嗜盐微生物底盘细胞:应用和前景
2021-03-30马悦原陈金春陈国强
马悦原,陈金春,陈国强,2
(1 清华大学生命科学学院,北京100084;2 清华大学化工系,北京100084)
工业生物技术生产了一些人类所需的化工、材料、能源、酶、食品和医药等产品[1]。但是,除了医药和食品,现代工业生物技术生产的化工、材料和能源等产品制造成本高昂,虽然制造过程安全,但竞争性不如化学工业。目前的工业生物技术主要利用模式微生物,特别是大肠杆菌、假单胞菌、芽孢杆菌、乳酸菌、链霉菌和酵母菌进行,过程有容易染菌、淡水消耗大、耗能高、发酵不连续、细胞密度低和产量低等缺点。为克服目前的工业生物技术的缺点,利用极端微生物进行工业生物制造的“下一代工业生物技术”应运而生。下一代工业生物技术使用嗜盐微生物、嗜冷微生物、嗜热微生物、嗜酸微生物、嗜碱微生物、旱生菌以及在特殊碳源或有毒物质中生长的微生物等特殊微生物进行工业发酵[2],以实现无需灭菌、节能节水、开放式、连续的发酵过程。其中,嗜盐微生物以其在高盐环境中生长的独特优势,成为“下一代工业生物技术”的最先成功开发的底盘细胞[3-4]。
1 嗜盐微生物的特点和优势
嗜盐微生物是一类生长于NaCl浓度大于1%的环境中的微生物,根据生长环境盐浓度的不同,嗜盐微生物可以分为低度嗜盐菌(1%~3%NaCl)、中度嗜盐菌(3%~15% NaCl)和高度嗜盐菌(15%~30%NaCl)[5],除了耐高盐,很多嗜盐微生物还具有耐高碱的特点[6]。嗜盐微生物主要生长于海洋、盐湖等环境中,在分类学上包括细菌、古菌、藻类和真菌, 常见的嗜盐微生物有Halomonas、Haloferax、Alkalibacterium、Bacillus、Pontibacillus和Marinobacter等[7]。
相较于大肠杆菌、假单胞菌、芽孢杆菌、天蓝链霉菌和酵母等底盘细胞,嗜盐微生物底盘细胞的遗传背景不够清晰、基因编辑难度更大,这为嗜盐微生物的研究和改造带来了许多挑战。但是,因为具有耐盐、不易染菌、生长速度快和鲁棒性强等诸多优势(表1),以嗜盐微生物为底盘细胞的“下一代工业生物技术”相较于现有的工业生物技术展现出如下优势:第一,由于其他微生物无法在嗜盐微生物生长所需的高盐高碱环境中生存,嗜盐微生物不易被其他微生物污染,所以在用嗜盐微生物进行工业发酵时,可以省去灭菌步骤,进行无灭菌、开放式、连续发酵,这极大地简化了发酵操作过程,减少了灭菌能耗和人力操作;第二,可以用海水替代淡水作为嗜盐微生物进行工业发酵的高盐浓度培养基,这节约了淡水资源;第三,进行发酵的生物反应器也无需灭菌,可以用塑料、陶瓷等材料制作,无需耐高温耐高压的金属材料;第四,工程化改造后,可以用厨余垃圾、秸秆等廉价碳源替代葡萄糖进行工业发酵;第五,可以用同一株嗜盐菌生产多种产品;第六,可以用全自动电子控制系统自动控制连续发酵过程,取代现有的人工操作。盐单胞菌属菌株Halomonas bluephagenesis TD01 和Halomonas campaniensis LS21 可以在高盐高碱的环境下生长,具有生长速度快、细胞干重大、不易染菌、产品丰富等优势,已经成为“下一代工业生物技术”的重要底盘细胞[2,4,8-10]。
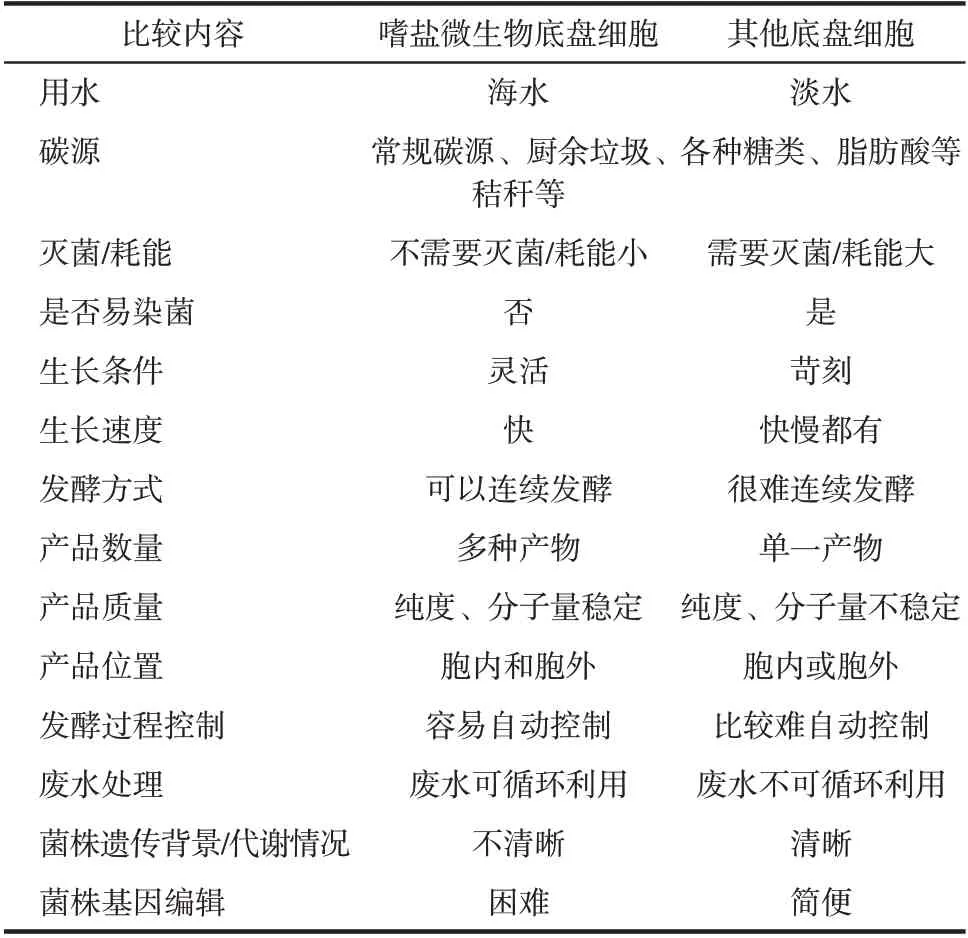
表1 嗜盐微生物底盘细胞与其他底盘细胞的优缺点
2 嗜盐微生物分子操作工具的开发
在工业生产中,提高产量和开拓新产品是关键,为了达到这个目的,经常需要用分子生物学手段对底盘细胞进行改造。作为“下一代工业生物技术”的底盘细胞,嗜盐微生物也需要进行分子生物学改造。但是不同于遗传背景清晰和分子操作工具完善的模式微生物,嗜盐微生物作为非模式微生物,缺乏足够的分子改造方法。为了解决这一弊端,首先需要开发出适用于嗜盐微生物的分子操作工具。
2.1 基因组编辑工具CRISPR-Cas
CRISPR(规律成簇间隔短回文重复序列)是原核生物基因组内的一段重复序列,CRISPR 与Cas 蛋白组成的CRISPR-Cas 系统作为一种在古生菌中存在的天然保护机制,可以抵御外源核酸侵入古生菌基因组[11]。在古嗜盐菌Haloferax volcanii 中存在着CRISPR-Cas Ⅰ-B 系统[12-13],将该系统的cas3基因敲除后,DNA双链将无法被剪切,此时导入靶向特定DNA 序列的crRNA,Cascade 蛋白被crRNA 引导到该DNA 片段所在的位置,由于Cascade 蛋白形成了空间位阻,导致RNA聚合酶无法正常结合,因此目标基因的转录会受到抑制,通过这样的方法可以抑制基因表达,将crRNA 靶向目标基因的启动子或者模板链导致基因转录水平下降到原来的8%,将crRNA 靶向目标基因的编码链导致基因转录水平下降到原来的88%[14]。
除了调控基因表达,嗜盐微生物的基因组编辑技术同样至关重要。在嗜盐微生物中建立CRISPR-Cas9系统可以实现对嗜盐细菌的基因组编辑:在盐单胞菌Halomonas bluephagenesis TD01 和Halomonas campaniensis LS21 中,用两个质粒分别表达酿脓链球菌的Cas9 蛋白和靶向特定基因组DNA 序列的向导RNA(gRNA),在gRNA 的引导下,Cas9 蛋白在靶向位点切割DNA 双链,质粒上的同源臂和切割位点附近的相同同源臂发生同源重组,两段同源臂中间的序列从基因组上缺失,实现基因敲除[图1(a)];如果在质粒上的两段同源臂之间加入外源基因,则可以在同源重组过程中将外源基因插入基因组或替换基因组上的DNA片段[图1(b)][15]。
2.2 组成型启动子库PPorin lib
为了在嗜盐微生物中生产某种产物或进行其他改造,经常需要在嗜盐微生物中用组成型启动子表达外源基因,由于同一启动子在不同物种之间的表达情况存在差异,所以需要开发在嗜盐微生物中适用的启动子。孔蛋白Porin 是在盐单胞菌H.bluephagenesis 中表达强度最高的蛋白,该蛋白的启动子PPorin也被开发为在盐单胞菌H.bluephagenesis 中持续表达外源基因的组成型启动子[16]。PPorin的表达强度非常强,适合表达需要超高表达强度的基因,而将PPorin-10 区前的3 个碱基或者-10 区后、-35 区前的4 个碱基进行随机突变可以得到组成型启动子库PPorinlib[图1(c)],该启动子库包含95 个不同强度的启动子,最高表达强度是最低表达强度的3500 倍,可以用各种不同的强度表达基因,该启动子库不但可以在盐单胞菌中使用,在模式生物大肠杆菌中的表达范围也很广[17]。用该启动子库在盐单胞菌H.bluephagenesis 中调控4-羟基丁酰辅酶A 转移酶基因orfZ、4-羟基丁酰辅酶A脱氢酶基因4hbd、琥珀酸半醛脱氢酶基因sucD 和α-酮戊二酸脱羧酶基因ogdA 的表达,可以有效提高3-羟基丁酸(3HB)和4-羟基丁酸(4HB)的共聚物P34HB 的产量和4HB 在共聚物中的含量[17-18]。
2.3 诱导型表达系统
在对嗜盐微生物进行改造的过程中,除了组成型表达外源基因,还需要在特定条件下诱导外源基因的表达,这就要用到诱导型启动子。
嗜盐古菌Haloferax volcanii 的色氨酸酶启动子p.tna 受色氨酸调控[19-20],在100μg/mL 色氨酸的诱导下,p.tna 启动子控制的基因的表达水平提高了100 倍[19],用色氨酸诱导乙醇脱氢酶基因的表达可以将乙醇脱氢酶ADH/D1 的产量提高到16.8mg/g CDW(细胞干重)[21]。
T7 诱导系统是在大肠杆菌中广泛使用的诱导系统,然而,T7 RNA 聚合酶无法在盐单胞菌中正常表达,导致该诱导系统在盐单胞菌中失效[22]。为了在盐单胞菌中建立有效的诱导系统,研究者用异丙基-β-硫代半乳糖苷(IPTG)诱导类T7 RNA 聚合酶的表达,建立了类T7启动子受IPTG调控的诱导系统,用类T7 启动子表达外源基因,在一定范围内,IPTG 的浓度越高,外源基因的表达强度越强[22][图1(d)]。该系统已经用来在H. bluephagenesis中控制细胞形态[22-23]和生产PHA[18,24-25]。
3 嗜盐微生物作为生物合成的底盘细胞
作为下一代工业生物技术的底盘细胞,嗜盐微生物具备高效、大量合成多种产品的能力,这些产品包括多种聚羟基脂肪酸酯(PHA)、蛋白质和小分子化合物(表2)。

表2 嗜盐微生物合成多种产品
3.1 PHA的合成

图2 聚羟基脂肪酸脂(PHA)结构式
聚羟基脂肪酸酯简称PHA,是由100~30000个羟基脂肪酸聚合而成的一类大分子生物聚酯(图2),PHA多由微生物天然合成,在胞内以碳源和能源贮藏物质的形式存在。PHA具有类似合成塑料的物理化学特性以及合成塑料所不具备的生物可降解性和生物相容性,因此,PHA在可降解塑料、组织工程材料、缓释材料等方面有广阔的应用前景[60-62]。根据组成聚合物的单体含碳原子数目的差异,PHA可以分为三类[62]:第一类是单体含3~5 个碳原子的短 链PHA (scl PHA), 如 聚3- 羟 基 丙 酸 酯(P3HP)、聚3-羟基丁酸酯(PHB)、聚4-羟基丁酸酯(P4HB)、聚3-羟基戊酸酯(P3HV)及其共聚物聚3-羟基丁酸-4-羟基丁酸酯[P(3HB-co-4HB)](P34HB)、聚3-羟基丁酸-3-羟基戊酸酯[P(3HBco-3HV)](PHBV)等;第二类是单体含6个碳原子以上的中长链PHA(mcl PHA),如聚3-羟基己酸酯(PHHx)、聚3-羟基辛酸酯(PHO)及其共聚物聚3-羟基己酸-3-羟基辛酸酯[P(3HB-co-3HO)]等;第三类是短链PHA 和中长链PHA 的共聚物(sclco-mcl PHA),如聚3-羟基丁酸-3-羟基癸酸酯[P(3HB-co-3HD)]、聚3-羟基丁酸-3-羟基十二烷酸酯[P(3HB-co-3HDD)]等。
很多嗜盐微生物可以天然合成PHA,这为以嗜盐微生物为底盘细胞进行的PHA 工业生产提供了优势。Halomonas sp. YLGW01 可以用果糖生产PHB,产量达到细胞干重的94.6%,与以葡萄糖为碳源相比,以果糖为碳源生长的Halomonas sp.YLGW01细胞更大、更长,细胞长度从葡萄糖培养的2.34μm 提高到果糖培养的8.39μm[34]。Haloferax mediterranei 可以以淀粉为碳源生产PHBV,产量达到细胞干重的50.8%,3HV 摩尔分数为10.4%[41]。H.mediterranei可以生产PHBV的随机共聚物或者高阶共聚物,通过改变葡萄糖和戊酸盐的比例,可以调控3HV 的摩尔分数[42]。与普通微生物合成的PHBV和PHB以及医用级材料PLA相比,这种嗜盐微生物合成的PHBV有更好的生物相容性和生物可降解性[63],敲除该嗜盐菌表多糖合成相关的基因簇还可以将PHBV 的产量提高约20%[64-65]。Halomonas boliviensis 可以在含有0~25% NaCl、pH=6~11 的环境中生长[29],并利用葡萄糖、淀粉水解物、蔗糖、木糖或农业残渣等多种碳源合成PHB[28-30],其细胞干重可达到44g/L,PHB 含量为81%,PHB 生产率为1.1g/(L·h)[33]。Vibrio proteolyticu 可 以 在 含 有5%NaCl 的培养基中积累PHB,其最高PHA 含量为54.67%,细胞干重为4.948g/L[26]。
工程改造以后,下一代工业生物技术的重要底盘细胞H.bluephagenesis 和H.campaniensis 具备大量积累多种PHA的能力。在56h半连续发酵中,野生型菌株H.bluephagenesis TD01的细胞干重可以达到80g/L,PHB 含量占细胞干重的80%[31]。重组型盐单胞菌Halomonas campaniensis LS21 可以在添加了厨余垃圾等废料的海水中进行65d的开放式、非灭菌、连续发酵,积累的PHB 达到细胞干重的70%[32]。用分子生物学手段改造H.bluephagenesis和H. campaniensis 可以有效提高其合成的PHA 的种类、数量或者改变PHA 的积累方式。研究发现,H.bluephagenesis 的乙酰乙酰辅酶A 还原酶PhaB 是NADH依赖型的,因此,加入乙酸钠、葡萄糖酸钠等物质可以提高胞内NADH/NAD+比例,进而为PhaB 蛋白提供更多的能量,提高PHA 产量[66];在H.bluephagenesis 内表达orfZ、4hbd、sucD 和ogA 基因可以生产P34HB,精确调控上述基因的表达强度可以有效提高P34HB 的产量和聚合物中4HB 的摩尔含量[35-36],在7L发酵罐中改造过的H.bluephagenesis细胞干重达48g/L,P34HB产量为36g/L,占细胞干重的75%,4HB的摩尔含量可以达到16%[18];敲除2-柠檬酸甲酯合成酶基因prpC 和琥珀酸脱氢酶组装因子2基因sdhE、过表达甲基丙二酰辅酶A变位酶基因scpA、甲基丙二酰辅酶A 脱羧酶基因scpB和磷酸烯醇丙酮酸羧化酶基因ppc 的重组型H.bluephagenesis 可以生产PHBV,摇瓶中的细胞干重为6.3g/L,PHBV 含量为细胞干重的65%,3HV摩尔含量最高可达25%[24];抑制细胞分裂环蛋白基因ftsZ 和细胞骨架蛋白基因mreB 的表达可以增大H. campaniensis LS21 的体积,为PHA 的积累提供了更多的空间[67];敲除包涵体蛋白基因phaP 后,H.bluephagenesis 的PHA 颗粒会从多个小颗粒变成一个大颗粒[23]。
3.2 蛋白质的合成
除了合成PHA,嗜盐微生物还可以合成多种蛋白质,例如各种酶和PHA 相关蛋白,嗜盐微生物合成的酶不但可以在高盐环境下行使催化功能,而且在高温下有更强的稳定性[68]。
Halorubrum lacusprofundi 可以合成β-半乳糖苷酶,用冷休克蛋白基因cspD2的启动子过表达β-半乳糖苷酶基因,可以将产量提高20 倍[44]。Haloarcula marismortui 合成的乙醇脱氢酶ADH12可以在pH=10、60℃的环境中发挥催化功能[45]。Halobacterium salinarum 表达的谷氨酸脱氢酶最高可 以 在pH=10 的 环 境 下 发 挥 活 性[46]。 在Staphylococcus equorum sp. AMC7 和表面活性剂Triton X-100的刺激下,Halomonas sp.LM1C脂肪酶的产量会提高9倍,最高达3000U/L[47]。Haloferax sp.HA10 产生的淀粉酶可以在pH=5~9、NaCl 浓度为0.5~3mol/L 环境中催化淀粉水解,在55℃的环境中,该淀粉酶活性保持在99%[48]。H.bluephagenesis可以生产PHA 包涵体蛋白PhaP 和PHA 调控蛋白PhaR,PhaP 蛋白具有调控PHA 颗粒的形成的功能,在H.bluephagenesis 中的最高产量为1.86g/L[49],PhaR蛋白对脂类分子有乳化作用,H.bluephagenesis生产的PhaR 耐高温,经过98℃、105℃和115℃处理20min后,PhaR蛋白仍然可以发挥活性[50]。
3.3 小分子化合物的合成
嗜盐微生物还可以合成控制小分子化合物合成的酶,进而合成小分子化合物。
四氢嘧啶对维持微生物渗透压平衡起着重要作用,嗜盐微生物生长在高渗透压的环境中,所以会合成大量的四氢嘧啶,四氢嘧啶的合成主要依靠L-2,4-二氨基丁酸二酸盐乙酰转移酶基因ectA、L-2,4-二氨基丁酸二酸盐氨基转移酶基因ectB和四氢嘧啶合成酶基因ectC。在H.bluephagenesis中进行精确的代谢流调控,调整ectABC 和其他上游基因的表达强度,可以有效提高四氢嘧啶的产量,在7L发酵罐中得到28g/L四氢嘧啶或者联产8g/L四氢嘧啶和24g/L PHB,其中PHB占细胞干重的75%[51]。在Halomonas elongate DSM 2581T中,四氢嘧啶转运蛋白TeaABC 将四氢嘧啶从胞内转移到胞外[52],用含15%和3% NaCl 的培养基交替培养H. elongate DSM 142T,重复9次,可以得到155mg/g CDW四氢嘧啶[53]。Halomonas boliviensis 两次补料发酵合成四氢嘧啶的速度为每天11.1g/L,通过改变渗透压分泌到胞外的四氢嘧啶产量为每天9.1g/L[54]。
除了四氢嘧啶,甜菜碱也在平衡渗透压中起着重 要 作 用 , Actinopolyspora halophila 和Ectothiorhodospira halochloris可以以甘氨酸为底物合成甜菜碱[55]。嗜盐微生物还可以合成萜类化合物,表达IPP异构酶基因idi、牻牛儿基牻牛儿基焦磷酸(GGPP)合成酶基因crtE、八氢番茄红素合成酶基因crtB、八氢番茄红素脱氢酶基因crtI 和番茄红素环化酶基因crtY 后,H. elongate 可以在2%NaCl 中生 产560mg/g CDW β - 胡 萝 卜 素[56], 在H.mediterranei 中表达crtB 和crtI 基因可以生产番茄红素,敲除番茄红素下游代谢途径和PHBV合成途径后,番茄红素的产量可以提高到119.25mg/g CDW[57]。此外,重组型H. bluephagenesis 还可以生产33g/L 的苏氨酸[58]或0.7g/L 的5-氨基乙酰丙酸(ALA)[59]。
4 高密度下嗜盐微生物的应用前景
高密度(细胞干重>50g/L)培养是工业生物技术的难题之一,在高密度环境下,细胞的代谢更为复杂,调控更加困难,以嗜盐菌为底盘细胞的下一代工业生物技术在高密度培养方面有应用前景。嗜盐微生物H. bluephagenesis TD01 或者其衍生菌株可以进行的开放式、非灭菌的连续发酵,已经完成了7L实验室小试、1000L和5000L工业生物反应器中试,在5000L 中试中,细胞干重达到100g/L,P[3HB-co-13.5% (摩尔分数) 4HB]含量达到60.4%[31,69]。但是,高密度下的细胞代谢更为复杂、基因诱导表达更为困难、耗能更大、稳定性更低,因此,研究并解决“高密度壁垒”对下一代工业生物技术的完善和发展有着重要的意义。
4.1 高密度下多代谢模块高效调控
在嗜盐微生物发酵培养时,在不同生长时期进行取样,对每个取样点进行转录组和蛋白组检测,可以获得不同生长时期细胞RNA 和蛋白质的表达情况,以及RNA 和蛋白质在不同生长时期的动态变化过程,根据这些信息可以解析高密度下嗜盐菌的代谢调控网络,结合生物量、生长趋势、膜组分、ATP需求量等关键参数,可以建立这些参数与基因转录和蛋白质表达在不同生长时期的关系网络,根据这些信息解析嗜盐微生物在高密度时期的代谢特点,而后对所表达的代谢通路进行适宜的表达调控,使得代谢调控将更加精确高效[70-71]。
4.2 高密度可诱导强启动子的开发与细胞体积调控
根据不同生长时期的转录组和蛋白组测序结果,还可以分析出嗜盐微生物各基因表达强度随生长情况变化的规律,从而找到在低密度情况下表达水平低,而在高密度情况下表达水平高的启动子,这样的启动子可以被高密度条件诱导。用这样的高密度可诱导启动子调控细胞分裂环蛋白基因ftsZ或者细胞骨架蛋白基因mreB 的表达,在高密度条件下实现对细胞体积的调控,使细胞响应高密度信号,体积变大,这有利于PHA 的积累和发酵结束后将细胞从发酵液中分离、进行下游工业环节[67]。
4.3 高密度下节能和高转化率
在高密度条件下细胞能获取的氧气会减少,能量供给随之降低,为了解决这一问题,可以通过敲除嗜盐微生物细胞外膜、在嗜盐微生物胞内或者细胞膜上表达透明颤菌血红蛋白VHb 等方式增加细胞获取氧气的能力[72],还可以在高密度条件下抑制三羧酸循环、引入外源的糖酵解途径,使得代谢流通往无氧呼吸途径,消耗NADH,为细胞提供能量,提高能量转化效率,保持嗜盐微生物在高密度条件下合成产物的能力。
4.4 高密度下酶的稳定性调控
为了降低各产物合成酶在高密度下的降解,提高高密度下产物的合成能力,可以通过定向进化等方式,对相关合成酶进行改造,提高合成酶的稳定性,或者突变蛋白酶,抑制其降解PHA 合成酶的能力,最终提高高密度下嗜盐微生物合成PHA 等产物的能力[73]。
5 结语
由于嗜盐微生物的发酵节水、节能、无需灭菌、过程连续,以嗜盐微生物为底盘细胞的下一代工业生物技术有广阔的发展前景(图3)。“下一代工业生物技术”将以嗜盐微生物等极端微生物为底盘细胞,利用海水、普通碳源、秸秆、厨余垃圾等特殊碳源进行无灭菌、开放式和连续发酵,在发酵的中后期,诱导嗜盐微生物发生形变以便于菌体的分离,沉降的菌体可以用来提取PHA 等产物,发酵液可以回收重复使用,或者还可以用一株嗜盐菌同时生产多种产物,从发酵液和菌体中同时获得产品。此外,结合新兴的人工智能(AI)技术,“下一代工业生物技术”还将打造“无人车间”,自动控制上述生产过程,最终实现PHA、蛋白质、小分子化合物等各类产物的简易、高效、低价生产。
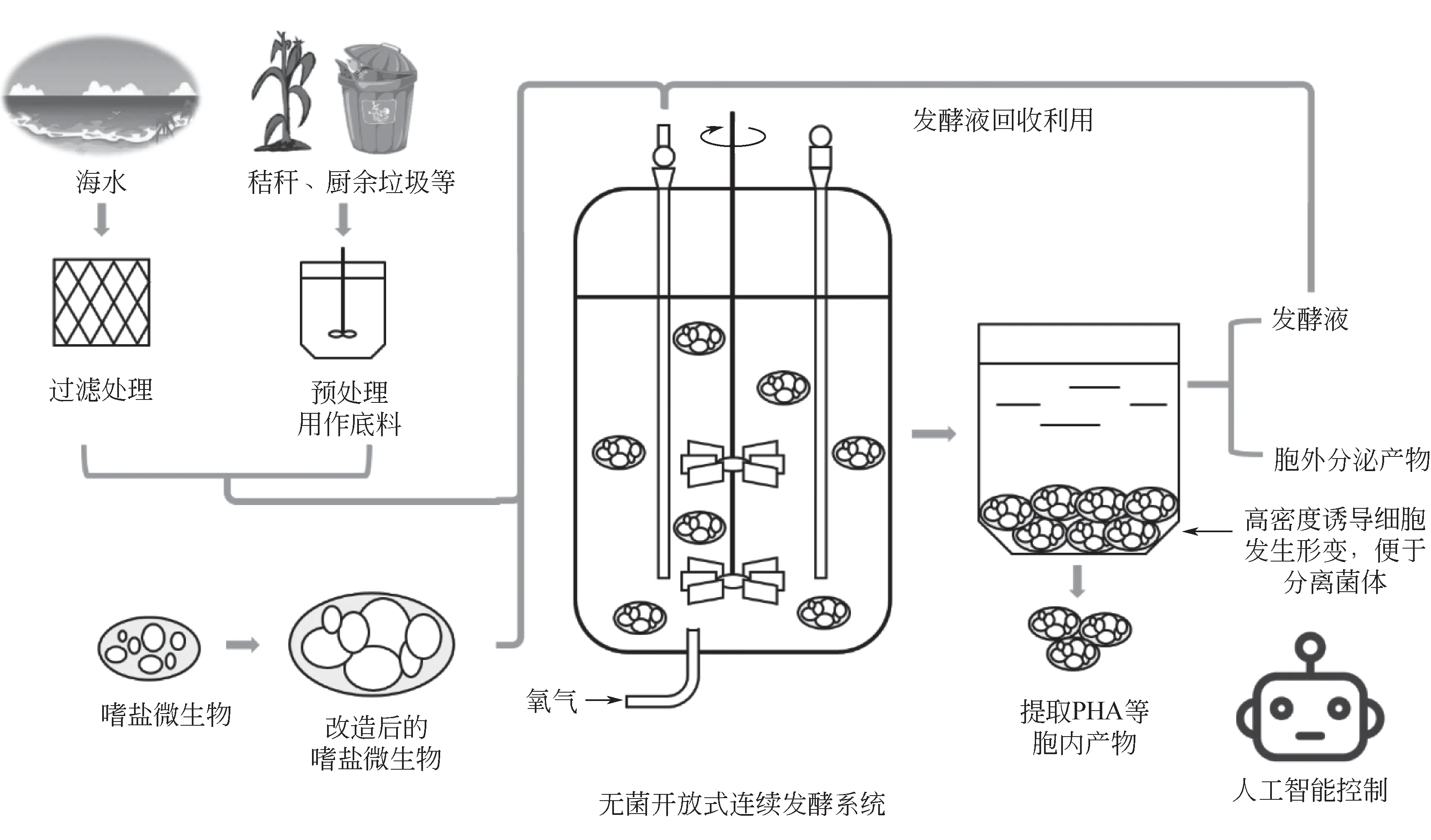
图3 以嗜盐微生物为底盘细胞的下一代工业生物技术构想
