大陈洋产卵场保护区鱼卵、仔稚鱼的种类组成及其与环境因子的关系
2021-03-31于春梅蒋日进李鹏飞朱海晨
于春梅,蒋日进,印 瑞,李鹏飞,朱海晨
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,农村农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
鱼类生活史包括鱼卵、仔稚鱼、幼鱼和成鱼4 个时期,其中鱼卵、仔稚鱼是鱼类生命周期中最为敏感的阶段,其数量变动和后续成长态势是鱼类资源得以补充及可持续利用的基础[1]。鱼卵、仔稚鱼作为浮游动物的重要组成部分[2],不仅在海洋食物网中起着重要的作用,而且对近岸海域生态系统功能的可持续发展也具有重要意义[3]。
产卵场是鱼类繁殖过程中的栖息场所,鱼类通过繁殖与其它环节紧密联系,保证了物种的增殖和延续,从而达到补充和增殖群体数量的目的[4]。20 世纪90 年代以来,我国海洋捕捞技术发展迅速,渔业资源被过度利用,传统经济鱼类数量急剧下降,带鱼Trichiurus lepturus 等经济鱼类渔汛逐步消失[5]。为保护近海主要经济鱼类的产卵场,2017 年浙江省在全国首次批复建立了10 个产卵场保护区,主要保护带鱼、小黄鱼Larimichthys polyactis、大黄鱼L.crocea 等重要经济种类,保护区内禁渔期开始时间相比伏季休渔时间提前至4 月1 日,结束时间不变,为经济鱼类资源提供了更长的繁殖和栖息时间。大陈洋产卵场保护区位于浙江省台州市东部海域,该海域受台湾暖流、椒江冲淡水及沿岸流的交汇影响,饵料生物资源丰富,为众多海洋生物提供了良好的栖息场所[6]。目前已有学者初步研究了中国沿海地区包括黄渤海、南海等海域鱼卵仔稚鱼的群落结构[7-9],发现温度、盐度等环境因子对鱼卵的影响较大。涉及鱼卵仔稚鱼与环境因子的研究方法主要有冗余分析(RDA)[10]、广义加性模型(GAM)[11]、广义线性模型(GLM)[12]等,但有关鱼类产卵场保护区鱼卵仔稚鱼的分布变化的研究较少。因此对2018 年4、5、11 月大陈洋产卵场保护区鱼卵、仔稚鱼的群聚结构等方面进行研究,旨在为产卵场保护区效果评价提供研究基础与参考,可进一步丰富东海区鱼类早期生活史研究,并为该海域生态环境修复和生物资源合理利用提供科学依据。
1 材料与方法
1.1 调查站位及样品采集
本实验分别于2018 年春季(4 月、5 月)、2018 年秋季(11 月)对大陈洋产卵场保护区鱼卵、仔稚鱼进行调查,共设15 个站位点(图1)。依据《海洋调查规范》(GB/T 12763.6-2007)[13]采集鱼卵、仔稚鱼,具体方法如下:采用口径80 cm、网目尺寸0.505 mm 的大型浮游生物网逐个站点进行表层水平拖网采样,以2 kn 拖速匀速拖网10 min,网口配备流量计;采用多功能水质检测仪(JFE-AAQ171),对水深、温度、溶解氧和盐度等相关环境因子进行数据采集。采用大型浮游生物网采集鱼卵、仔稚鱼样品,使用5%的甲醛溶液将样品固定,带回实验室进行鉴定和计数。使用Leica M205 C 体视显微镜,参考《日本产稚鱼图鉴》[14]、《中国近海鱼卵与仔鱼》[15]、《中国近海及其邻近海域鱼卵与仔稚鱼》[16]、《东海鱼类志》[17]等书籍,根据鱼卵、仔稚鱼的形态特征,将其鉴定到最低分类阶元。
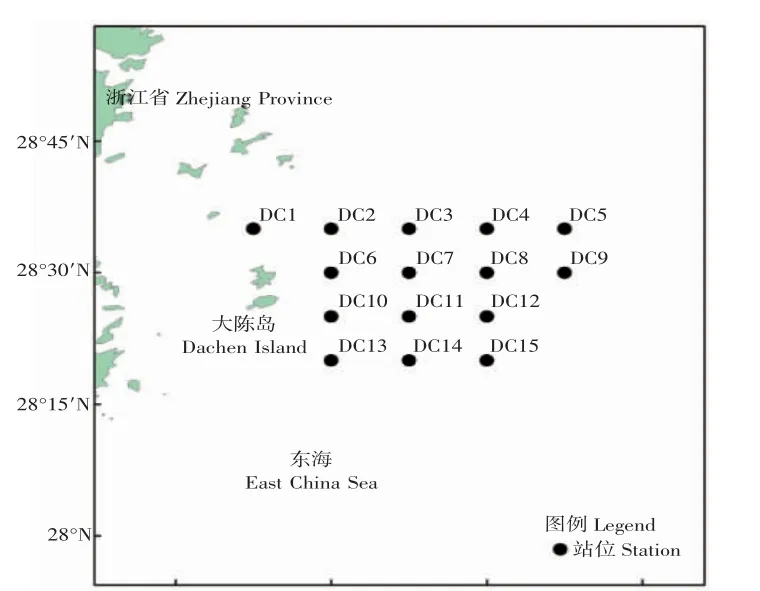
图1 大陈洋产卵场保护区鱼卵、仔稚鱼调查站位Fig.1 Sampling station of ichthyoplankton in Dachenyang spawning ground reserve
1.2 数据处理与分析方法
1.2.1 生态优势度
根据每网采集的鱼卵、仔稚鱼的实际数量(粒、尾)计算其密度,单位为ind·m-3。
利用PINKAS,et al[18]相对重要性指数(index of relative importance,IRI)计算采集到的鱼卵、仔稚鱼的生态优势度,公式为:

式中:N 为某一种类鱼卵或仔稚鱼尾数密度占采获总尾数密度的百分比,%;F 为某一类出现的站位占总调查站位的百分比,%;优势种范围为IRI 值>1 000,重要种范围为IRI 值100~1 000。
1.2.2 物种多样性指数
采用Shannon-Wiener 多样性指数(H')、Pielou 均匀度指数(J')、Margalef 种类丰富度指数(D)[19]等表示调查海域的物种多样性,其计算公式如下:
香农-威纳指数(H')计算公式为:

丰富度指数(D)计算公式为:

均匀度(J')计算公式为:

式中:S 为总种类数;ni为第i 种鱼卵、仔稚鱼的个体数;N 为总个体数[20]。
1.2.3 环境因子分析
首先,建立GAM 模型之前在R 语言中对环境因子进行了相关分析,根据Pearson 相关分析性结果,共线性强的两个环境因子保留一个。最终对鱼卵的分析选择的因子为:表层水温、表层盐度、水深、叶绿素a含量、浑浊度;对仔稚鱼分析选择的因子为:表层水温、表层盐度、叶绿素a 含量。本研究以大陈洋产卵场保护区鱼卵、仔稚鱼丰度及相关环境因子进行回归分析。GAM 模型的一般形式为[21]。

式中:Y 为鱼卵、仔稚鱼丰度,ind·m-3;α 为函数的规定截距;xi表示各环境因子的解释变量;ε 为随机误差项;fi(xj)是各环境变量xj的任意单变量函数,通过样本平滑函数来估计。模型的误差分布估计为Gaussian 分布,连接函数为自然对数[21]。
采用AIC 最小准则筛选得出最优模型,首先进行单环境因子模型预测,在AIC 值最小的预测模型基础上加入其它因子,之后进行多环境因子的模型预测,重复这一选择过程(即逐步回归),直到AIC 值不再变小,最终AIC 值最小的模型即为拟合效果最好的模型[21]。
本文使用Microsoft Excel 2013 进行数据的预处理;采用ArcGIS 软件绘制大陈洋产卵场保护区站位图,模型的构建通过R 软件中的gam 包来进行。
2 结果
2.1 种类组成
春季共采集到鱼卵、仔稚鱼22 种,隶属于7 目18 科,6 种鉴定到科,1 种鉴定到属(表1)。其中,鲈形目的种类数是最多的,占50%;灯笼鱼目次之,占14%;鲱形目、鲉形目、鲽形目各占9%;鲻形目和鲀形目各占4%。

表1 2018 年大陈洋产卵场保护区鱼卵、仔稚鱼的种类组成Tab.1 Species composition of ichthyoplankton in Dachenyang spawning ground reserve in 2018
秋季共采集到鱼卵、仔稚鱼4 种,隶属于3 目3 科,有1 种鉴定到科。其中,灯笼鱼目种类最多,占50%;鲱形目、鲈形目各占25%。
2018 年共采集到鱼卵、仔稚鱼25 种,隶属于7 目18 科,春季采集到的鱼卵、仔稚鱼种类数大于秋季,其中鲈形目在春季的种类数远高于秋季。
2.2 优势种及其季节变化
根据大陈洋产卵场保护区调查结果计算各鱼卵、仔稚鱼优势度(表2),鱼卵春季、秋季的共有优势种为龙头鱼Harpodon nehereus,秋季龙头鱼的丰度最高,蛇鲻属未定种Saurida sp.的鱼卵占总丰度的比例最高;仔稚鱼春季、秋季无共有优势种,季节更替变化明显。其中龙头鱼仔稚鱼的丰度均为春秋季最高,为绝对优势种。
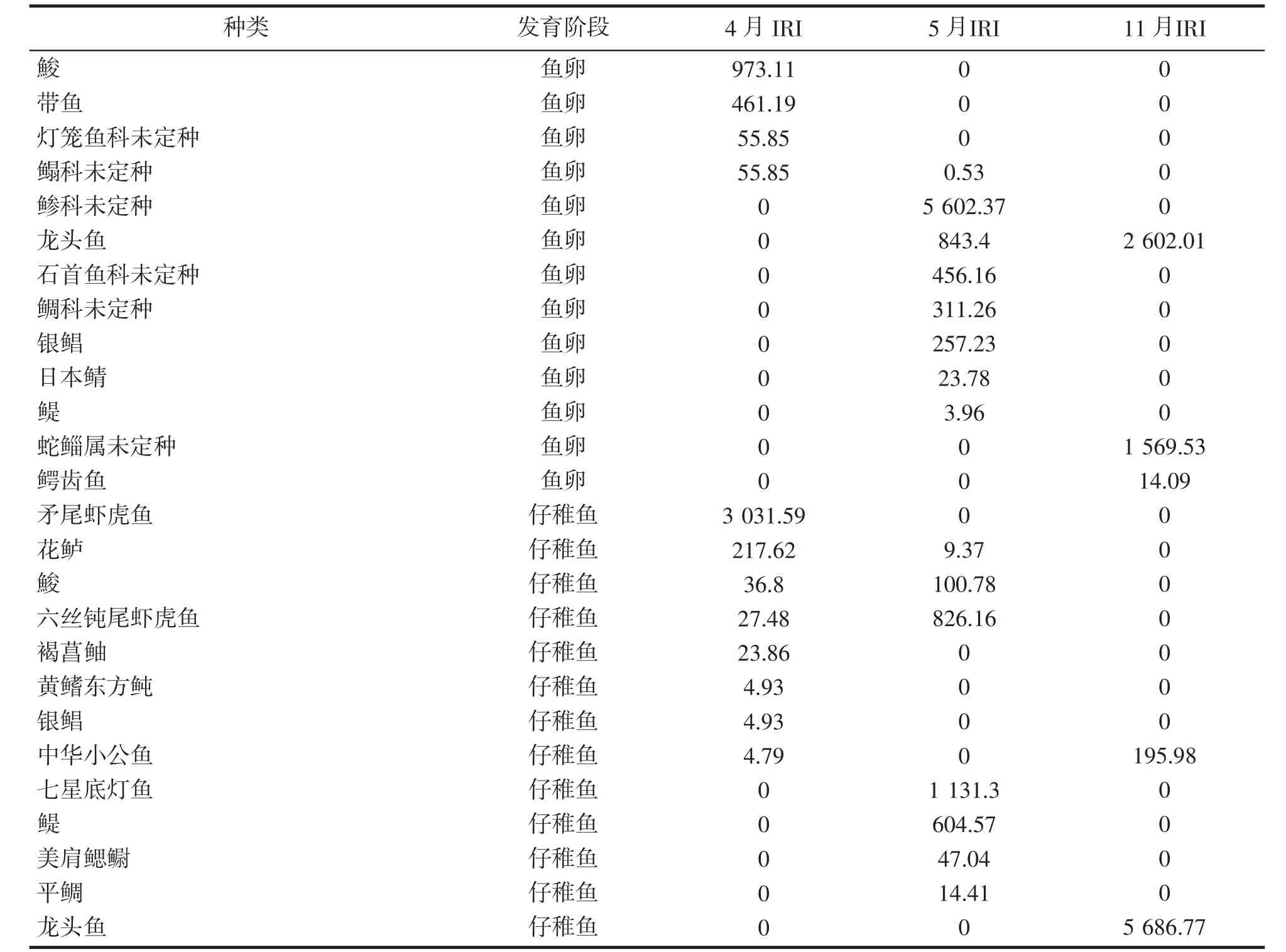
表2 2018 年大陈洋产卵场保护区鱼卵、仔稚鱼优势种和重要种组成Tab.2 Dominant and important species composition of ichthyoplankton in Dachenyang spawning ground reserve in 2018
4 月大陈洋产卵场保护区鱼卵重要种以带鱼和鮻Liza haematocheilus 为主,带鱼鱼卵丰度占总丰度的35%,海域平均密度为0.73×10-3ind·m-3;鮻鱼卵丰度占总丰度的49%,海域平均密度为1.03×10-3ind·m-3;5 月鱼卵优势种重要种以鲹科Carangidae sp.和龙头鱼为主,其中鲹科鱼卵丰度占总丰度的51%,海域平均密度为129.23×10-3ind·m-3;龙头鱼鱼卵丰度占总丰度的14%,海域平均密度为32.43×10-3ind·m-3;11 月鱼卵优势种以蛇鲻属和龙头鱼为主,其中蛇鲻属鱼卵丰度占总丰度的58%,海域平均密度为6.97×10-3ind·m-3;龙头鱼卵丰度占总丰度的39%,海域平均密度为4.62×10-3ind·m-3。
4 月大陈洋产卵场保护区仔稚鱼优势种重要种以矛尾虾虎鱼Chaeturichthys stigmatias 和花鲈Lateolabrax japonicus 为主,其中矛尾虾虎鱼丰度占总丰度的76%,海域平均密度为18.62×10-3ind·m-3;花鲈丰度占总丰度的16%,海域平均密度为4×10-3ind·m-3;5 月仔稚鱼优势种重要种以七星底灯鱼Benthosema pterotum 和六丝钝尾虾虎鱼Amblychaeturichthys hexanema 为主,其中七星底灯鱼丰度占总丰度的42%,海域平均密度为4.14×10-3ind·m-3;六丝钝尾虾虎鱼丰度占总丰度的24%,海域平均密度为2.42×10-3ind·m-3;11 月仔稚鱼优势种为龙头鱼,丰度占总丰度的90%,海域平均密度为5×10-3ind·m-3。
2.3 物种多样性指数
鱼卵物种多样性指数(H')变化范围为1.055~2.056,最大值出现在5 月,数值为2.056。11 月多样性指数明显低于5 月份,数值为1.055,这与蛇鲻属和龙头鱼等优势种类大量产卵有很大关系,优势度达到16.15%和15.21%。均匀度指数(J')变化范围为0.666~0.861,11 月最低,数值为0.666;4 月均匀度指数最高,与鱼类产卵量普遍少,数量比较平均有关。丰富度指数(D)变化范围为0.469~1.303。
仔稚鱼物种多样性指数(H')变化范围为0.454~2.182,最大值出现在5 月,最小值出现在11 月;均匀度指数(J')变化范围为0.395~0.777;丰富度指数(D)变化范围为0.328~1.763(图2)。
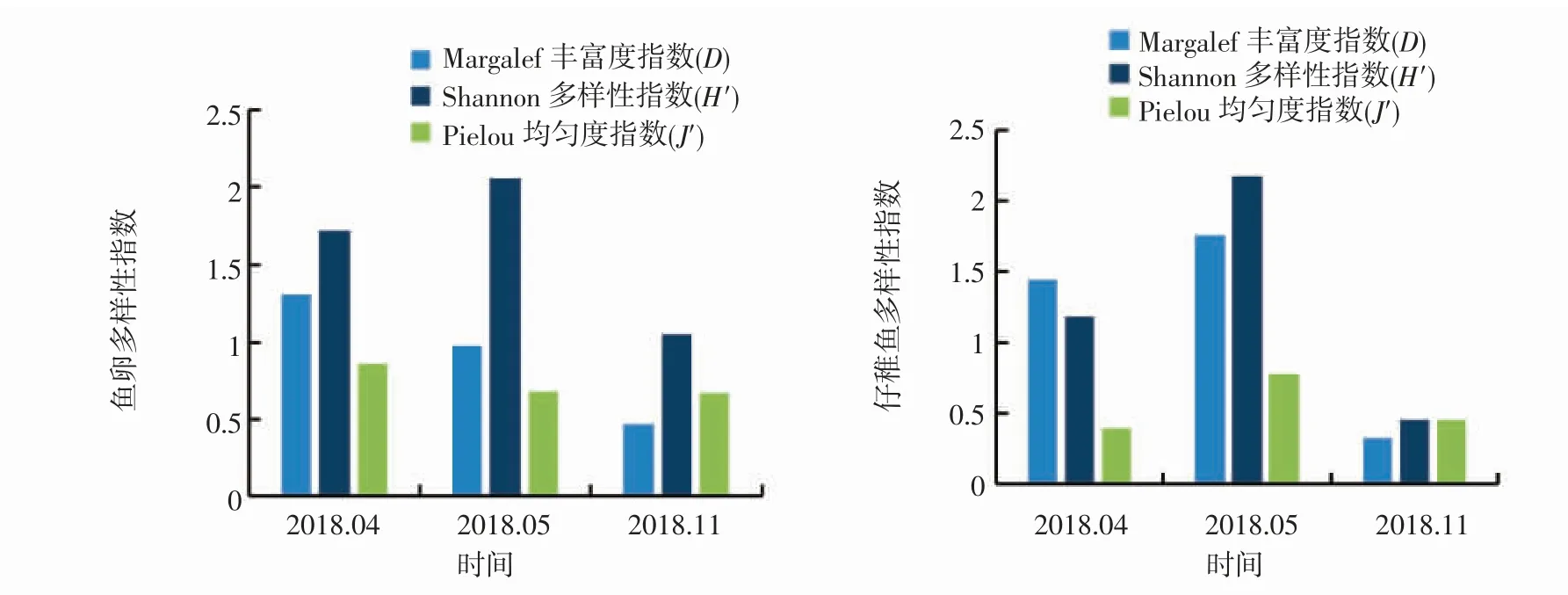
图2 2018 年大陈洋产卵场保护区鱼卵、仔稚鱼物种多样性指数Fig.2 Species diversity index of ichthyoplankton in Dachenyang spawning ground reserve in 2018
基于GAM 模型对鱼卵、仔稚鱼尾数密度(density) 与环境因子间的关系进行拟合。根据AIC(akaike information criterion)最小原则筛选出最优模型(表3~6)。得到鱼卵的最优模型表达式为:Lg(density+1)=α+s(SST)+s(SSS)+s(Chl.a)。仔稚鱼的最优模型表达式为:Lg(density+1)=α+s(SST)+s(Chl.a)。结果显示,表层水温s(SST)和表层盐度s(SSS)对鱼卵丰度变化具有显著影响(P<0.05),表层水温对仔稚鱼丰度变化具有显著影响(P<0.05)。
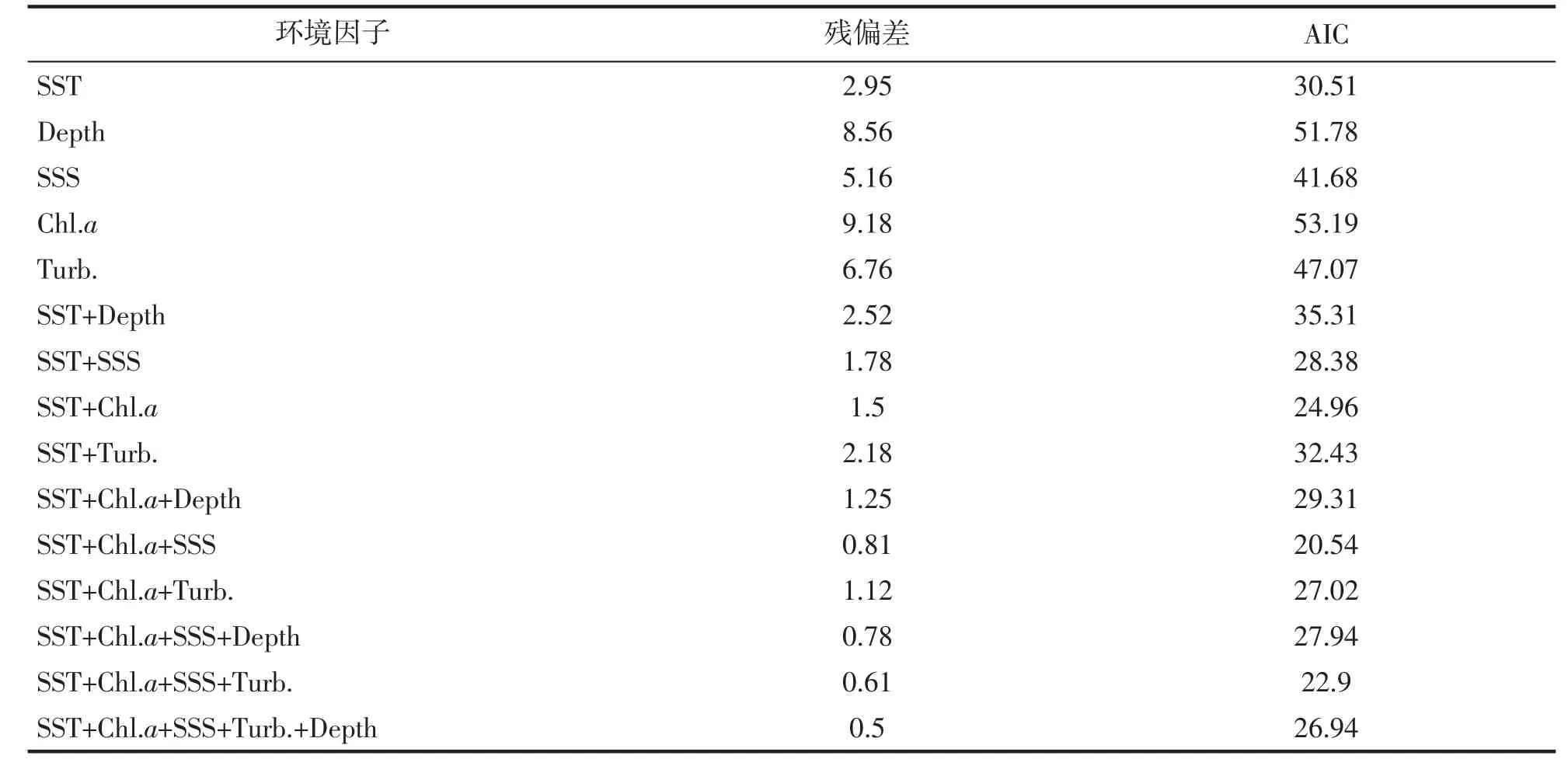
表3 鱼卵GAM 模型变量筛选及影响因子的参数分析。Tab.3 The variable screening process for GAM and parameters analysis of fish eggs

表4 鱼卵GAM 模型拟合结果的偏差分析Tab.4 The analysis of deviance for GAM of fish eggs
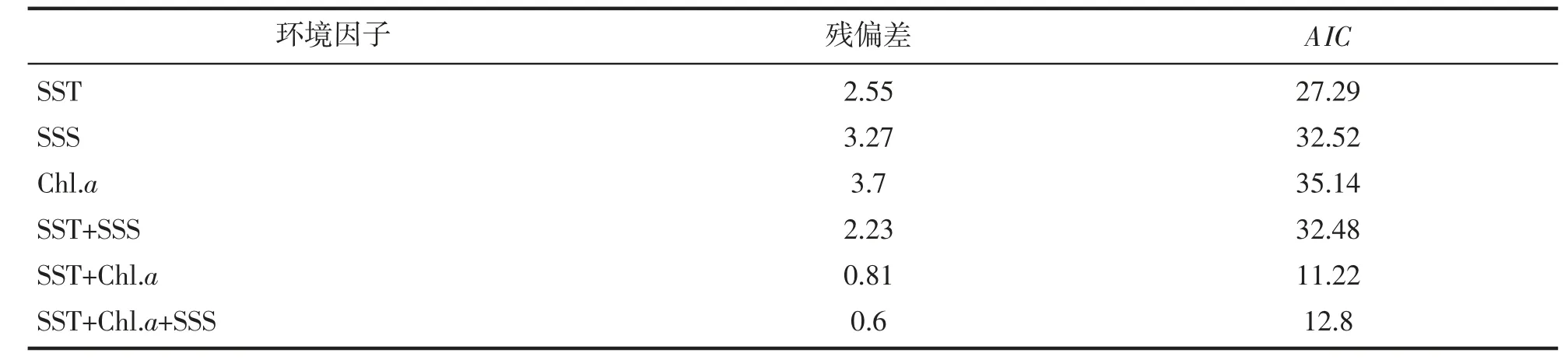
表5 仔稚鱼GAM 模型变量筛选及影响因子的参数分析。Tab.5 The variable screening process for GAM and parameters analysis of fish larvae

表6 仔稚鱼GAM 模型拟合结果的偏差分析Tab.6 The analysis of deviance for GAM of fish larvae
由图3 所示,表层水温、表层盐度、叶绿素a 含量对鱼卵丰度存在单波谷的非线性相关关系。表层水温与鱼卵丰度之间存在正相关关系,随着温度的升高,鱼卵丰度呈现逐步上升的趋势。在表层水温为17 ℃时出现鱼卵丰度的低值点。当表层水温超过17 ℃之后,鱼卵丰度基本随表温增加而增加。表层盐度对于鱼卵丰度的影响小于表层水温的影响,在表层盐度为30.5 左右出现鱼卵丰度的最低值。鱼卵丰度随着叶绿素浓度的升高呈现先增后降的趋势,在5~10 左右,叶绿素a 含量与鱼卵丰度存在负的非线性相关关系。鱼卵丰度在叶绿素浓度为10 时,鱼卵丰度降为最低值。
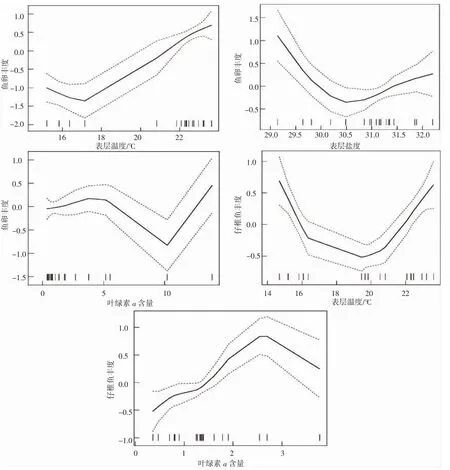
图3 不同因子对大陈洋产卵场保护区鱼卵、仔稚鱼丰度的影响Fig.3 Effects of influential factors on relative density of ichthyoplankton in Dachenyang spawning ground reserve
表层水温对仔稚鱼丰度存在单波谷的非线性相关关系。表层水温对大陈洋产卵场保护区仔稚鱼丰度的影响最大,随着表层水温的增加,仔稚鱼丰度整体呈现先下降后上升的趋势。表层水温为16~20 ℃时仔稚鱼丰度较低,20~22 ℃之间,表层水温与仔稚鱼丰度之间存在负的非线性相关关系。叶绿素a 含量对仔稚鱼丰度存在单波峰的非线性相关关系。叶绿素a 含量对仔稚鱼丰度的影响小于表层水温的影响,随着叶绿素a 含量浓度的增加,仔稚鱼丰度呈现先上升后下降的趋势,当叶绿素浓度为2~3 时,仔稚鱼丰度较高。
3 讨论
鱼卵、仔稚鱼资源量的密度变化是衡量鱼类生殖群体生产力及资源补充机制的基础[22]。调查发现,春季为鱼类产卵的高峰期,该季节采集到的鱼卵、仔稚鱼的数量占采获总量的94%,远高于秋季,此时对产卵场和繁殖群体进行保护将会有效增加其资源量[23]。仔稚鱼的优势种发生了显著的季节变化,优势种由春季的矛尾虾虎鱼、七星底灯鱼等底层小型鱼类更替为秋季的龙头鱼等鱼类,龙头鱼为中下层鱼类,主要分布在浙江南部海域。近年来,东海近海渔业资源结构发生了显著的变化,群落中底层鱼类如大黄鱼、小黄鱼和带鱼等经济种类的总体资源量不断减少,中小型鱼类如龙头鱼、鳀等逐渐占据主导地位[24]。造成这一结果的原因主要是由于过度捕捞,在浙江实施“一打三整治”、“幼鱼保护”等渔业管理政策之前,“密眼”拖网、流刺网等有害渔具的使用,也导致该区域渔业资源的补充群体遭到非法捕捞[25]。
大陈洋产卵场保护区海域在浙闽沿岸流、台湾暖流及江河冲淡水的综合作用下,水文环境适宜,基础饵料丰富,使多种鱼类在大陈洋产卵场保护区聚集产卵。虽然本次调查带鱼、大黄鱼和小黄鱼等重要经济种类的鱼卵仔稚鱼数量相对较少,多以六丝钝尾虾虎鱼和七星底灯鱼等饵料性小型鱼类的仔稚鱼为主,但是也有一些经济种类如鲹科鱼类鱼卵在2018 年春季大量出现,密度达到1 938.5×10-3ind·m-3,远高于其他物种,鲹科鱼类繁殖力极强、近海暖水性、喜集聚、具有趋光性的中上层鱼类[26],其群体资源量较大。大陈洋产卵场保护区西部毗邻岛礁和河口,一些岛礁种类如平鲷Rhabdosargus sarba、美肩鳃鳚Omobranchus elegans 主要栖息于产卵场保护区的岩礁或礁沙交错处,以虾、蟹、海藻等为食[27-28],二者的摄食习性和栖息环境相似。
由GAM 模型拟合结果可知,温度对大陈洋产卵场保护区鱼卵、仔稚鱼丰度的影响最为显著。温度会直接影响鱼类受精卵的孵化,仔稚鱼的成长代谢,不适的温度也可能会导致鱼卵畸形[29]。同时,每种鱼都有自己狭窄的产卵温度阈,温度升高或者降低会致使某种鱼种产卵提前或者延后[30],因此温度对鱼类早期阶段的影响极大。GAM 模型拟合结果表明,大陈洋产卵场保护区的鱼卵丰度随着温度的升高而上升,温度升高促使鱼类性腺的提前发育[31],保护区内产卵的种类数量也增多[3]。除温度影响外,盐度对鱼卵的影响也很大。盐度直接影响鱼类渗透压的功能,进而对其应激、免疫、消化等生理功能发生作用,对其生长发育有不可忽视的影响[32]。春季,东南季风使得沿岸流向北与台湾暖流向北移动的方向一致,带来高温高盐的海水[33]。大部分鱼类在春季产卵,中华小公鱼Stolephorus chinensis、花鲈等种类分布在低盐区,鲹科等鱼类在高盐海区繁殖[34]。秋季,随着水温降低,很少有鱼类在保护区产卵,但是大黄鱼、龙头鱼、短鳄齿鱼Champsodon snyderi 和中华小公鱼等鱼类仍旧在保护区繁殖[34]。随着叶绿素的增加,鱼卵、仔稚鱼的丰度均呈现先增后减的趋势。大陈洋产卵场保护区水温适宜海藻等浮游植物的生长,光合作用使水体氧气含量上升,浮游动物随氧气含量上升而上升,营造了适合仔稚鱼栖息的良好场所[35]。同时过量的藻类生长会造成水体富营养化,不利于浮游动物的生长繁殖,也会对鱼卵、仔稚鱼造成不利影响。
