生态渔业养殖模式下汤逊湖鱼产力估算及对内源污染的影响
2021-03-23杜明普王红丽刘康福袁赛波张静李震宇阮锐
杜明普,王红丽,刘康福,袁赛波,张静,李震宇,阮锐
武汉市城市防洪勘测设计院有限公司
武汉市被称为“百湖之市”,可进行渔业养殖的水面面积达10.1万hm2[1],水产品年产量为50万t左右。传统湖泊渔业养殖多为密集养殖,因投肥、投饵等导致养殖水体中含有大量残饵、水产动物粪便、渔药等污染物[2],对湖泊水体造成污染,使水质恶化。汤逊湖是武汉市最大的城中湖,根据《第一次全国污染源普查水产养殖业污染源产排污系数手册》[3],按可养水面及武汉市平均鱼产量对该湖因传统渔业养殖产生的内源污染负荷量进行估算,不同养殖方式(如围网围栏养殖、网箱养殖)下总氮负荷量为108~965 ta,总磷负荷量为23~207 ta,约为汤逊湖TN和TP水环境容量的10%~91%和35%~313%(以主湖体水质达GB 3838—2002《地表水环境质量标准》Ⅳ类水质为基准)。传统渔业和密集养殖降低了汤逊湖水体自净能力,所造成的内源污染负荷量甚至超出水体的环境容量。而生态渔业养殖——不投肥、不投饵,利用湖泊天然饵料进行合理放养的渔业养殖模式可将水体中营养物转化成渔产品,从而减轻水体氮、磷污染负荷,因此逐渐被关注并应用到水污染控制工程中。
有学者在生态渔业养殖模式下,开展利用鱼类控制湖泊富营养化的理论和实践探索。如谢平[4]提出利用鲢(Hypophthalmichthysmolitrix)、鳙(Hypophthalmichthysnobilis)控制蓝藻的非经典生物操纵理论,当鲢、鳙的投放密度为46~50 gm3时,能有效控制东湖的水华发生,该理论被应用于上海陈行水库蓝藻控制、宝钢水库蓝藻控制等工程中,并取得一定效果[5];Domizon等[6]研究认为,鲢的放养密度为30~90 gm3时,浮游植物生物量和蓝藻生物量占比均明显减小。生态渔业养殖在水质管理和水体富营养化控制方面作用的发挥,与鱼类的放养密度有重要关系[7]。笔者根据汤逊湖现存天然饵料生物量进行鱼产力估算,提出生态渔业养殖中鱼类放养密度,并通过估算鱼类代谢产生的污染负荷量及生长增重存储营养盐的量,分析生态渔业养殖对内源污染负荷的影响,以期为水生态治理中充分利用生态渔业养殖发挥湖泊的净化功能提供参考。
1 数据来源与研究方法
1.1 研究区概况
汤逊湖位于湖北省武汉市东南部(114°15′E~114°35′E,30°22′N~30°30′N),分为内汤逊湖和外汤逊湖。汤逊湖水域面积为47.62 km2,平均水深为2.2 m,湖泊容积约10 360.7万m3。在《武汉市禁止渔业养殖湖泊目录》发布之前,汤逊湖区主要以围网围栏集约化养殖(投饵、投肥)为主,以鲢、鳙、鲤(Cyprinuscarpio)、鲫(Carassiusauratus)、鳊(Parabramispekinensis)、黄颡鱼(Pelteobagrusfulvidraco)、翘嘴鲌(Culteralburnus)、鳜(Sinipercachuatsi)等经济鱼类为主要养殖对象,其中鲢、鳙的放养量占总量的90%以上。由于高密度养殖以及周边大量污水汇入,汤逊湖在2014—2018年一直为Ⅴ类或劣Ⅴ类水质[8-10],处于中度富营养化水平,并在2017年暴发了严重的蓝藻水华。为控制富营养化,减少水华的发生,汤逊湖于2017年逐渐完成围网、围栏与网箱的拆除,将集约化渔业养殖转化为生态渔业养殖。2019年随着基于蓝藻水华控制的渔业结构调整等工作的开展,汤逊湖实施“以渔控藻”工程,投放了208万kg鱼苗,其中滤食性的鲢、鳙占97%,其他肉食性鱼类等占3%,并采取不投饵、不投肥的生态养殖模式。渔业结构调整后虽然汤逊湖局部水域仍存在蓝藻堆积的现象,总氮整体处于劣Ⅴ类水平,但总磷整体处于Ⅳ类~Ⅴ类水平,蓝藻水华暴发得到了明显的遏制[9]。
1.2 数据来源
汤逊湖天然饵料包括浮游植物、浮游动物、底栖动物、软体动物、有机碎屑及着生藻类等,其天然饵料现存量密度如表1所示。

表1 汤逊湖天然饵料现存量密度
1.3 研究方法
1.3.1鱼产力估算
鱼产力是指在不投饵、不投肥条件下,单位时间内单位水体面积中饵料基础转化为鱼产品的能力[15]。汤逊湖在生态渔业养殖模式下的鱼产力参照《湖泊渔业生态类型参数》[14]和《水库鱼产力评价标准》[15]进行估算。
汤逊湖浮游植物鱼产力(F浮游植物)按下式估算:
F浮游植物=FSC×EHy×Hy/c
(1)
FSC=PG×(PNa/PGt)×a×K
(2)
PG=33Cchla×LDH×HSD
(3)
式中:FSC为浮游植物对鲢、鳙的供饵能力,t(以氧计,全文同);EHy为鲢对浮游植物的能量转化率,取0.032;Hy为水中鲢占鲢、鳙数量的比例,取0.7;c为1 g鲢、鳙鲜肉所含热量,取值为5.021 kJ/g;PG为湖区浮游植物年生产量,t(以氧计,全文同),参考王骥等[16]的方法计算;PNa/PGt为浮游植物单位面积净产量与毛产量之比,取0.8;a为浮游植物最大利用率,取50%;K为1 g氧的热当量,取值为14.685 kJ/g;Cchla为叶绿素a浓度,内外汤逊湖分别为48.64和45.71 μg/L;LDH为平均日照时数,h;HSD为水体透明度,m,根据调查取0.45 m。
汤逊湖浮游动物鱼产力(F浮游动物)按下式估算:
F浮游动物=FSC×EAr×Ar/c
(4)
式中:EAr为鳙对浮游植物的能量转化率,取0.072;Ar为水中鳙占鲢、鳙数量的比例,取0.3。
汤逊湖底栖动物鱼产力按下式估算:
F底栖动物=(0.032BM+0.183BI+0.235BO)×S
(5)
式中:F底栖动物为底栖动物鱼产力,t;BM、BI和BO分别为软体动物、底栖水生昆虫、底栖寡毛类的生物量,g/m2;S为养殖面积,km2。
汤逊湖沉水植物鱼产力按下式估算:
Fg=Bp×Pp/(kp×100)
(6)
式中:Fg为沉水植物的供饵能力,kg/hm2;Bp为可被草食性鱼类利用的沉水植物的最高生物量,kg/hm2,由于实测调查汤逊湖沉水植物覆盖度低于50%,按平均生物量500 g/m2计算[14];Pp为计划利用的沉水植物生物量的占比,取60%;kp为草食性鱼类(长13 cm的鱼种)的平均饵料系数,取120。
汤逊湖有机碎屑鱼产力估算公式如下:
F=CsV(19.58%Hy+22.6%Ar)×
3 900 000/(3 560Hy+3 350Ar)
(7)
Cs=Ct-(BG+Bzp)×0.2×0.4
(8)
式中:F为有机碎屑提供的鲢、鳙鱼产力,t;Cs为有机碎屑中碳浓度,mg/L;V为水体表层10 m以内的水体容积,m3;Ct为有机碎屑中有机碳年平均浓度,mg/L,参考林婉莲等[17]的方法测定。由于汤逊湖有机碎屑现存量为47.68 mg/L,颗粒有机碎屑中有机碳浓度为4.08%[18],推测汤逊湖Ct为1.945 mg/L。BG和Bzp分别为浮游植物和浮游动物平均生物量,mg/L。
着生藻类的鱼产力采用下式估算:
F着生藻类=Bpb×(Pf/Bf)×U×S/k
(9)
式中:F着生藻类为着生藻类的鱼产力,t;Bpb为着生藻类平均生物量,μg/cm2;Pf/Bf为着生藻类年生产量与年平均生物量之比,也即Pf/Bf系数,取100;U为着生藻类最大利用率,取20%;k为着生藻类的饵料系数,取100。
1.3.2渔业养殖污染及营养盐存储量计算
鲢、鳙摄食天然饵料后,其中的氮、磷营养盐一部分被鱼类吸收利用转化为鱼体的增重部分(存储量),另一部分则随鱼的排泄和排粪过程重新参与水体的物质循环,被湖泊中的生物再次利用。每尾鲢、鳙因排泄和排粪的氮、磷估算参数如表2所示。鲢经过1年生长可增重1.05 kg,每增重1 g可存储氮26.29 mg,存储磷5.72 mg;鳙经过1年生长可增重1.3 kg,每增重1 g可存储氮26.40 mg,存储磷5.74 mg[19]。则:
渔业养殖产生的污染物量=鱼产力/个体年增质量×排泄和排粪回归水体氮、磷估算参数
(10)
鱼体内存储的营养盐量=鱼产力×单位质量鱼存储的营养盐量
(11)
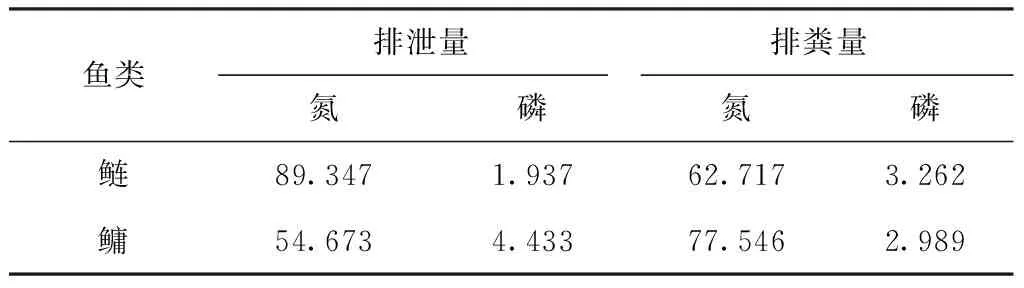
表2 鲢、鳙排泄和排粪回归水体的氮、磷估算参数
2 结果与讨论
2.1 汤逊湖鱼产力
根据汤逊湖天然饵料现存量,生态渔业养模式下天然饵料转化为鱼产力的能力如表3所示。由表3可知,汤逊湖总鱼产力为2 975.49 t/a,其中内、外汤逊湖滤食性鱼类鱼产力(主要为鲢、鳙的鱼产力,包括浮游植物鱼产力、浮游动物鱼产力及有机碎屑鱼产力)分别为1 059.05和1 715.9 t/a。2017年汤逊湖实施生态渔业养殖,内汤逊湖渔产量为900 t,平均产量为900 kg/hm2,其中鳙占40%,鲢占60%[9],该数据与表3的结果相近,鱼产力估算结果与实际捕捞情况相符,表明鱼产力估算结果一定程度上可靠。另外,滤食性鱼类鱼产力最高(占93.26%),而沉水植物鱼产力最低(占0.03%),该估算结果也与汤逊湖现状相符,即湖泊浮游植物及浮游动物饵料现存量大,沉水植物饵料现存量极少。
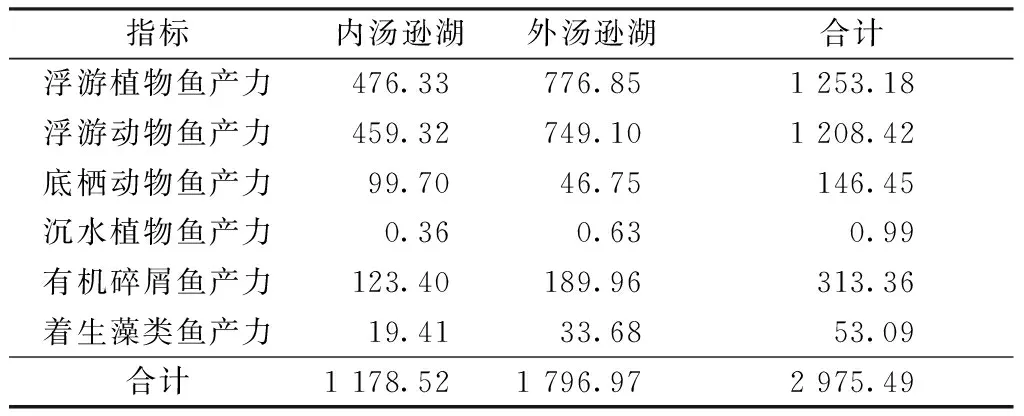
表3 生态渔业养殖模式下汤逊湖鱼产力
由表3可得,汤逊湖现状鱼产力密度为28.7 g/m3,低于2018年汤逊湖应急控藻的实际养殖密度(46 g/m3)[8];现状滤食性鱼类鱼产力密度为26.8 g/m3,低于谢平[4]提出的非经典生物操纵理论中滤食性鱼类放养的密度(46~50 g/m3)。随着汤逊湖控藻措施的实施,汤逊湖生物链将逐渐丰富与稳定,湖泊中浮游植物浓度将逐年降低,可能导致滤食性鱼产力逐年下降,但其他生物饵料可能逐渐增多,多种类型的鱼产力丰度将逐渐增大与稳定。因此,在利用渔业生态养殖控藻与净化水体时,与汤逊湖水质类似的湖泊,建议养殖密度控制在29 g/m3左右,并根据实测饵料量进行相应的动态调整,否则饵料现存量可能难以维持高密度养殖量。在养殖的动态调整过程中,可根据不同饵料类型,充分利用湖泊的其他鱼产力,促进湖泊营养物的快速循环。如随着湖泊富营养化得到控制,沉水植物将形成一定规模的斑块,可根据沉水植物生物量放养适量黄尾鲴(Xenocyprisdavidi)、细鳞鲴(Xenocyprismicrolepis)等,以充分利用植物有机碎屑等天然饵料;当沉水植物生物量足够高时,可放养适量草鱼(Ctenopharyngodonidellus)和团头鲂(Megalobramaamblycephala)等草食性鱼类,促使沉水植物体内营养盐向鱼体内转移,最终通过捕捞而去除。
2.2 生态渔业养殖产生的内源污染负荷
生态渔业养殖产生的内源污染负荷为鱼体排泄和排粪产生的营养盐量与鱼体内存储的营养盐量的差值。根据汤逊湖鱼产力估算结果,现状以鲢、鳙鱼产力为主,2015年至今主要养殖品种均为鲢、鳙[9],因此,生态渔业养殖中鱼类代谢产生的污染物主要考虑由鲢、鳙代谢产生。考虑到鱼粪中约有22%的氮、磷随幸存的活藻回归到浮游植物中[19],此部分为未被充分利用的天然饵料,应排除其对内源负荷的影响。根据鲢、鳙排泄和排粪回归水体氮、磷量,估算得出汤逊湖生态渔业养殖产生的内源污染负荷量,结果如表4所示。由表4可知,汤逊湖生态渔业养殖因排泄和排粪作用产生的内源氮、磷污染负荷量分别为309.28和13.06 t/a。以主湖区达Ⅳ类水质为基准时,汤逊湖总氮、总磷的水环境容量分别为1 061.5、66.23 t/a[8],氮、磷污染负荷量分别为总氮、总磷水环境容量的29.1%和19.7%,占用了汤逊湖部分自净容量。
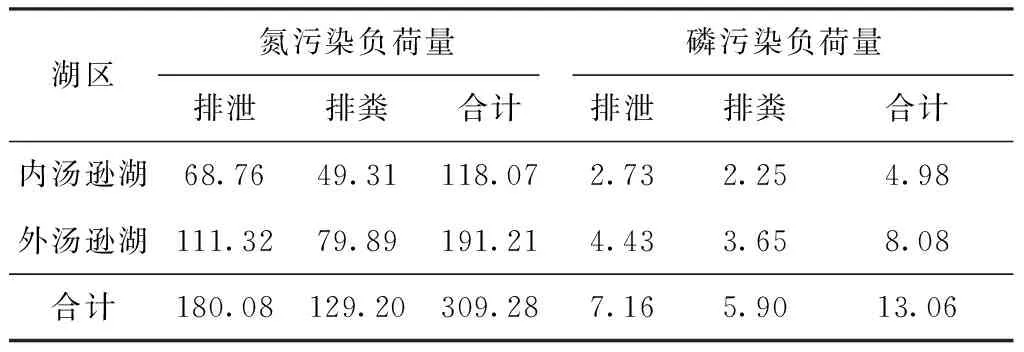
表4 鲢、鳙产生的内源污染负荷量估算
汤逊湖生态渔业养殖过程中,部分饵料通过鱼类增重而存储在鱼体内,使水体内的营养盐转移至渔产品中,并通过合理捕捞被移出湖泊。根据汤逊湖鲢、鳙鱼产力估算结果,当养殖周期为1年时,鲢、鳙因增重存储在体内的氮、磷量如表5所示。由表5结合表4可知,汤逊湖生态渔业养殖过程中,因排泄和排粪作用产生的氮量高于因增重存储在鱼体内的氮量,但因增重而存储在鱼体内的磷量超过排泄和排粪作用产生的磷量。可见,生态渔业养殖过程中,存储在鱼体内的营养盐磷的量高于转化为内源磷的量。
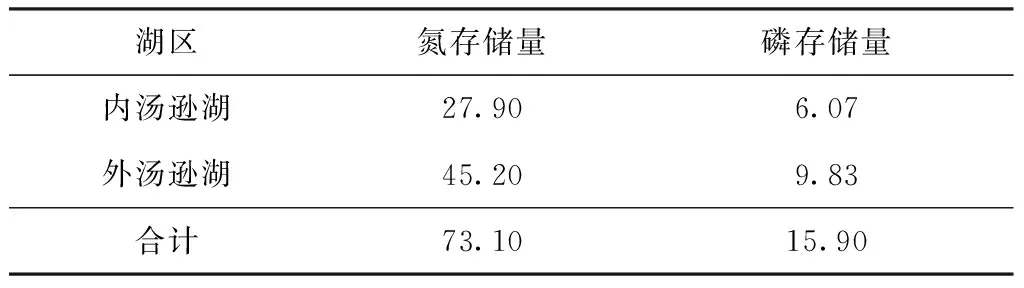
表5 鲢、鳙增重存储的营养盐的量
2.3 生态渔业养殖对内源污染负荷削减的贡献
生态渔业养殖模式下,汤逊湖鱼体内存储的氮、磷量分别为73.10、15.90 t/a,分别占入湖氮、磷污染负荷量(3 896.9、281.99 t/a)的1.88%和5.64%;部分氮、磷通过排泄和排粪作用重新回到水体,或通过渔业捕捞被移出水体,可见,鱼类在湖泊养分循环过程中起着重要作用。
鲢、鳙排泄和排粪产生的氮量高于存储在鱼体内的氮量,因此会对水体产生一定的氮内源污染。鲢、鳙主要摄食生命形态的氮、磷(浮游动物和浮游植物中含的氮、磷),将其转化为非生命形态的代谢物(如氨、尿素和尿酸)[19],其中氨和尿素易被微生物及其他生物利用,经过氨化、硝化/反硝化等作用转化为气态氮化物逸出水体被去除[20]。因此生态渔业养殖有利于内源氮的去除。鲢、鳙排泄和排粪产生的磷量低于存储在鱼体的磷量,由于渔产品可被移出湖体,因此可认为生态渔业养殖总体不贡献内源磷污染,而是可削减内源磷污染。
通常湖泊内源的去除可采用黏土矿物覆盖、化学药剂钝化以及底泥疏浚措施,或通过微生物以及水生植物吸收转换等方法[21]。在水生植物净化工程中,沉水植物吸收氮、磷营养盐的能力分别为700和小于100 kg/(hm2·a)[22],若要在沉水植物体内存储汤逊湖生态渔业养殖产生的营养盐,至少需要恢复159 hm2沉水植物,然而由于沉水植物生长受到环境等自然规律的限制,其修复过程中对内源污染物净化的作用缓慢[23]。相比之下,生态渔业养殖捕捞渔产品移除湖泊内源污染物是一种经济可行、易操作的技术,以鲢、鳙为主要养殖对象的生态渔业养殖适合作为沉水植物修复早期湖泊内源污染削减措施,通过放养和捕捞实现持续移出湖泊营养盐。
3 结论
(1)汤逊湖总鱼产力为2 975.49 t/a,生态渔业养殖模式下,建议鱼类养殖密度控制在29 g/m3左右。
(2)生态渔业养殖会产生一定量的氮内源污染,但能削减水体中磷内源污染。在湖泊水污染治理中,生态渔业养殖经济可行、易操作,适合作为湖泊生态修复早期湖泊内源污染削减的技术。
