茶足柄瘤蚜茧蜂碳水化合物代谢相关的滞育关联基因差异表达分析
2021-03-30刘爱萍韩海斌陈国泽黄海广
刘 敏,刘爱萍*,韩海斌,甘 霖,陈国泽,黄海广
(1. 中国农业科学院草原研究所,呼和浩特 010010;2. 阿拉善科技信息研究所,内蒙古阿拉善左旗,750399;3. 阿拉善右旗曼德拉苏木农牧业综合站,内蒙古阿拉善右旗,750300;4. 内蒙古自治区林业科学研究院,呼和浩特 010010)
近几年内蒙古地区的田间调查发现,茶足柄瘤蚜茧蜂LysiphlebustestaceipesCresson是苜蓿蚜AphiscraccivoraKoch的优势寄生性天敌,对控制苜蓿蚜有重要作用(郑永善和唐保善, 1989; 刘爱萍等, 2012)。滞育并不是简单的发育停止,而是由环境与遗传因素决定的、程序化启动的极其缓慢的发育过程。昆虫在滞育期间,体内会发生一系列生理生化变化,如脂类、碳水化合物和氨基酸等内源营养物质特异性地积累或转化,以确保滞育个体在逆境中存活及滞育解除后发育的能量需求(任小云等, 2016)。对茶足柄瘤蚜茧蜂等天敌昆虫来说,阐明滞育的分子调控机理,有可能通过遗传信息调控实现昆虫滞育的人工控制,从而延长天敌昆虫的寿命,保证天敌昆虫产品的货架期,最终发挥生物防治的最佳效果。
糖类作为昆虫生命活动过程的中间代谢产物和重要的能源物质,与昆虫滞育有密切的关系(刘流等, 2010),葡萄糖是大部分生命体的主要供能分子,在新陈代谢中起着重要作用。当机体能量充足时,葡萄糖可以以淀粉或糖原等聚合物形式进行储备;对能量需求增加时,葡萄糖又可以迅速从细胞内贮存的聚合物中释放,通过有氧或者无氧呼吸提供能量。葡萄糖不仅是供能分子,同时也可以提供氨基酸、核苷酸、脂肪等生物合成反应的代谢中间物质。
昆虫滞育期间变化明显的糖类物质主要有糖原、海藻糖。在昆虫体内,海藻糖(trehalose)主要是由脂肪体合成并分泌到血淋巴,被其他的组织吸收后被利用(Bouniasetal., 1993),是昆虫血糖的主要组成,海藻糖的含量与海藻糖酶(trehalase, Treh)活力的变化影响着昆虫的滞育、滞育解除和发育(仵均祥等, 2004)。糖原是昆虫滞育期间主要的能源物质,也是抗冻保护剂(糖、醇类)的主要来源之一。已知滞育昆虫的糖原代谢分为糖—醇积累型和海藻糖积累型,两类物质都在滞育昆虫的抗冻、蛋白质保护、降低渗透压或过冷却点等方面发挥着重要的作用。鞭角华扁叶蜂Chinolydaflagellicornis在滞育期间,血淋巴中的糖原逐渐减少,海藻糖含量逐渐增加,在滞育前期,海藻糖与糖原相互转化,同时发现滞育阶段及温度与这种转化关系有关(任小云等, 2016)。斑蛾Zygaentrlfolii进入滞育后糖原是最重要的能源物质,在滞育个体中糖原含量是非滞育个体的2倍多(任小云等, 2016)。有研究表明,糖原是棉铃虫Helicoverpaarmigera滞育蛹生命活动的主要能量来源,随着滞育强度的深入,糖原含量逐渐减少(李玉艳, 2011)。
过去人们认为滞育是由滞育激素导致的,但越来越多的研究证明,海藻糖在两者之间发挥重要作用。在对家蚕Bombyxmori的研究中发现,进入滞育前,滞育激素通过调控海藻糖酶的活性,促使海藻糖被分解成葡萄糖进入卵细胞,葡萄糖再转化为糖原诱导卵滞育(Okitsugu, 1996)。海藻糖酶由海藻糖酶基因编码调控,干扰海藻糖酶基因会影响昆虫的生长发育,表明Treh在糖代谢通路中发挥重要作用,参与昆虫重要的生理过程(Chenetal., 2010; 张倩等, 2012; Zhaoetal., 2016)。
目前对于茶足柄瘤蚜茧蜂的滞育研究中,涉及分子机理的研究较少,究竟在茶足柄瘤蚜茧蜂蛹中,碳水化合物代谢与滞育过程有怎样的关系尚不清楚。本试验对茶足柄瘤蚜茧蜂滞育蛹与非滞育蛹进行了转录组测序,重点分析了与糖代谢相关的差异表达基因,结合糖酵解,淀粉与蔗糖代谢,柠檬酸循环及茶足柄瘤蚜茧蜂具体的生理学和环境调控在转录组水平的研究等,从整体上和细节上为茶足柄瘤蚜茧蜂滞育相关的环境调控、物质能量的代谢机理提供参考。
1 材料与方法
1.1 供试虫源、供试寄主植物与滞育诱导
茶足柄瘤蚜茧蜂、寄主蚜虫苜蓿蚜采自中国农业科学院草原研究所沙尔沁基地,供试寄主植物为蚕豆Viciafaba。
苜蓿蚜采自基地的羊柴Hedysarummongolicum植株上,并转接在室内的水培蚕豆苗上繁殖,接虫后对蚕豆苗进行笼罩(100目防虫网笼,55 cm×55 cm×55 cm),确保苜蓿蚜未被天敌寄生,试验用2~3龄的苜蓿蚜若蚜作为寄主,在温室内饲养5代以上作为供试虫源。
从基地采集被寄生的苜蓿蚜僵蚜,从中挑取未羽化破壳的僵蚜置于人工气候箱(上海一恒公司MGC-HP系列)温度25±1℃,相对湿度70%±5%,光周期L ∶D=14 h ∶10 h条件下培养,待蜂羽化后,挑选茶足柄瘤蚜茧蜂转移至试管(10 cm×3 cm)内,用20%蜂蜜水作为补充营养,接入具有苜蓿蚜的蚕豆苗上,建立茶足柄瘤蚜茧蜂种群作为供试虫源,并在室温下用苜蓿蚜有效扩繁10代以上。
在室温下养虫笼中将刚羽化成蜂按1 ∶100的蜂蚜比释放成对茶足柄瘤蚜茧蜂,根据实验室前期研究基础可知,苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,寄生蜂卵继续发育120 h,此时僵蚜体内寄生蜂处于高龄幼虫(3~4龄)阶段,高龄幼虫为茶足柄瘤蚜茧蜂感受滞育信号的敏感虫态,将此时的僵蚜放入人工气候箱中进行滞育诱导。高龄幼虫处于滞育环境条件时,并不会立刻停止发育,而是继续发育一段时间,经试验验证,当发育至蛹时,便不再继续发育(孙程鹏, 2018)。本实验中,诱导茶足柄瘤蚜茧蜂滞育的温光组合为,温度8℃、光周期L ∶D=8 h ∶16 h,诱导时长为30 d。选取经过30 d滞育诱导的僵蚜进行解剖,选择虫态是蛹的茶足柄瘤蚜茧蜂进行收集,以获得滞育组样品;苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,放置在25±0.5℃、RH70%±5%、光周期L ∶D=14 h ∶10 h、光照强度8 800 Lx(人工气候箱,上海一恒公司MGC-HP系列)条件下,寄生蜂卵继续发育168 h(此时蚜茧蜂处于蛹态),对僵蚜进行解剖,挑选饱满,有活力的蛹作为正常发育组样品。
1.2 测序和组装
茶足柄瘤蚜茧蜂正常发育组、滞育组转录组的测序工作由北京诺禾致源科技股份有限公司完成。测序进行3个生物学重复,每个重复需100头,共构建6个文库。测序完成后,经数据预处理,采用Trinity(Grabherr, 2011)对clean reads进行拼接。
1.3 序列筛选
1)拼接得到的茶足柄瘤蚜茧蜂转录组测序结果与Nr数据库进行Blastx比对(将核酸序列翻译为蛋白,再进行比对),筛选条件E-value<1e-5,得到注释基因。
2)应用每个样本的RPKM(reads per kilo base of exon model per million mapped reads),以每个样本的总表达量为内标进行Fisher-test差异检验,将正常发育组和滞育组转录组结果进行比较,采用FDR与表达差异倍数Fold-Change方法进行差异基因挑选,筛选条件为:FDR≤0.05,Fold-change≥2。
3)筛选的差异基因中,与正常发育组和滞育组相比,被注释成功的差异基因定义为滞育关联基因,并提取序列。
2 结果与分析
2.1 测序及组装
正常发育蛹与滞育蛹经Illumina HiSeq平台测序,将Trinity拼接得到的转录本序列,作为后续分析的参考序列。以Corset层次聚类后得到最长Cluster序列进行后续的分析。对转录本及聚类序列长度分别进行统计,结果见表1和图1。

图1 Nr比对上的物种分布图Fig.1 Homology search against Nr database for Lysiphlebus testaceipes transcriptome unigenes

表1 拼接长度分布情况一览表
2.2 Unigenes在其他物种中的分布情况
通过Nr库比对注释的结果,可以统计并绘制比对上的物种分布图。因为目前还没有报道茶足柄瘤蚜茧蜂的基因组序列,所以利用其他物种的已知基因组对实验中的unigene序列进行了比对。在种间分布上,络新妇蛛Nephilaclavipes(16.0%)、丽蝇蛹集金小蜂Nasoniavitripennis(9.1%)、多胚跳小蜂Copidosomafloridanum(8.0%)、胡蜂Diachasmaalloeum(6.3%)、Trichomalopsissarcophag(6.0%)和其他种others(图1)。
2.3 差异基因GO富集分析
在非滞育组与滞育组茶足柄瘤蚜茧蜂差异表达的基因中,GO注释到25 666个差异基因(图3),总共分为生物过程(BP),细胞成分(CC),分子功能(MF)三部分。差异基因的GO功能富集主要集中于代谢过程(metabolic process),包括脂代谢、氨基酸代谢、碳水化合物代谢等,信号转导(signal transduction),结合(binding)功能、催化活性(catalytic activity)。其中GO注释到碳水化合物代谢(carbohydrate metabolic process)条目的差异基因数为1 050。

图2 差异基因GO富集条目图Fig.2 GO enrichment map of differentially expressed genes for Lysiphlebus testaceipes

图3 茶足柄瘤蚜茧蜂滞育蛹与非滞育蛹差异表达基因KEGG富集气泡图Fig.3 Scatter plot of KEGG enrichment of differentially expressed genes in diapause and non-diapause pupae of Lysiphlebus testaceipes注: 图中横坐标轴为相应通路中差异基因的数目与鉴定出的总基因数目的比值,值越大,说明在该通路中差异基因富集程度越高。点的颜色代表超几何检验的P值,值越小,说明检验的可靠性越大、越具统计学意义。点的大小代表相应通路中差异基因的数目,值越大,该通路内差异基因越多。Note: The abscissa in the figure is the ratio of the number of differentially expressed genes in the corresponding pathway to the number of total proteins identified. The larger the value is, the higher the concentration of genes in this pathway is. The color of points represents the P-value of hypergeometric test. The smaller the value is, the more reliable and statistically significant the test is. The size of the dots represents the number of genes in the corresponding pathway. The larger the dot, the more genes in the pathway.
2.4 差异基因KEGG富集分析
将茶足柄瘤蚜茧蜂非滞育组与滞育组差异表达基因序列进行KEGG在线分析,通过KEGG pathway数据库分析,共分为新陈代谢、遗传信息加工、环境信息处理、细胞过程和有机体系统五大类。7 944个差异表达基因共映射到228个通路,分析发现这些基因主要集中在碳水化合物代谢、脂质代谢、信号转导等途径中。
对茶足柄瘤蚜茧蜂滞育蛹与非滞育蛹进行转录组测序,KEGG注释到的与糖代谢相关的差异表达基因共149个,此次重点分析糖酵解/糖异生(glycolysis/gluconeogenesis),淀粉与蔗糖代谢(starch and sucrose metabolism),柠檬酸循环三条途径(citrate cycle),差异表达基因分别为18,10和18个。图3-图5展示了三条代谢途径的KEGG富集通路图,明确代谢路径。在表2中,糖酵解/糖异生(glycolysis/ Gluconeogenesis)途径中,磷酸果糖激酶(phosphofructo kinase, PFK)基因、磷酸甘油酸激酶(phosphoglycerate kinase, PGK)基因、醛缩酶(aldolase, ALDO)基因上调表达,甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH)基因、磷酸甘油酸变位酶(phosphoglycerate mutase, PGAM)基因、磷酸烯醇式丙酮酸羧激酶(Phosphoenolpyruvate carboxykinase, PEPCK)基因下调表达。PFK和PGK是糖酵解途径中的关键酶,基因上调表达,两种酶含量增加,导致的结果是糖酵解途径活跃表达。GAPDH和PGAM是糖酵解和糖异生途径中共有的酶,同时PEPCK是糖异生途径中的关键酶。
在茶足柄瘤蚜茧蜂淀粉和蔗糖代谢(Starch and sucrose)途径中,与非滞育组相比,滞育组糖原合酶(glycogen synthase, GYS)基因、海藻糖合酶(Trehalose 6-phosphate synthase, TreS)基因上调表达,海藻糖酶(trehalase, TreH)基因下调表达。
在柠檬酸循环(citric acid cycle)途径中,苹果酸脱氢酶(malate dehydrogenase, MDH)基因上调表达,异柠檬酸脱氢酶(isocitrate dehydrogenase, IDH)基因下调表达。MDH基因上调表达,MDH增加,推测其与滞育状态下的生理需求相关。IDH是柠檬酸循环中重要的限速酶,IDH基因下调表达,IDH合成减少,导致柠檬酸循环反应速率降低。
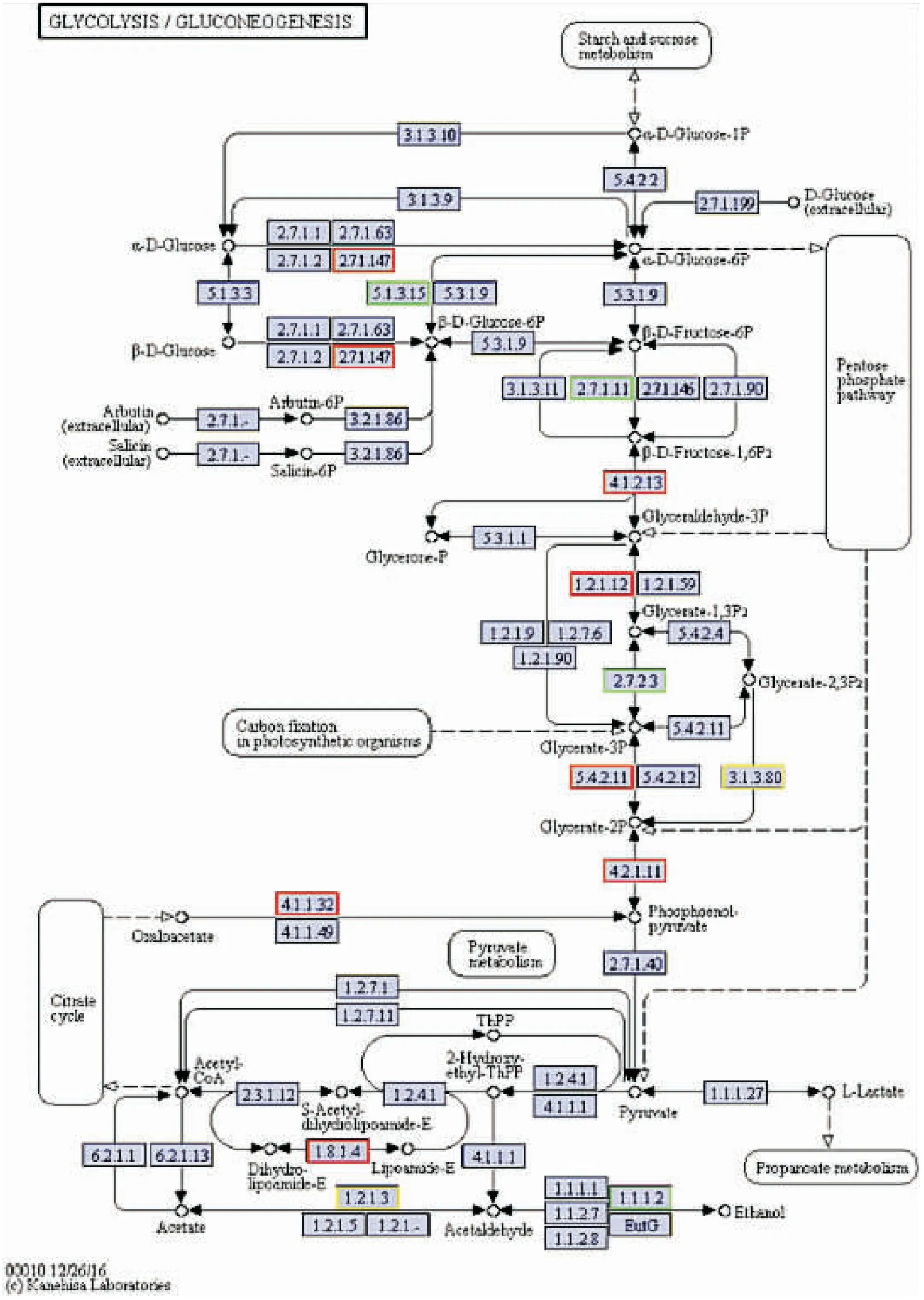
图4 差异基因在糖酵解/糖异生途径的富集通路图Fig.4 Pathway map of differentially expressed genes enrichment in glycolysis/ gluconeogenesis
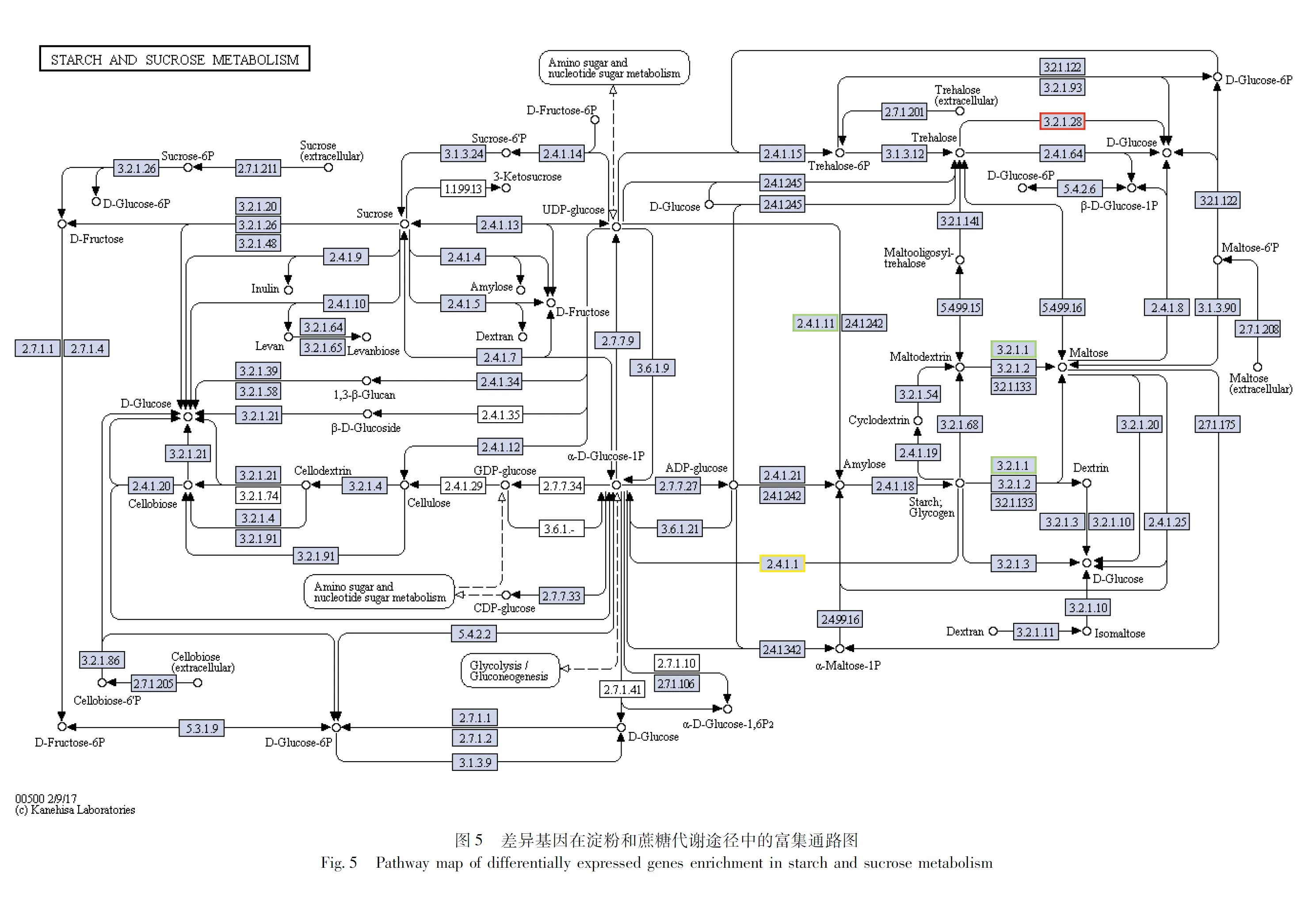

图6 差异基因在柠檬酸循环途径中的富集通路图Fig.6 Pathway map of differentially expressed genes enrichment in citrate cycle注:包含上调基因的KO节点边框标红色,包含下调基因的KO节点边框标绿色,包含上下调的边框标黄色。实心箭头代表分子间的相互作用或关系,圆圈代表化学分子。方框-实心箭头-圆圈相连,代表表达;方框-实心箭头-竖线-圆圈相连,代表抑制;方框-虚线箭头-圆圈相连,代表间接作用;直线-方框-实心箭头-圆圈-直线-方框-实心箭头相连,代表两个连续的反应步骤。带箭头的直线上标有“+P”代表磷酸化,标有“-P”代表去磷酸化。Note: The KO node border containing the up-regulated genes is red, containing the down-regulated genes is green, and the border containing up-and-down regulated genes is yellow. A solid arrow represents an interaction or relationship between molecules, and a circle represents a chemical molecule. Box-solid arrow-circle linked, representing expression; Box-solid arrow-vertical line-circle linked, representing inhibition; Box-dashed arrow-circle linked, representing indirect action; Straight-box-solid arrow-circle-straight line-box-solid arrow linked, representing two consecutive reaction steps; Lines with arrows are marked “p” for phosphorylation and “-p” for dephosphorylation.
3 结论与讨论
在本研究中,重点分析了与茶足柄瘤蚜茧蜂滞育过程中糖代谢相关的三条途径,分别是糖酵解/糖异生,淀粉和蔗糖代谢和柠檬酸循环。涉及到的基因主要有PFK,PGK,ALDO,GAPDH,PGAM,PEPCK,GYS,TreS,TreH,MDH,IDH等。
糖酵解途径是葡萄糖的主要代谢途径,为线粒体TCA循环提供中间代谢产物,从而为机体提供大部分生存所需的能量。PFK和PGK这两种酶是糖酵解途径中的关键酶,在滞育的茶足柄瘤蚜茧蜂蛹中,磷酸果糖激酶基因和磷酸甘油酸激酶基因较非滞育蛹上调表达,表明在茶足柄瘤蚜茧蜂滞育过程中糖酵解途径活跃进行,因此推测茶足柄瘤蚜茧蜂在滞育过程中依赖糖酵解途径转换能量,供生物体维持生命活动。
糖异生指的是以非糖物质作为前体合成葡萄糖的作用,GAPDH,PGAM,ALDO为糖酵解与糖异生途径共有的酶,磷酸甘油酸变位酶基因,甘油醛-3-磷酸脱氢酶基因和醛缩酶基因下调表达,可以认为是糖异生途径表达受到抑制或糖酵解途径不能顺利进行下去。PEPCK是糖异生途径中的关键酶,而磷酸烯醇式丙酮酸羧激酶基因在滞育过程中下调表达,因此猜测在茶足柄瘤蚜茧蜂蛹滞育过程中GAPDH基因、PGAM基因、PEPCK基因的下调表达共同抑制了糖异生途径的进行。
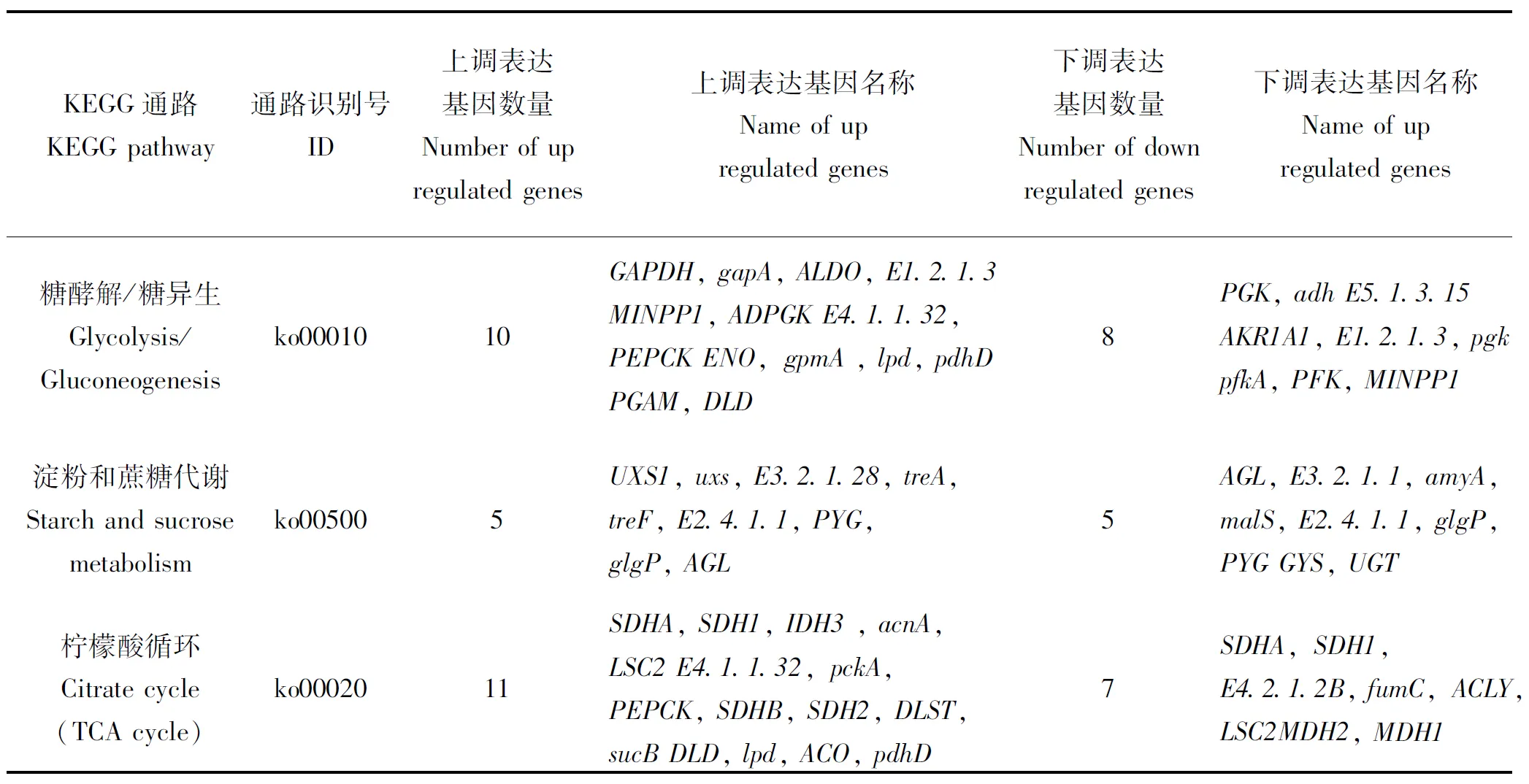
表2 糖代谢重要途径差异基因表达情况
海藻糖是昆虫的血糖,是昆虫的能量来源,在海藻糖酶的作用下,分解成为两分子葡萄糖,再进一步通过糖酵解为昆虫提供能量(Chenetal., 2010; Shuklaetal., 2015; Xiongetal., 2016; Zhaoetal., 2016; Yangetal., 2017)。另一方面,海藻糖在应对逆境胁迫时可作为应激代谢反应的主要产物发挥作用(Wharton, 2011; Shietal., 2016),是昆虫重要的储能物质,也是应激代谢产物之一。海藻糖酶能够分解海藻糖,通过基因的表达,影响酶活性,进而对昆虫蜕皮、变态、发育及繁殖等生命过程造成影响(唐斌等, 2012),海藻糖酶是滞育激素调控代谢过程中的关键酶(徐卫华, 1999)。在滞育的茶足柄瘤蚜茧蜂蛹中,海藻糖酶基因基因下调表达,海藻糖合酶基因上调表达,说明海藻糖分解少,合成多,导致海藻糖积累。糖原是主要的能量物质,其作用与脂肪类似(丁惠梅等, 2011)。滞育的茶足柄瘤蚜茧蜂蛹中,糖原合酶基因上调表达,糖原积累,糖原经水解转变为葡萄糖,进入糖酵解途径,糖酵解的产物经柠檬酸循环后释放出大量能量,供生物进行正常的生命活动,这说明糖原可能与脂肪一样作为储备能源物质,参与茶足柄瘤蚜茧蜂体内能量代谢。而海藻糖则可能作为保护剂与糖原相互转化,参与了茶足柄瘤蚜茧蜂的滞育调节。
在需氧生物中,柠檬酸循环是普遍存在的一种获取能量的代谢方式,是联系糖类、脂质、蛋白质三大主要营养物质的枢纽及最终代谢通路(王荫长, 2001)。研究发现,滞育过程中昆虫代谢水平显著下调,有些甚至代谢抑制达到90%(Ragland, 2009)。本试验结果显示,参与柠檬酸循环的异柠檬酸脱氢酶基因在滞育个体中下调表达,苹果酸脱氢酶基因上调表达。异柠檬酸脱氢酶作为催化异柠檬酸氧化形成α-酮戊二酸反应中的限速酶,此反应不可逆,是柠檬酸循环中重要的限速步骤。已有研究表明, 柠檬酸循环在昆虫滞育期间受到抑制(Michaud and Denlinger, 2007; Xuetal., 2012; Luetal., 2014)。在调节能量释放速率中起到关键作用,在茶足柄瘤蚜茧蜂滞育蛹中基因下调表达,也反应出能量供应的关键循环之一—柠檬酸循环整体反应速率的降低,表现在滞育茶足柄瘤蚜茧蜂体内维持了低能量代谢,与茶足柄瘤蚜茧蜂蛹在滞育条件下整体呈现的低水平代谢相符。
在研究中发现,甘油、山梨醇等醇类物质在低温环境中对苹果酸活性保持具有重要作用,而在滞育个体中,已知甘油、山梨醇、甘露醇等多元醇是不断积累增加的,它们可以降低生物体的过冷却点,保护细胞膜免受结冰损伤(Mansingh and Smallman, 1972; Zachariassen, 1985; Jaindl and Popp, 2006)。苹果酸脱氢酶为NAD-依赖型,定位于线粒体基质中,催化柠檬酸循环中 L-苹果酸羟基氧化形成羰基,生成NADH并与草酰乙酸相互转化,以此调节生物体内的物质能量代谢(Goward and Nicholls, 1994)。在茶足柄瘤蚜茧蜂滞育蛹中,苹果酸脱氢酶基因上调表达,苹果酸脱氢酶增加,在滞育过程中苹果酸脱氢酶的主要作用是参与NAD(烟酰胺腺嘌呤二核苷酸, nicotinamide adenine dinucleotide)的循环利用及物质循环(王启龙等, 2012),推测其与滞育状态下的生理需求相关,一方面与茶足柄瘤蚜茧蜂对NAD的合成和利用有关,另一方面或许是应对滞育环境条件的一种应激方式,与正常个体的代谢通路相比,滞育个体开启了另外的代谢通路以适应环境条件的改变。苹果酸脱氢酶和异柠檬酸脱氢酶作为糖代谢中重要的酶类,它们的含量变化与滞育期间昆虫的能量调节及代谢密切相关(Hayakawa and Chino, 1981, 1982; Hahn and Denlinger, 2011)。
本实验重点研究糖代谢在茶足柄瘤蚜茧蜂滞育中的作用,糖类物质在滞育过程中的变化对昆虫有着怎样的影响。从整体上和细节上为茶足柄瘤蚜茧蜂滞育相关的环境调控、物质能量的代谢机理提供参考。研究糖代谢对揭示滞育过程的分子机制至关重要,阐明分子机理有可能通过遗传信息调控实现人工控制昆虫滞育,丰富常规的温光互作滞育诱导技术,从而延长天敌昆虫的寿命,保证天敌昆虫产品的货架期,最终发挥生物防治的最佳效果。
