基于HS-SPME-GC-MS探究郫县豆瓣优势菌株地衣芽孢杆菌和埃切假丝酵母的酯合成途径及能力
2021-03-24杨林子卢云浩何强
杨林子,卢云浩,何强
(四川大学 轻工科学与工程学院,成都 610065)
郫县豆瓣是中国传统发酵调味品,迄今已有三百多年历史,以其酱香浓郁、味辣香醇、红棕油亮等特点,被誉为“川菜之魂”[1]。郫县豆瓣在自然条件下,由多种类型的微生物与原料经长达半年的协同发酵而成,其独特风味的形成主要来源于微生物在相对封闭的混合发酵环境中产生的各种呈香物质[2]。酯类物质是郫县豆瓣等发酵食品中常见的挥发性风味物质[3],尤其是乙酯类和乙酸酯类物质,其阈值较低,在低浓度时可以呈现出令人愉悦的香气,平衡产品整体风味;浓度较高时,则会使食品中缺少果香风味,影响产品质量[4]。发酵食品中的酯类物质主要通过微生物途径产生,化学合成途径反应时间长且缓慢[5],因此,探究微生物的酯合成途径和能力对于控制发酵食品整体风味具有重要意义。
本研究采用气相色谱-质谱联用技术,通过建立化
学反应体系探究郫县豆瓣中地衣芽孢杆菌和埃切假丝酵母的酯合成途径和能力,以期进一步加深对发酵过程中酯合成机制的认识,为加速和改善郫县豆瓣酯类风味物质的形成提供了理论依据。
1 材料与方法
1.1 实验菌株
地衣芽孢杆菌(NCBI登记号:MK063711)、埃切假丝酵母(NCBI登记号:MK063704):四川大学食品系实验室保藏菌株,分离自郫县豆瓣。
1.2 材料与试剂
营养肉汤、YPD液体培养基:1%酵母膏、2%葡萄糖、2%蛋白胨,购自北京奥博星生物技术有限责任公司。
己酸乙酯、乙酸己酯等标准品:购自Sigma公司;己醇、三乙酸甘油酯及其他化学试剂:均为分析纯,购自阿拉丁公司。
1.3 仪器与设备
SPME手动进样手柄、DVB/CAR/PDMS(50/30 μm)萃取头 美国Supelco公司;QP2010 GC-MS 日本岛津公司。
1.4 实验方法
1.4.1 胞外提取液的制备
地衣芽孢杆菌接种至营养肉汤,埃切假丝酵母接种至YPD液体培养基,分别于28 ℃和37 ℃恒温摇床培养至对数生长期末期,取活化2次后的菌液在10000 g、4 ℃的条件下离心10 min,取其上清液作为胞外提取液,保存于-18 ℃备用,采用Bradford法测定其蛋白含量[6]。
1.4.2 建立化学反应体系
酯合成反应体系的建立参照Liu等[7]的方法。各体系包括菌株胞外提取液、磷酸钠盐缓冲液以及反应底物,在酯化体系中,反应底物为100 mmol/L己酸(乙酸)和100 mmol/L乙醇(己醇);醇解体系为100 mmol/L己醇与33 mmol/L三乙酸甘油酯。反应体系pH均调整为5.5,接近郫县豆瓣发酵pH值。各体系建立后立即静置于30 ℃恒温水浴锅中进行反应,每隔20 min检测一次。
1.4.3 pH值对菌株酯合成活力的影响
酯合成反应体系的建立参照1.4.2,利用缓冲液调节各体系的pH值分别为4,5,6,在30 ℃条件下静置反应1 h后检测其产酯情况。
1.4.4 挥发性风味物质的提取
各体系反应结束后,立即放置于60 ℃水浴锅中,插入萃取头,采用顶空固相微萃取的方法吸附30 min后,将萃取头插入GC-MS进样口,250 ℃解吸3 min。
1.4.5 GC-MS分析
1.4.5.1 气相色谱条件
DB-5MS毛细管色谱柱(30 m×0.25 mm×0.25 μm),载气为氦气,流速为1.0 mL/min;不分流进样;升温程序:初始温度40 ℃,保持2 min,以5 ℃/min的速度升至60 ℃,再以8 ℃/min升至140 ℃,最后以12 ℃/min升至250 ℃保持8 min。
1.4.5.2 质谱条件
电子电离源,电子能量70 eV,离子源温度230 ℃,接口温度250 ℃,质量扫描范围30~500 m/z,溶剂延迟时间1 min。
1.4.5.3 挥发性物质的定性
主要采用谱库检索和保留指数两种方法对各体系中酯类和前体物质进行定性。谱库检索主要依据所检测物质的质谱信息与NIST 14谱库作比较,根据相似度进行取舍,一般大于80%时对该物质予以确认;保留指数鉴定法是以C8~C12的正构烷烃为标准,计算样品中挥发性物质的保留指数,与其他文献中相同物质的保留物质进行比较。
1.4.5.4 挥发性物质的定量
采用外标法进行定量(μg/mL),并定义1个酯合成酶活力单位(1 U)为1 h内体系合成1 nmol对应的酯,酯合成比活力为U/μg蛋白。
1.5 数据处理
所有实验均设置3个平行,并采用SPSS 25.0对实验结果进行方差分析和显著性分析(P<0.05)。
2 结果与分析
2.1 酯化途径
微生物合成酯类物质的常见途径主要包括酯化、醇解、酸解和酯交换途径,酯化途径是指一种酸和一种醇在酯酶的作用下合成相应酯类的过程,酯酶是能够催化酯键水解和形成的酶的总称[8];醇解途径是指一种醇与甘油酯类物质在酯酶或酰基转移酶的作用下合成相应酯的过程,酰基转移酶可以将甘油酯上的酰基直接转移给醇,且酯酶在水环境下也可以发挥酰基转移酶的作用[9-10];酸解和酯交换途径通常用于合成具有营养功能的结构化油脂,与食品风味关系不大,本次实验中未考察。
在酯化反应体系中,地衣芽孢杆菌和埃切假丝酵母能够合成己酸乙酯和乙酸己酯,且产量随时间的增加而不断增长,由图1可知,实验菌株可以通过酯化途径合成酯类物质。随着反应时间的增加,各体系中的酯合成速率不断变化,但总体呈减缓趋势,表明在微生物酯酶的作用下,酯化反应作为可逆反应,逐渐趋向于平衡的状态。在反应60 min时,埃切假丝酵母合成己酸乙酯和乙酸己酯产量分别为101.546 μg/mL和1.573 μg/mL,均显著高于地衣芽孢杆菌,表明埃切假丝酵母的酯化能力远远高于地衣芽孢杆菌。在整个反应过程中,各菌株合成己酸乙酯的产量始终比乙酸己酯高,说明实验菌株通过酯化途径合成乙酯类物质的能力相对较强,这与Lu等[11]在郫县豆瓣中检测到大量乙酯类物质的实验结果一致。


图1 酯化体系中地衣芽孢杆菌(A)与埃切假丝酵母(B)合成酯类物质的产量Fig.1 The yield of esters synthesized by Bacillus licheniformis (A) and Candida etchellsii (B) in esterification systems
2.2 醇解途径
由图2可知,地衣芽孢杆菌和埃切假丝酵母均可在醇解体系中合成乙酸己酯,但其合成的途径存在两种可能:一是醇解途径,三乙酸甘油酯上的乙酰基在微生物酯酶或酰基转移酶的作用下,被直接转移至己醇而形成乙酸己酯;二是“两步”反应,即三乙酸甘油酯上的乙酰基被大量水解生成乙酸,随后乙酸与己醇反应形成乙酸己酯,这个过程的反应结果与醇解途径类似,但实为酯化途径[12]。因此,地衣芽孢杆菌和埃切假丝酵母是否可以通过醇解途径合成乙酸己酯还需要进一步分析。

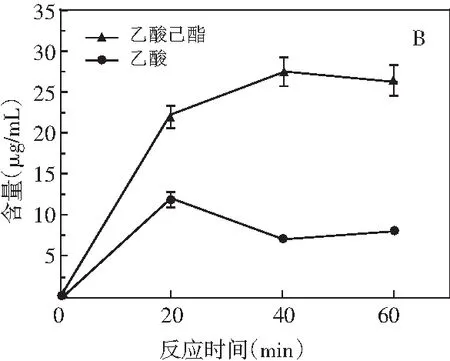
图2 醇解体系中地衣芽孢杆菌(A)和埃切假丝酵母(B)合成乙酸己酯和乙酸的含量Fig.2 The content of hexyl acetate and acetic acid synthesized by Bacillus licheniformis (A) and Candida etchellsii (B) in alcoholysis systems
在反应前期(0~20 min),乙酸己酯和乙酸的含量随时间的增长不断增加,其中乙酸己酯的增长速率和含量均高于乙酸,表明在地衣芽孢杆菌和埃切假丝酵母的作用下,三乙酸甘油酯上的乙酰基在反应前期被同时转移到己醇和水中,且转移至己醇合成乙酸己酯的速率大于其水解形成乙酸的速率,即在这个过程中有醇解反应的发生,表明这两株菌可以通过醇解途径合成酯类物质。Liu等[13]的研究中有类似的实验结果,酯合成速率及产量远高于脂肪酸的释放,相对于水分子,乙酰基可能更倾向于选择己醇作为目标底物,但也不能完全排除酯化反应的存在。反应20 min时,图2中A的乙酸含量达到最大值9.61 μg/mL,随后乙酸大幅度下降,乙酸己酯含量出现波动且在60 min时达到11.17 μg/mL,这可能是由于在地衣芽孢杆菌还原酶的作用下,乙酸形成的速率远低于被还原形成乙醇的速率,导致乙醇的大量增长(数据未显示),打破了体系的平衡状态,使乙酸己酯在反应40 min后发生了水解。由图2中B可知,乙酸己酯在20 min后以较低的速率继续增长,而乙酸含量下降,表明三乙酸甘油酯水解形成乙酸的速率远低于参与酯化反应消耗乙酸的速率;在反应40 min后,乙酸己酯发生水解,其含量小幅度下降,最终达到26.52 μg/mL,导致乙酸含量的适度增长,故在该体系的反应中后期有酯化反应的存在。
综合以上实验结果,埃切假丝酵母和地衣芽孢杆菌可以通过酯化和醇解两种途径合成酯类物质,具有较强的酯合成能力,不仅是郫县豆瓣中的优势产酯菌株,还广泛应用于酱油、白酒等发酵食品中发挥增香作用[14],相对来说,埃切假丝酵母的酯合成能力远高于地衣芽孢杆菌。此外,实验菌株通过酯化途径合成乙酸己酯的产量远低于醇解途径,这可能是由于水分子作为酯化反应的产物,相对于醇解反应来说,酯化反应在水环境中受到了一定程度的抑制,导致其产量相对较低[15]。
2.3 pH对菌株酯合成活力的影响
微生物无论通过酯化或醇解途径合成酯类物质,都是由相关的酶催化反应的发生,而pH是影响酶活力的重要因素,酯化体系和醇解体系中菌株的酯合成比活力受pH变化的影响情况见图3和图4。
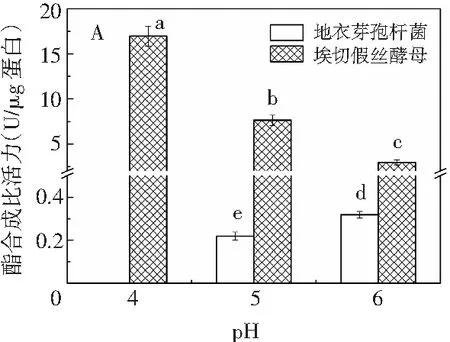

图3 不同pH条件下实验菌株在酯化体系中合成己酸乙酯(A)和乙酸己酯(B)的含量Fig.3 The content of ethyl hexanoate (A) and hexyl acetate (B) synthesized by experimental strains in esterification systems at different pH values
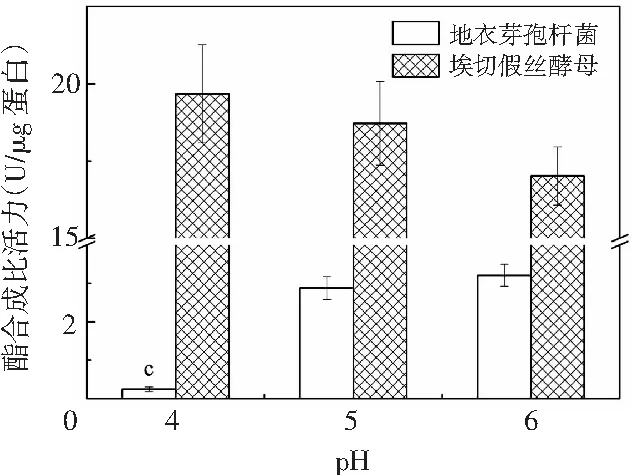
图4 不同pH条件下实验菌株在醇解体系中合成乙酸己酯的含量变化Fig.4 The content of hexyl acetate synthesized by experimental strains in alcoholysis systems at different pH values
在酯化体系中,随着pH值由4增加到6,埃切假丝酵母合成己酸乙酯和乙酸己酯的酯合成比活力不断降低,分别由16.956 U/μg蛋白降至2.965 U/μg蛋白、0.707 U/μg蛋白降至0.055 U/μg蛋白;此外,埃切假丝酵母在pH为4时通过酯化途径合成酯类物质的能力远高于另外两个pH值,表明埃切假丝酵母所产出的酯酶更倾向于在酸性环境中进行催化反应的发生。地衣芽孢杆菌合成乙酸己酯的酯合成比活力变化与埃切假丝酵母相反,其呈不断增加的趋势,由0.014 U/μg蛋白增至0.016 U/μg蛋白;由图3 中A可知,地衣芽孢杆菌在pH为4时,无法通过酯化途径合成己酸乙酯,随着pH值继续增长到6,合成己酸乙酯的酯合成比活力不断增加至0.318 U/μg蛋白。因此,地衣芽孢杆菌和埃切假丝酵母均更倾向于通过酯化途径合成己酸乙酯,其酯合成比活力远高于乙酸己酯。由图4可知,在醇解体系中,随着pH值的增加,地衣芽孢杆菌和埃切假丝酵母合成乙酸己酯的酯合成比活力的变化趋势与酯化体系类似,分别由0.245 U/μg蛋白增至3.210 U/μg蛋白、19.673 U/μg蛋白降至17.020 U/μg蛋白。因此,在pH值为4~6的范围内,地衣芽孢杆菌更倾向于在偏中性的条件下通过酯化或醇解途径合成酯类物质,而埃切假丝酵母在偏酸性的环境中酯合成比活力较强;此外,埃切假丝酵母的酯合成比活力在不同pH条件下仍远高于地衣芽孢杆菌,与2.1、2.2中的结果一致。
3 结论
地衣芽孢杆菌和埃切假丝酵母作为郫县豆瓣发酵过程中的优势菌株,可以通过酯化和醇解两种途径合成酯类物质,其中酯化途径合成乙酯类物质的能力相对较强,而乙酸酯类物质倾向于通过醇解途径合成。在pH为4~6的范围内,地衣芽孢杆菌酯合成活力随pH值的增加而增强,而埃切假丝酵母在pH为4时酯合成比活力最强。此外,在不同pH条件下,埃切假丝酵母通过酯化或醇解途径合成酯类物质的能力总是高于地衣芽孢杆菌。本研究通过探究地衣芽孢杆菌和埃切假丝酵母的酯合成途径及能力,为加速积累和改善发酵过程中的酯类物质和平衡郫县豆瓣的整体风味提供了理论依据。
