甘草来源内生真菌多样性和抗菌活性研究
2021-03-08刘文杰李陇强唐佳慧姜淑玲陈春晨郭文强
刘文杰,李陇强,唐佳慧,姜淑玲,杨 平,陈春晨,郭文强
井冈山大学医学部,吉安 343009
深色有隔内生真菌(dark septate endophytes,DSE)作为一群定殖于植物根内且具有深色菌丝和明显横隔的特殊真菌,其生活史中的某段时期存在于植物根的表皮、皮层甚至维管束组织的细胞内或细胞间隙中,并能够形成独特的微菌核,但不会在根组织内转化成病原菌而引起植物病理学特征[1]。大部分植物体内的内生真菌都属于DSE范畴,随着自然界长期的进化选择,该类真菌与宿主之间形成了动态平衡关系。在植物的生长周期中,内生真菌在一定程度上具有促进植物生长、提高宿主植物的抗病虫害以及抵御环境压力的作用。同时,内生真菌与宿主植物次生代谢产物的积累也有密不可分的关系[2]。DSE广泛分布于地球的所有生境中,越来越多的研究表明,DSE可能具有与根际土壤真菌类似的生态学功能,在不同生境下的微生态系统中发挥着重要共生作用,由此也可能具有异于普通生境下真菌的代谢途径,能够产生结构新型、活性独特的天然产物[3,4]。
甘草属于豆科甘草属多年生草本植物,具有补脾益气、清热解毒、祛痰止咳的功效,主要道地产区分布在宁夏、甘肃、内蒙古和新疆等地,因特殊的药效而被广泛使用[5]。而不同产地甘草药材在品质上存在差异,其主要原因可能为外部环境因子如产地气候、土壤等造成产地差异性[6]。近年来,关于甘草中的DSE功能研究逐渐引起了人们的重视,相关研究表明,从甘草内生真菌发酵物中发现了与宿主相同的有效成分,如甘草酸、甘草苷、异甘草苷、甘草次酸、甘草素等[7-9]。此外,从甘草中分离的内生真菌中筛选具有生物活性的菌株,聚焦其活性化合物也已成为天然产物研究的新方向[10,11]。鉴于此,本研究将展开对不同产地甘草来源内生真菌种群多样性和抗菌活性研究。
1 材料和方法
1.1 材料
1.1.1 实验样品
新鲜健康的乌拉尔甘草样品分别于2018年9月、10月采集于内蒙古赤峰市、甘肃张掖市、宁夏吴忠市。甘草样品采集后置于4 °C冰箱中保藏,并且由北京中医药大学孙志蓉教授鉴定为乌拉尔甘草。
1.1.2 培养基
固体培养基:马铃薯葡萄糖琼脂培养基(PDA)。
液体发酵培养基(真菌2号):葡萄糖(10 g/L),麦芽糖(20 g/L),谷氨酸钠(10 g/L),甘露糖醇(20 g/L),KH2PO4(0.5 g/L),玉米浆(1 g/L),酵母浸粉(3 g/L),MgSO4·7H2O(0.3 g/L)和蒸馏水1L,调节pH为6.5。
1.1.3 实验仪器
超净工作台SW-CJ-2D(苏州苏洁净化设备有限公司);高压灭菌器KXQ-LS(上海博讯有限公司);电热恒温培养箱DNP-9082P03Ⅳ(上海新苗医疗器械制造有限公司);旋转蒸发器RE-2000A(郑州科泰实验设备有限公司);低温冷却液循环泵DLSK-5/20(郑州科泰实验设备有限公司);循环水式多用真空泵SHK-Ⅲ(郑州科泰实验设备有限公司);超声波清洗器KQ-300E;电子天平BSA-124SNMR。
1.2 方法
1.2.1 内生真菌分离纯化
甘草样品经过75%酒精浸泡处理1 min后无菌水冲洗干净,加水充分研碎并吸取上清液,置于马铃薯葡萄糖琼脂培养基上于28 °C孵育3~5天。从中分离出具有不同形态的真菌菌落,将其重新接种到新的PDA培养基,并在28 °C下再孵育培养36 h。根据在PDA平板上的菌落形态(颜色、质地、边界类型和径向生长速率等),将其分为不同的形态类型,最终得到内生真菌纯菌株。
1.2.2 内生真菌种属鉴定
将数据由Gardner模型拟合得到如图2所示的曲线,图中各点表示实测数据,曲线为拟合结果,各点与曲线基本重合。
从该来源的内生真菌中提取DNA并进行PCR反应,实验采用通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)扩增内部转录的间隔区(ITS)[12-14]。PCR产物由睿博生物科技有限公司(中国,北京)纯化和测序。通过BLAST分析将鉴定出的序列与GenBank数据库(http://www.ncbi.nlm.nih.gov)中的其他序列进行比较分析,并在ClustalW中进行比对[15]。利用Godinho提出的标准来分析验证GenBank数据库的BLAST结果:对于查询覆盖率和序列同一性≥98%,该物种种属可以被确定;对于覆盖率和序列同一性在95%~97%之间的可以确定到种,而对于覆盖率和序列同一性≤95%,则可以认为是新菌[16]。系统进化分析则是采用MEGA X进行的,N-J法被用来估计种属进化距离,其自举值是根据1 000次重复计算而得出的结果[17]。
1.2.3 内生真菌发酵培养与处理
将各菌株接种到含有100 mL真菌2号液体培养基的500 mL三角瓶中,28 °C静置发酵培养30天。发酵完成后将发酵混合物用等体积的乙酸乙酯超声萃取2次,经减压蒸馏获得粗提物后,甲醇溶解,并用微孔滤膜过滤除去不溶固体杂质。挥干溶剂后称重,用甲醇重新溶解,配成浓度为5 mg/mL的溶液,低温冷藏备用。
1.2.4 指纹图谱分析
真菌粗提物在HPLC系统(Waters Co.)中分析,该系统包含996型二极管阵列紫外检测器和ODS-C18色谱柱(YMC,4.6 mm×250 mm,5 μm)。40 min内从5%~100%甲醇梯度洗脱,并在100%甲醇条件下继续洗脱10 min。
1.2.5 抗菌活性测试
以草分支杆菌、枯草芽孢杆菌、耻垢分枝杆菌、金黄色葡萄球菌、大肠埃希氏杆菌作为抗菌活性指示菌,利用纸片扩散法进行粗提物抗菌活性筛选与评价。氯霉素(0.1 μg/μL)被用作抗菌活性测定的阳性对照。将纸片(直径10 mm)吸取菌株甲醇粗提液后挥干,并放置在鉴定菌琼脂表面上。37 ℃下培养24 h后,测量五种鉴定菌下纸片的生长抑制区直径。
2 结果与讨论
2.1 甘草来源内生真菌种属分析与鉴定
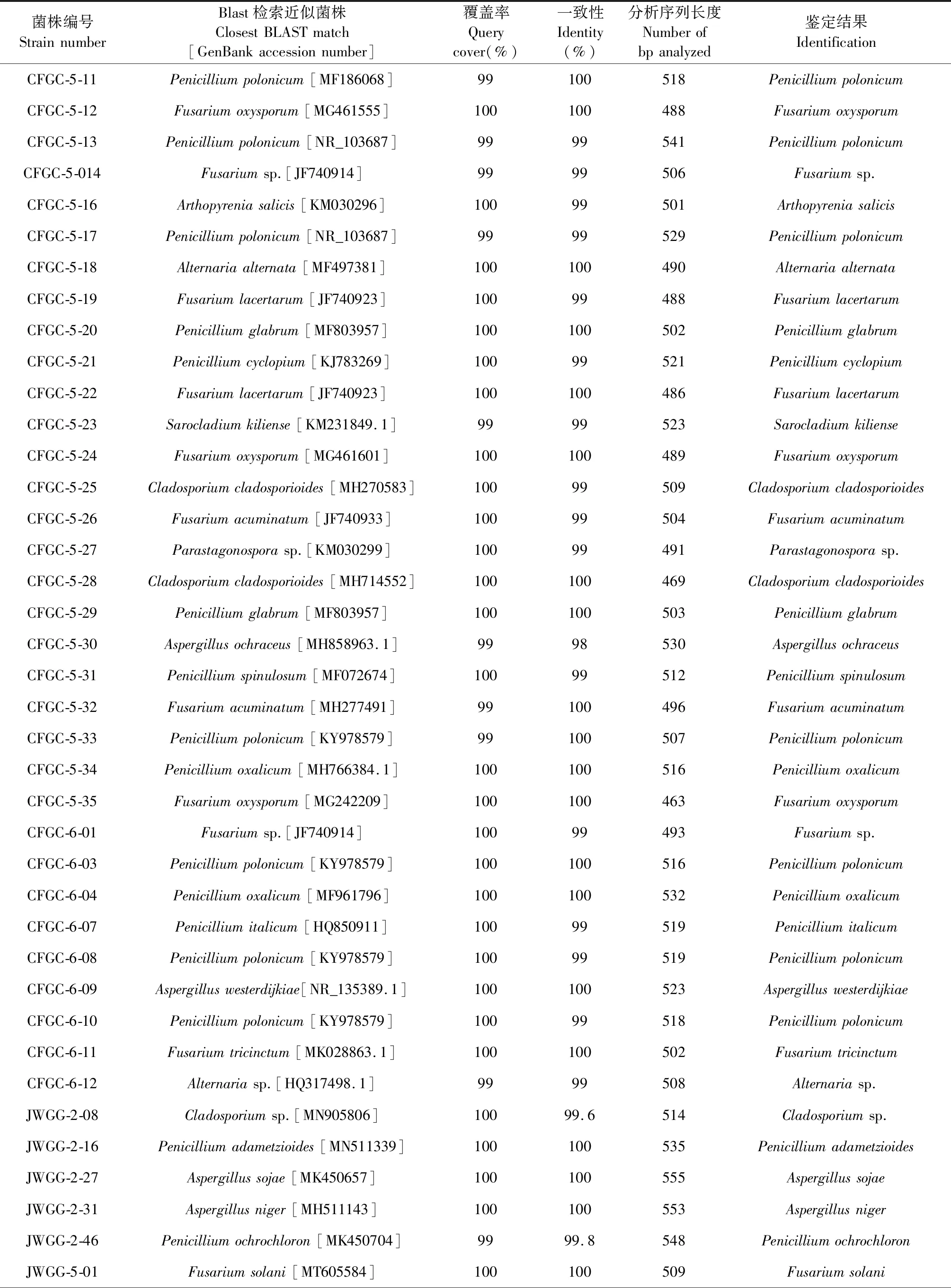
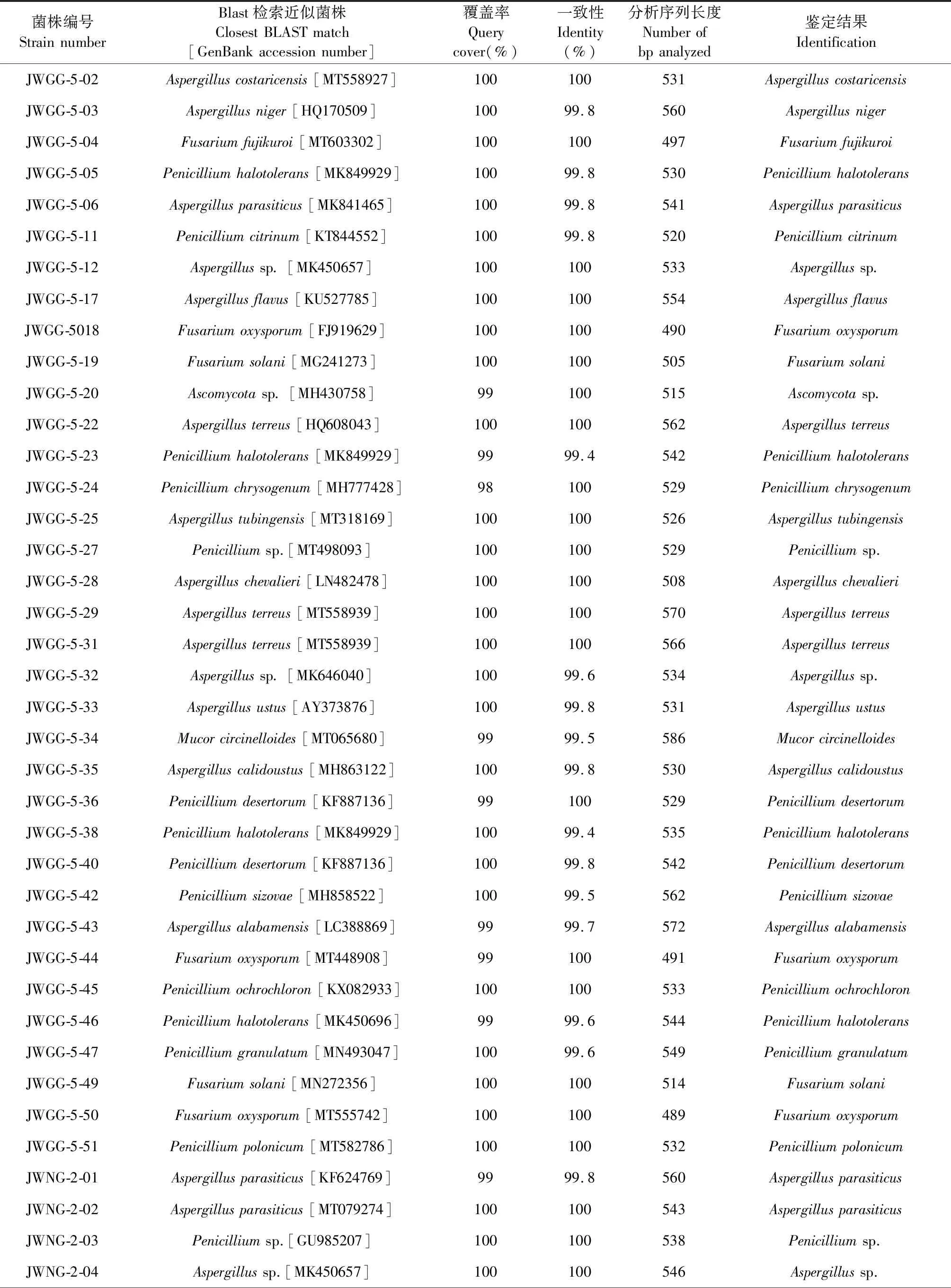
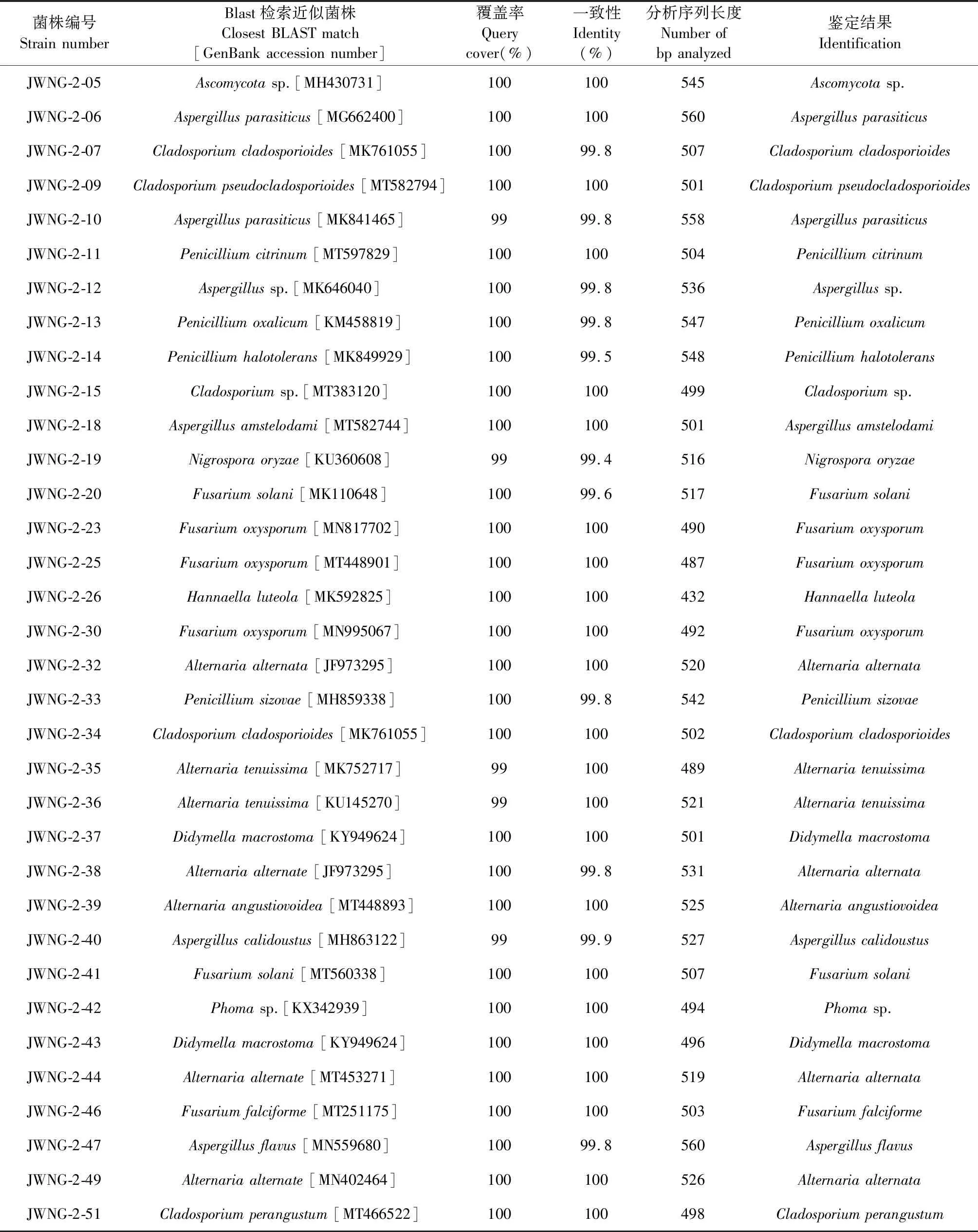
对该来源的134株内生真菌进行序列分析发现其包含子囊菌类和接合菌类两大真菌类群。依据Godinho的分类标准,对提交到GenBank数据库的序列与相似菌株进行覆盖率和相似度的比对。结果可知大部分菌株与其最接近的种属展示了较高的相似度,可以确定到种属,由此发现不同产地甘草内生真菌群落组成也各不相同。内蒙产地的55株甘草内生真菌归属于11个种属(Penicillium、Aspergillus、Byssochlamys、Talaromyces、Cladosporium、Fusarium、Clonostachys、Arthopyrenia、Alternaria、Sarocladium、Parastagonospora),其中以青霉属和镰刀菌属为优势菌属,分别占分离菌株总数的41.81%和16.36%。甘肃产地的41株内生真菌隶属于6个种属(Cladosporium、Penicillium、Aspergillus、Fusarium、Ascomycota、Mucor),其中以曲霉属为优势类群,占分离菌株总数的39.02%。宁夏产地的38株内生真菌隶属于9个种属(Penicillium、Aspergillus、Ascomycota、Cladosporium、Nigrospora、Hannaella、Alternaria、Didymella、Phoma),以曲霉属为优势菌,占分离菌株总数的23.68%。综上,青霉和曲霉属为各地甘草内生真菌的优势菌属,但各产地之间真菌种属间差异性较大,初步发现甘肃产地甘草的内生真菌种群多样性较其他两地低(见表1)。
续表1(Continued Tab.1)
续表1(Continued Tab.1)
续表1(Continued Tab.1)
将所有甘草内生菌株的ITS序列输入MEGA X进行进化分析,并通过构建系统发生树来对甘肃、内蒙和宁夏的甘草内生真菌的亲缘性进行种系发育研究(见图1~3)。

图1 内蒙甘草来源内生真菌的系统发生关系Fig.1 Phylogenetic relationship of endophytic fungi from G.uralensis in Inner Mongolia

图2 甘肃甘草来源内生真菌的系统发生关系Fig.2 Phylogenetic relationship of endophytic fungi from G.uralensis in Gansu
2.2 甘草来源内生真菌抗菌活性
将所有菌株采用真菌2号培养基静置发酵30天后,获得134个代谢粗提物。在对其进行五种鉴定菌的抗菌活性筛选后,发现35株内生真菌的次级代谢产物表现出较好的抗菌活性,综合活性率为26.1%。其中菌株AspergilluswesterdijkiaeCFGC-5-04、Fusariumsp.CFGC-5-14、Parastagonosporasp.CF
GC-5-27、FusariumoxysporumJWGG-5-50、DidymellamacrostomaJWNG-2-37、AlternariaangustiovoideaJWNG-2-39表现出广谱的抗菌活性,同时的Cladosporiumsp.JWNG-2-15和Fusariumsp.CFGC-5-14的代谢产物对金黄色葡萄球菌显示出较好的抑制活性(见表2),显示出潜在的代谢产物研究价值。
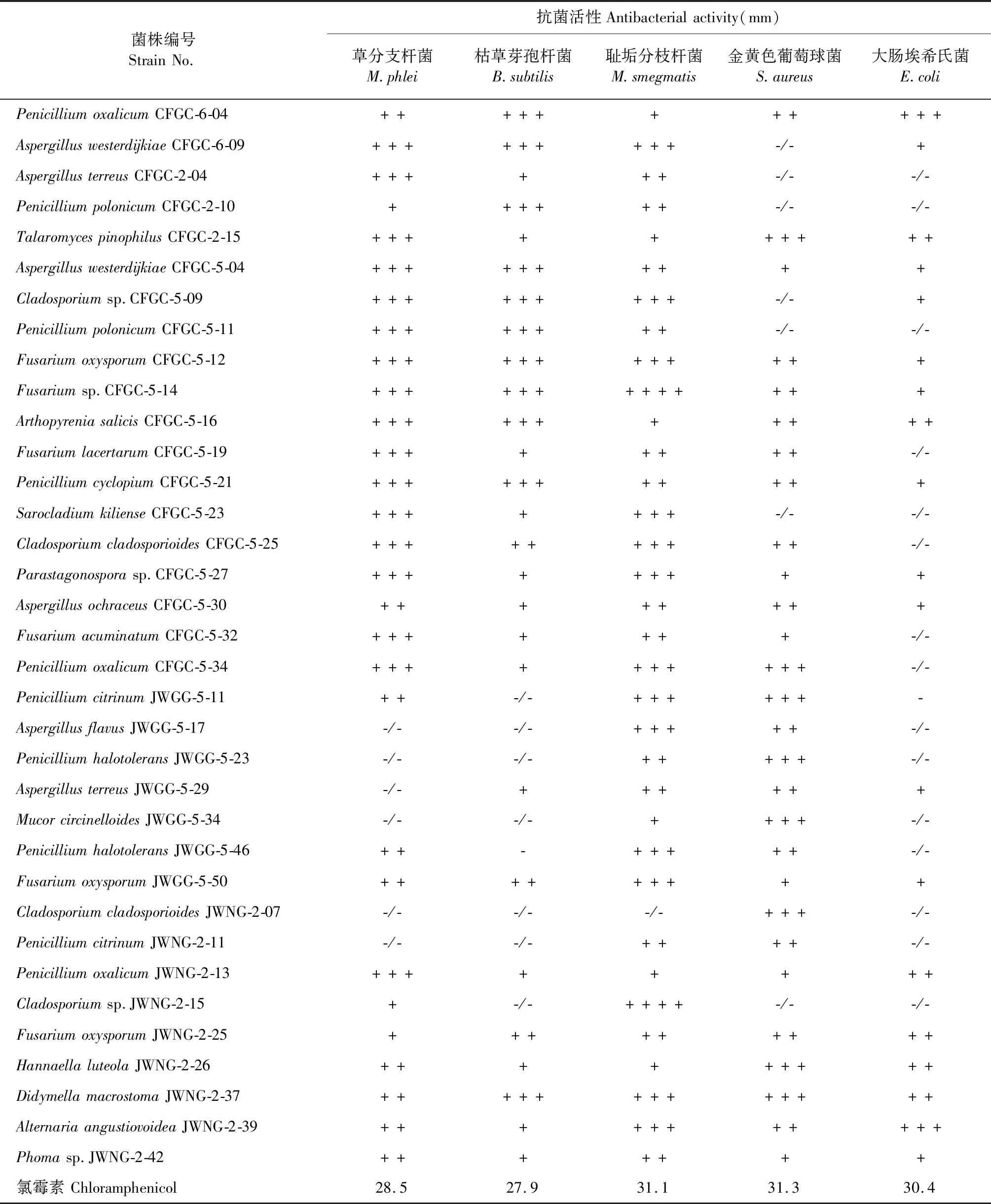
表2 甘草来源内生真菌次级代谢产物抗菌活性Table 2 Antibacterial activity of secondary metabolites of endophytic fungi from G.uralensis
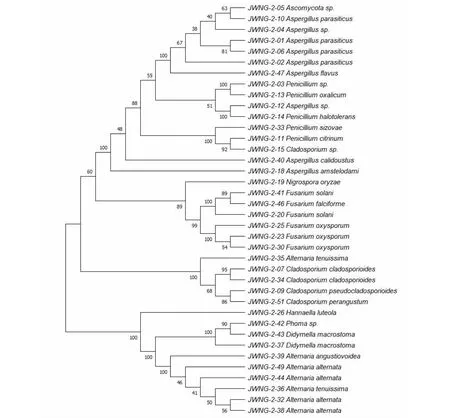
图3 宁夏甘草来源内生真菌的系统发生关系Fig.3 Phylogenetic relationship of endophytic fungi from G.uralensis in Ningxia
2.3 内生真菌次级代谢产物分析
通过对所有甘草来源内生真菌的次级代谢产物的HPLC-UV图谱进行分析后,发现其有多个菌株的代谢产物具有特征的指纹图谱(见图4)。
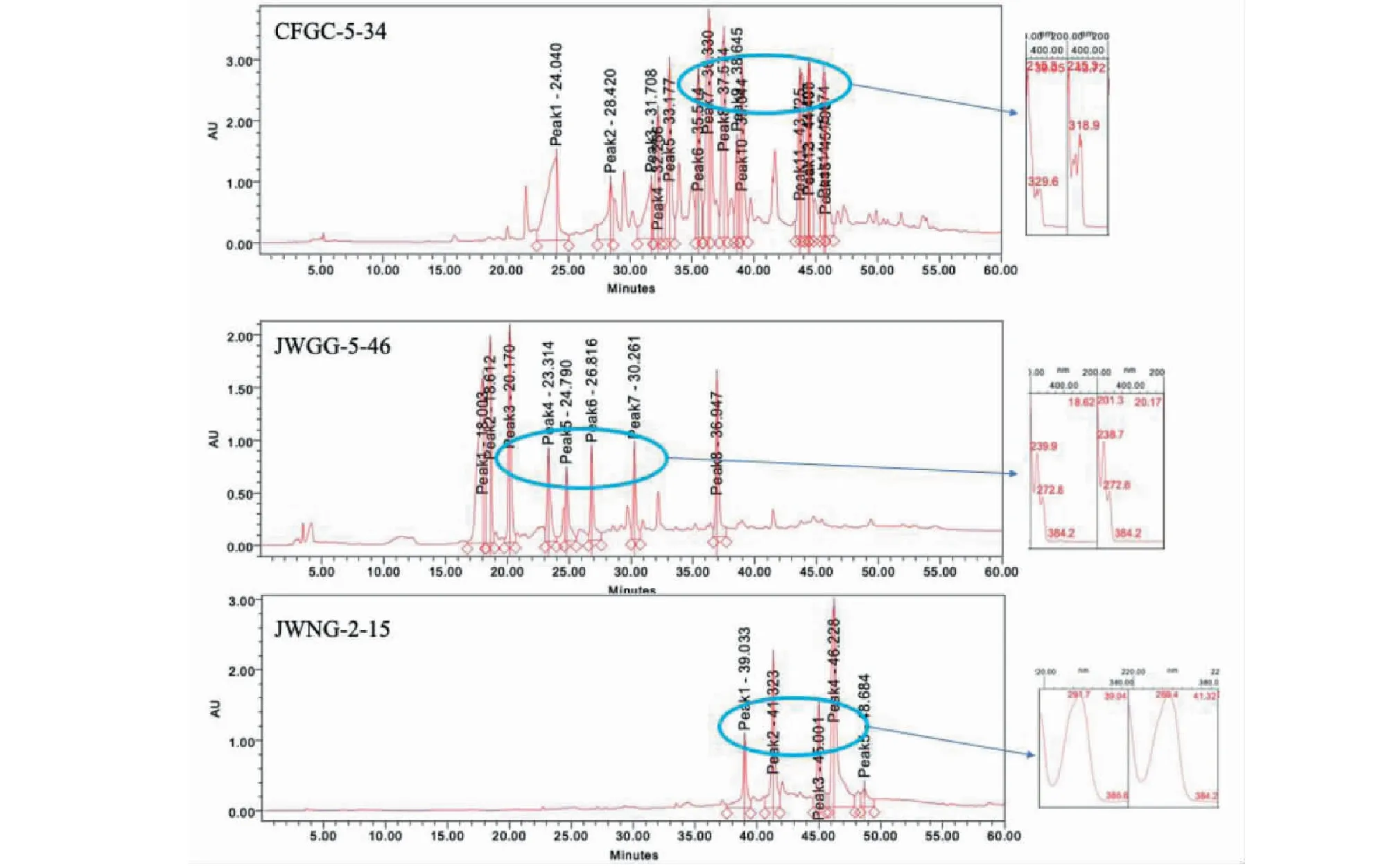
图4 部分内生真菌次级代谢产物HPLC-UV指纹图谱Fig.4 Comparison of metabolite fingerprints using HPLC-UV in representative endophytic fungi
其中活性菌株PenicilliumoxalicumCFGC-5-34、PenicilliumhalotoleransJWGG-5-46、Cladosporiumsp.JWNG-2-15的指纹图谱中分别表现出λmax320 nm、λmax270 nm、λmax285 nm的系列的紫外吸收,预示着活性内生真菌次级代谢产物中具有系列的天然产物。
3 讨论
已有研究表明,DSE的分布范围包含了不同的生境:从沿海滩涂到内陆高原山地;从热带、温带到冻原地区及南北极地区均有DSE的分布。越来越多的研究结果支持DSE与宿主植物之间能形成互惠共生的内在关系[1]。并且,DSE能够促进植物对土壤营养物质的吸收利用、增强植物的抗逆性(包括抗旱、抗寒、耐重金属等)、增强植物的抗病能力、影响植物群落、对不良环境植被的恢复和重建都有促进作用[18]。近年来,传统中药中DSE越来越受到重视,其不仅在植物生理学、代谢组学等方面表现出巨大的研究价值,还被发现是良好生物活性天然产物的重要资源库。
本研究以不同产地的甘草样品为研究对象,研究其可培养内生真菌的物种多样性和生物活性。与以往的研究相似,我们发现该来源DSE群落中既有优势种,也有稀有真菌种,属于地方性、冷适应型和世界性类群,表现出有趣而独特的生态学特征。此外,我们还发现了一些具有高度生物活性的菌株,有待进一步研究其次级代谢产物的结构和活性的多样性,这些未开发的DSE菌株可能是独特的微生物资源。
通过对不同产地甘草内生真菌进行分离纯化来考察其生物多样性,在参考已有报道研究中对甘草内生真菌分离方法的基础上[5],对甘肃、内蒙和宁夏甘草主产地的DSE进行分析研究发现3个产区分离率、真菌种属多样性等方面都具有较大的差异性,其显示出可能的甘草地域性与内生真菌种属之间的潜在关系。对于中药的道地性等问题上,传统观点认为道地药材是植物基因和环境相互作用的产物,多集中于特定的地理因素或气候因素等植物外环境的作用,很少从植物内环境方面进行考虑,而内生菌又是植物内环境的重要组成部分[19]。研究也推测DSE是道地药材在特定区域分布的重要原因之一,也是道地药材与非道地药材品质差异的一个潜在原因。同时还发现植物DSE能够影响中药材的道地性,能够提高药用植物对某些次级代谢产物的累积[20,21]。
对不同产地甘草内生真菌的次级代谢产物的抗菌活性筛选与评价,发现各个产地甘草的内生真菌对多种病原菌表现出较好的抑菌活性,同时HPLC-UV分析也发现多个系列类型的天然产物存在,这在挖掘其次级代谢产物的结构多样性和生物活性方面具有潜在的研究价值。因此,分析甘草DSE的种属多样性,对相关菌株代谢产物的深入研究仍有必要,其对于甘草的道地性意义阐明和活性天然产物挖掘研究都具有重要的意义。
