的吞噬功能,并与HIBD后神经元突触的丢失有关。
近年来,大量研究证实脑损伤后脑内CD11b+细胞可以分为来自于外周入侵的单核细胞及来自于中枢的MG 两群细胞[20-22],提示脑内CD11b+细胞的变化并不能完全代表脑内驻留MG的改变。因此,为了进一步确定HIBD 后脑内MG 的吞噬能力变化及其与突触的关系,本研究通过CX3CR1GFP转基因小鼠构建了HIBD 模型。结果发现,HIBD 后3 d 脑内MG(绿色荧光)胞内TREM2 表达较对照组增多,提示HIBD后脑内MG 吞噬能力增加,这与Fisch 等[23]及谌贝贝等[24]的研究一致。正常发育脑内,MG 通过吞噬作用参与突触修剪帮助建立精确的神经环路。HIBD后MG 的吞噬功能异常活跃,是否会影响其突触修剪,导致患者出现行为异常?本研究证实,HIBD 后3 d 的MG 胞内出现更多的突触前膜蛋白,这证明了HIBD 后MG 直接参与吞噬突触,这将影响神经元突触结构。2017年Aono等[25]在帕金森病中发现MG 吞噬丘脑底核谷氨酸能突触,但HIBD 后MG 吞噬突触的研究未见报道。因此,本研究证实了HIBD 后脑内驻留巨噬细胞MG 吞噬功能激活,并通过TREM2 上调吞噬功能,介导突触异常,进而在HIBD 后的行为异常中发挥作用。但MG 吞噬完整突触的方式和相关信号通路,及其可能的干预策略还有待进一步研究。
