ATP敏感钾通道负向调节海马长时程增强的维持
2021-03-19张晓琳张小云贺桂琴周子凯
张晓琳,张小云,贺桂琴,孔 月,周子凯
上海交通大学医学院附属精神卫生中心,上海200030
记忆长期存储的分子基础是脑科学的核心科学问题之一。突触可塑性是指神经细胞间的连接,即突触的功能和形态,尤其是突触传递强度可随着自身活动的加强与减弱相应发生较为持久的改变的特性或现象。突触可塑性被认为是大脑高级功能例如学习和记忆的基础。主要实验模型包括长时程增强(long-term potentiation,LTP)和长时程抑制(long-term depression,LTD),并以LTP 为主[1-5]。LTP 与记忆类似,都可分为诱导、维持和消退3个阶段。不同阶段不仅在时程上区分明确,也各自对应着截然不同的复杂的分子调控机制。长期以来,针对LTP 分子机制开展的大量研究主要集中于诱导阶段,发现了许多重要信号分子和转导通路,包括钙调蛋白依赖性蛋白激酶Ⅱ(calcium/calmodulin-dependent kinaseⅡ,CaMKⅡ)、蛋白激酶A(protein kinase A,PKA)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、磷脂酰肌醇3 激酶(phosphoinositide 3-kinase,PI3K)等[1,6]。相较而言,关于LTP维持的关键分子研究较少,近年来主要发现有蛋白激酶Mζ(protein kinase Mζ,PKMζ) 和 蛋 白 激 酶Cι/λ (protein kinase Cι/λ,PKCι/λ)[7-10]。
ATP 敏 感 钾 通 道(ATP-sensitive potassium channel,KATP)是一类非电压依赖性的配体门控离子通道。KATP是由2种不同的亚基以1∶1的比例组成的异源八聚体,即4个内向整流钾通道(inwardly rectified potassium channel,Kir6.x)亚基与4 个磺脲类受体(sulfonylurea receptor,SUR)亚基形成的一个功能通道。KATP在体内分布广泛,尤其在中枢神经系统中高表达,主要分布于皮层、海马体、纹状体、黑质和小脑等[11]。在正常生理情况下,细胞内ATP 充足,与KATP位于Kir 亚基上的结合位点相结合,抑制KATP通道的开放,KATP处于关闭状态;而当细胞代谢异常时,ATP/ADP 比值下降,KATP通道开放,引起K+外流,使得细胞膜超极化,降低神经元兴奋性,减少电活动[12]。因此KATP起到关联神经元细胞代谢状态与电活动的作用。迄今,KATP在多种神经系统疾病中的作用都有研究报道,包括癫痫[13]、帕金森病[14]、阿尔茨海默病[15]、病理学疼痛[16]等。一般认为KATP通道的激活开放可以降低神经元兴奋性,在上述病理条件下减轻能量代谢需求,从而起到神经保护作用。
既往研究已对KATP在多种脑疾病中的作用和机制进行探讨,但其在突触可塑性中的作用不明。此外,多种代谢异常时的大脑病理状态,例如脑缺血、脑缺氧时,都可造成记忆受损,但具体机制不清。本研究采用急性海马切片的电生理记录为主要研究手段,检测KATP在LTP不同阶段中的调节作用;并采用瞬时氧糖剥夺(oxygen glucose deprivation,OGD)模型探讨KATP在代谢异常条件下引起LTP功能障碍过程中的作用。
1 材料与方法
1.1 实验材料
1.1.1 实验动物 5~6 周龄健康雄性C57BL/6 小鼠,由南京大学-南京生物医药研究院(原南京大学模式动物中心)提供。实验动物生产许可证号为SCXK(苏)2018-0008。实验动物使用许可证号为SYXK(沪)2018-0028。饲养条件为12 h 光照/12 h 黑暗、20~25 ℃、相对湿度40%~70%。研究符合上海交通大学医学院附属精神卫生中心实验动物伦理操作规范。
1.1.2 主要试剂 人工脑脊液成分:NaCl 120 mmol/L,KCl 3 mmol/L,MgSO41.2 mmol/L,NaH2PO41 mmol/L,NaHCO326 mmol/L,CaCl22 mmol/L,D-葡萄糖11 mmol/L。KATP激活剂克罗卡林(cromakalim,CRO)、KATP阻断剂甲苯磺丁脲(tolbutamide,TOL)及所有人工脑脊液成分试剂均购自美国Sigma-Aldrich公司。
1.1.3 主要仪器 Multiclamp 700B 膜片钳放大器(Molecular Devices,美国),Digidata 1440A 数模转换器(Molecular Devices,美国),Grass S88 电刺激器(A-M Systems,美国),Leica VT1200 半自动振动式切片机(Leica Biosystems Inc.,美国)。
1.2 实验方法
1.2.1 急性海马脑片制备 小鼠断颈处死后,将大脑快速剥离并移至95%O2和5%CO2混合气预饱和的4 ℃人工脑脊液中。1 min 后取出,将鼠脑从正中矢向切开,左右两半球分别用乐泰404胶粘在振动式切片机载物槽的平台上,以350µm厚度切片。随后置于孵育槽内,在28 ℃混合气预饱和的人工脑脊液中恢复至少2 h 用于电生理记录。
1.2.2 场电位电生理记录 所有兴奋性突触后场电位(field excitatory postsynaptic potential,fEPSP)记录都在急性海马切片CA3-CA1 投射通路进行。玻璃记录电极内充人工脑脊液。所有电生理记录实验组均选择来自6只小鼠的6张脑片(即n=6)。
基础水平的输入-输出曲线(input-output curve,I-O曲线)记录:以不同强度的单脉冲方波刺激激起CA1 锥体神经元的fEPSP,线下分析得出对应不同刺激强度的fEPSP降支10%~85%段的斜率后,规格化制得I-O曲线。
双脉冲易化(paired-pulse facilitation,PPF)记录:分 别 以 间 隔 为25、50、100、200、300、400、500、1 000 ms 的双脉冲方波刺激诱发fEPSP,线下分析得出fEPSP 降支10%~85%段的斜率后,以双脉冲后一fEPSP斜率除以前一fEPSP斜率,规格化制得PPF曲线。
LTP记录:在记录了稳定的20 min fEPSP基线后,立即给予持续2次100 Hz高频刺激,每次持续1 s,2次间隔10 s;随后继续记录fEPSP 60 min。
OGD 模型:用95% N2/5% CO2混合气处理的无糖人工脑脊液替换95%O2/5%CO2混合气处理的含糖人工脑脊液,灌流5 min。
1.3 统计学分析
所有数据都通过pCLAMP 10 电生理信号采集与分析软件(Molecular Devices,美国)分析。采用SigmaStat软件(Systat Software,美国)进行统计学分析。基础水平的突触传递在各刺激强度和各时间点上进行组间比较。LTP 的幅度以各组记录的最后10 min 取平均值后计算x±s并进行组间比较。组间比较采用student t 检验(Student's t test),P<0.05为差异有统计学意义。
2 结果
2.1 KATP开放不影响基础水平的突触传递
如图1A 所示,在KATP开放剂CRO(100 µmol/L)持续灌流的情况下,I-O 曲线上CRO 组对应各刺激强度的fEPSP 斜率与使用磷酸缓冲盐溶液(phosphate buffer saline,PBS)灌流的对照组相比,差异无统计学意义(P>0.05)。说明当脑片处于基础状态时,突触传递不受KATP开放的影响。由于研究[11]报道KATP亚细胞定位于突触前,可能对突触前的神经递质囊泡释放过程起调节作用,于是我们检测了CRO 对PPF 这一经典的突触前功能指标的影响。
如图1B 所示,当双脉冲方波刺激之间时间间隔较短时,例如200 ms 以下时,第二个突触后反应的fEPSP 斜率与第一个反应的fEPSP 斜率的比值大于100%,表明存在PPF。随着时间间隔的增加,这一比值趋近于100%。CRO 组与PBS 对照组差异并无统计学意义(P>0.05)。上述实验说明KATP的异常开放不影响基础水平的突触传递。
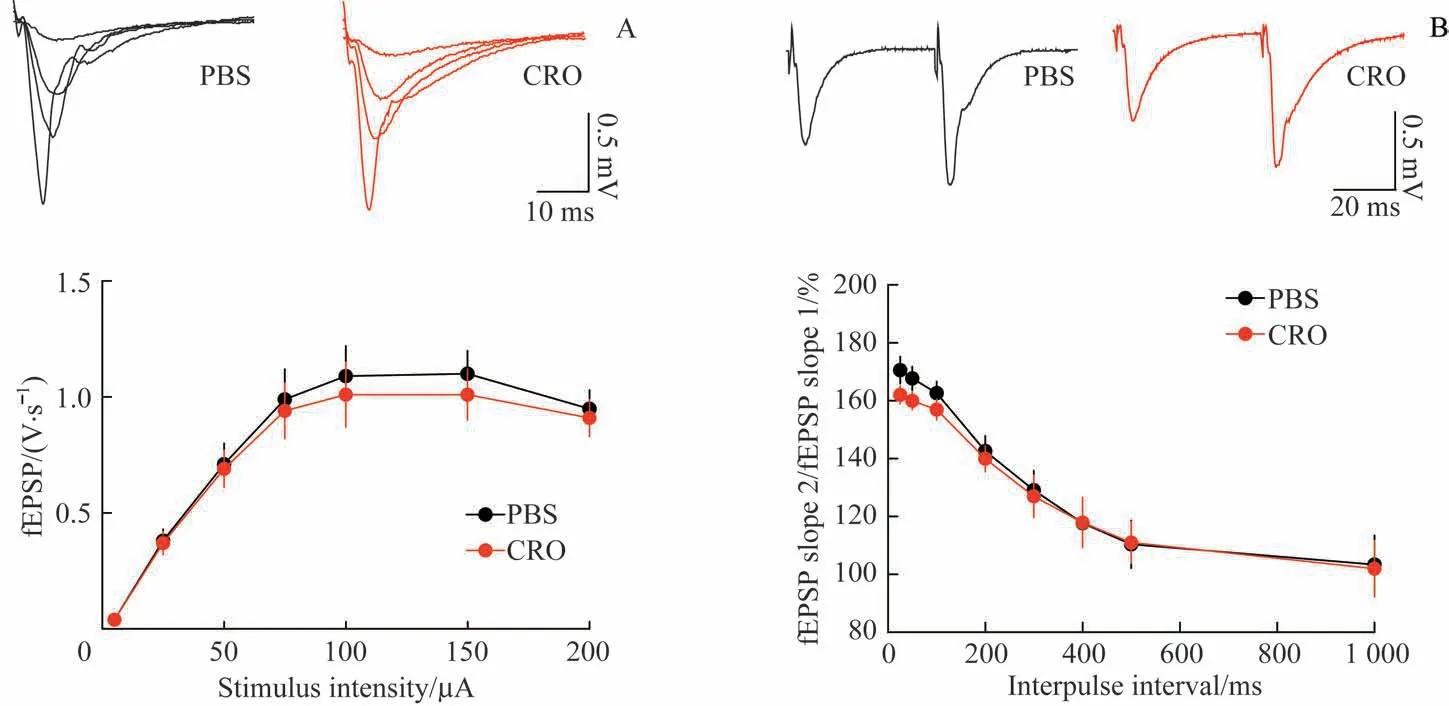
图1 KATP开放剂CRO不改变基础水平的突触传递Fig 1 KATP opener CRO did not modify synaptic properties at basal level.
2.2 KATP负向调节LTP的维持
为了检测KATP在LTP的诱导和维持阶段中的作用,分别在LTP 的诱导和维持阶段灌流含有KATP开放剂CRO(100µmol/L)的人工脑脊液,并观察其作用。如图2A 所示,高频刺激可以诱导出至少持续60 min的突触传递增强,即LTP。根据高频刺激后50~60 min 的fEPSP 斜率,发现CRO造成的KATP通道开放并未影响LTP的幅度。然而,当LTP产生并稳定后,在图2B所示的维持阶段,即40 min时间点起开始灌流CRO,可显著降低LTP 的幅度[PBS 组fEPSP 斜率=(141.4±6.9)%,CRO 组fEPSP 斜率=(118.2±6.4)%,P=0.010 6],而KATP阻断剂TOL 对LTP 的幅度无影响。这一结果说明KATP的开放状态对LTP的诱导过程无影响,但是KATP保持关闭对LTP正常幅度的维持十分关键,因此KATP对LTP维持的调节作用属于负向调节。
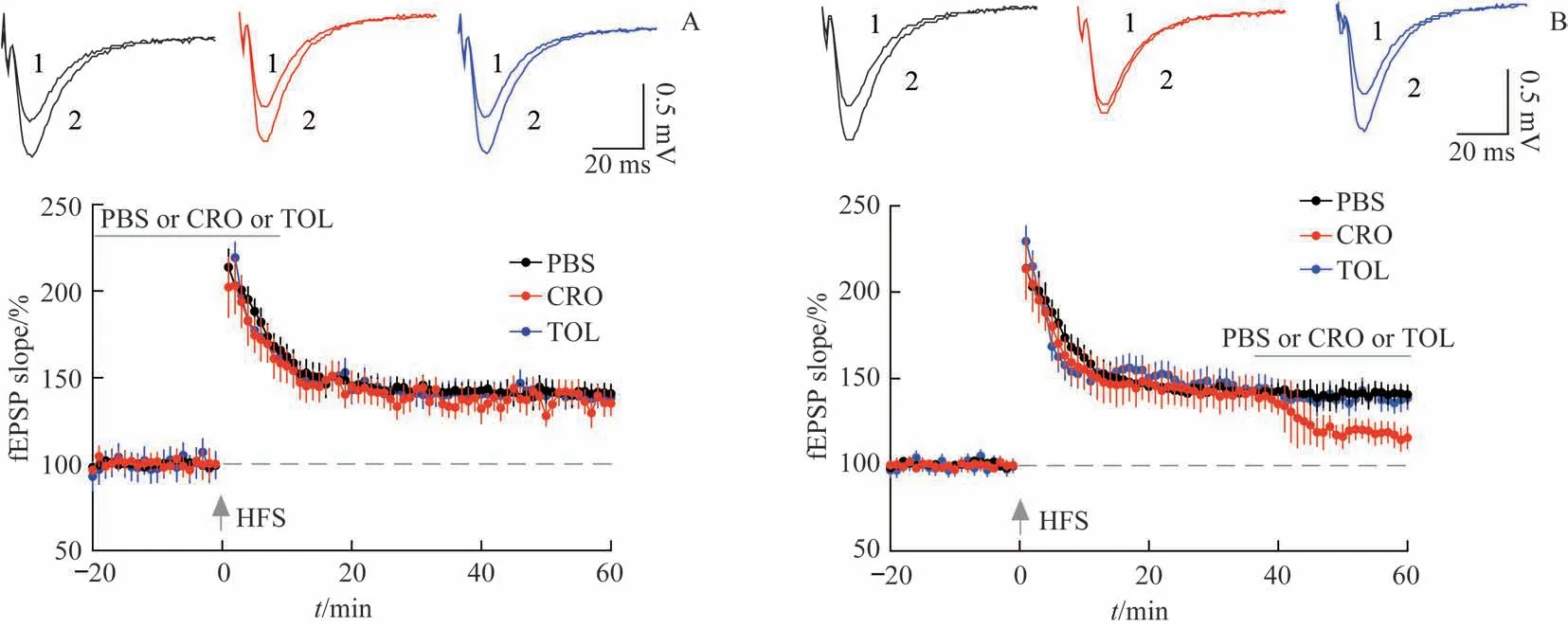
图2 KATP开放剂CRO选择性降低LTP维持阶段的fEPSP幅度Fig 2 KATP opener CRO selectively reduced LTP magnitude at the maintenance stage
2.3 OGD通过异常开放KATP破坏LTP的维持
上述实验结果发现LTP的维持需要KATP保持关闭,而脑缺血缺氧状态又公认可造成显著记忆损伤。因此,我们猜想脑缺血状态有可能是通过异常激活KATP破坏LTP的维持,最终损伤记忆。为了验证这一猜想,我们对脑片进行OGD 来模拟在体脑缺血缺氧状态。如图3A 所示,当给予高频刺激诱导LTP 稳定40 min 后,OGD 可造成fEPSP 反应的幅度快速、显著减弱(图3A)。当撤去OGD 后,fEPSP 可在数分钟内快速恢复至正常水平。可见OGD 可以破坏LTP 的维持。图3B 显示在OGD 前10 min 开始进行TOL 预处理,阻断KATP可以显著缓解OGD 对LTP 的 维 持 造 成 的 影 响[OGD 组fEPSP 斜 率=(139.4±6.5)% , TOL+OGD 组fEPSP 斜 率= (127.1±6.3)%,P<0.05]。
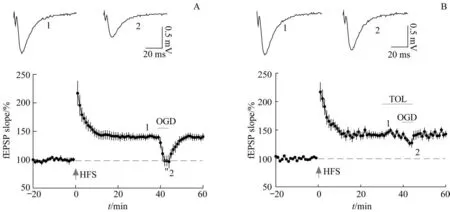
图3 OGD损伤LTP的维持过程且可通过阻断KATP预防Fig 3 OGD impaired LTP maintenance that can be largely prevented by pre-treatment of KATP blocker
3 讨论
学习和记忆是大脑最基本的高级功能之一。在细胞和分子水平介导记忆存储的神经生物学机制长期以来一直是脑科学的研究热点。在记忆的实验模型LTP 的维持阶段,一般认为快反应α-氨基-3-羟基-5-甲基-4-异唑(α-amino-3-hydroxy-5-methyl-4-isox-azolepropionic acid,AMPA)型谷氨酸受体持续向细胞膜转运和cAMP 反应原件 结 合 蛋 白 (cyclic-AMP response element-binding protein,CREB)介导的转录调控及蛋白合成过程是突触增强得以长时程维持的关键。研究[7]表明PKMζ 在海马脑片LTP 及长时程联合记忆的维持中起到了决定性的作用,被称为“记忆分子”。随后大量研究[9-10]发现PKMζ和PKCι/λ 这2 种非典型PKC 可相互代偿,影响LTP 和海马相关学习记忆的不同阶段。
本研究发现,作为联接细胞代谢状态和神经元活性的关键分子,KATP保持关闭状态也是LTP得以维持的重要条件。与其他已知的LTP维持机制不同的是,KATP主要分布于突触前膜,因此其造成的fEPSP 突触反应快速减弱主要可以归因于突触前神经递质的释放受到抑制。这与发生在突触后的AMPA 型谷氨酸受体转运以及细胞核内CREB介导的转录过程并行存在,在不同的亚细胞组分上以不同的机制共同调节LTP的维持。本研究发现,KATP可能也是一个关键的“记忆分子”,起负向调节作用。进一步通过动物行为学研究检测KATP的开放状态是否与PKMζ和PKCι/λ 的失活类似,以及是否会“擦除”空间记忆、恐惧记忆等,是非常必要的。
此外,本研究发现KATP的开放状态不影响基础状态的突触传递,也不影响LTP 的诱导过程,说明LTP 的诱导过程可能不涉及KATP介导的神经元兴奋性改变。而在LTP的巩固阶段则可能发生了改变神经元膜兴奋性的细胞生物学过程,因此LTP的维持阶段选择性地受到KATP开放状态的影响。由于离体脑片记录需持续灌流糖、氧饱和的人工脑脊液,无法完全模拟在体代谢状态,因此在体记录活体动物在体电生理记录,如多通道方式记录在体LTP,可能提供更多的实验证据支持这一结论。
在异常细胞能量代谢状态下,如脑缺血缺氧时,ATP的快速耗竭可有效激活KATP通道,引起K+外流,使得细胞膜电位超极化,降低谷氨酸的过度释放以及随之而来的兴奋毒性效应,产生神经保护效应。该效应已在癫痫[13]、帕金森病[14]、阿尔茨海默病[15]、病理学疼痛[16]等病理状态有研究报道。阿尔茨海默病是一种以认知障碍和记忆减退为特征的神经退行性疾病。Moriguchi等[15]研究发现阿尔茨海默病治疗的一线药物美金刚(memantine)可直接调节神经元KATP通道中Kir6.2亚基的活性,并提出美金刚的起效机制是通过抑制Kir6.2通道开放来恢复LTP,并改善阿尔茨海默病患者记忆障碍。这与本研究中OGD 的实验结果及推论一致,即KATP的病理性开放有损神经可塑性,而抑制KATP可能缓解记忆障碍。值得注意的是,若长期过度抑制KATP的开放,理论上可能又会加重神经元的损伤,甚至引起其他并发症,如癫痫。但是,阿尔茨海默病患者大脑的兴奋性与抑制性突触传递平衡状态显著异于正常大脑。美金刚在实际应用中是如何通过阻断N-甲基-D-天冬氨酸(N-methyl-Daspartate,NMDA)型谷氨酸受体和KATP通道起效的具体机制,以及造成癫痫的可能性还有待进一步研究证实。
综上,本研究表明在生理或病理条件下,KATP的激活均可负向调节LTP 的维持过程。该结果为记忆存储的分子机制提供了新的认识,也为代谢异常型脑损伤造成的记忆障碍提供了新的认识。
