未分化型甲状腺癌差异表达基因的生物信息学分析*
2021-03-03陈妹妹梁瑜祯李争明
陈妹妹,梁瑜祯,李争明,夏 宁△
(1.广西医科大学第一附属医院内分泌科,南宁 530021;2.广西医科大学第二附属医院内分泌科,南宁 530007)
未分化型甲状腺癌(anaplastic thyroid carcino‐ma,ATC)由《内分泌器官肿瘤WHO 分类》定义为由未分化的滤泡性甲状腺细胞组成的高度侵袭性甲状腺恶性肿瘤[1]。ATC 是一种极其罕见的恶性肿瘤,占甲状腺癌的1%~2%。它具有侵袭性强、恶性程度高、治疗效果及预后极差和生存率低等特点[2]。该病通常发生在老年人身上,平均年龄在60岁左右,女性居多[3]。目前尚无有效的治疗方法可以治愈或延长ATC 患者的生存。近年来,随着基因芯片、高通量测序、基因微阵列等技术的发展,为寻找ATC 发生发展相关差异表达基因提供了充分的条件。目前,关于ATC 的分子机制研究较少,因此,本研究旨在通过生物信息学方法来挖掘与ATC 发生发展相关的关键基因及其信号通路,以期为ATC的诊断及其治疗提供依据。
1 资料和方法
1.1 数据来源 从GEO 数据库(http://www.ncbi.nlm.nih.gov/geo)中检索种属是人的与ATC 相关的基因芯片数据库,共检索出GSE85457、GSE65144和GSE53072 3 个基因芯片。GSE85457 包含4 个ATC 组织样本和3个正常组织样本,GSE65144包含12 个ATC 组织样本和13 个正常组织样本,GSE53072 包含5 个ATC 组织样本和4 个正常组织样本。
1.2 差异表达基因的筛选及数据处理 GEO2R(http://www.ncbi.nlm.nih.gov/geo/geo2r)是GEO 数据库的一个在线分析工具,它可以直接对GEO数据中两个或两个以上的样本数据进行差异表达基因分析。以P<0.01、logFC(fold change)>1 或log‐FC<-1为筛选条件,筛选得到ATC组织样本与正常组织样本对比后的差异表达基因,然后进一步用韦恩图在线分析工具(http://bioinformatics.psb.ugent.be/webtools/Venn/),得到3 个数据集中都包含的差异表达基因,即共同差异基因同时得到绘制的Venn图。
1.3 共同差异表达基因的GO 富集分析和KEGG通路分析 DAVID(http://david.ncifcrf.gov)(版本6.8)是一个在线生物信息数据库,它结合了生物学数据和分析工具,并提供一套全面的基因和蛋白质的注释信息。GO 是一个重要的用于注释基因和分析这些基因参与的生物学过程的生物信息学工具。KEGG 是一个用于了解高级功能和生物系统,由大型分子数据集生成的基因组测序和其他高通量实验技术的实用程序数据库资源。利用DAVID对共同差异表达基因进行GO 富集分析,包括分子功能、细胞组成和生物过程3个方面,以及KEGG 通路富集分析,P<0.05认为差异有统计学意义。
1.4 共同差异表达基因的蛋白质互作(PPI)网络分析和显著基因模块分析 利用在线分析工具STRING(http://www.string-db.org/)(版本11.0),构建共同差异表达基因的PPI 网络图。进而应用Cyto‐scape(版本3.6.1)软件进行可视化分析,并应用其中的插件MCODE 筛选显著基因模块。MCODE 插件设置参数为:Degree Cut-off:2,Node Score Cut-off:0.2,K-Core:2 and Max Depth:100。
1.5 关键基因的筛选和分析 应用Cytoscape(版本3.6.1)软件的插件cytoHubba 对共同差异表达基因筛选出节点度(node degree)>10 的关键基因。基因节点度越高表示该基因与ATC 的关系越密切。应用DAVID 对关键基因进行GO 富集分析,包括分子功能、细胞组成和生物过程3 个方面,以及KEGG 通路富集分析,P<0.05 为差异有统计学意义。应用UCSC 癌症基因组数据库(http://genomecancer.ucsc.edu)构建层次聚类图。
2 结果
2.1 差异表达基因的筛选结果 通过对基因芯片数据对比筛选过后,GSE85457、GSE65144 和GSE53072分别获得1941、4653 和3052 个差异表达基因。然后用Venn 图取交集获得328 个共同差异表达基因(图1)。其中上调基因有103个,下调基因有225个。

图1 Venn图
2.2 共同差异表达基因的GO 富集分析和KEGG通路分析结果 通过DAVID 在线工具分析,GO 富集结果中分子功能主要包含金属离子结合、ATP 结合、蛋白质同源二聚活性、氧化还原酶活性和激酶活性等(图2A);细胞组成主要包含细胞质、等离子体膜、原生质膜的组成部分、粘着斑和细胞骨架等(图2B);生物过程主要包括信号转导、氧化还原过程、细胞粘附和细胞外基质的组织等(图2C)。KEGG通路分析结果表明共同差异表达基因主要参与神经营养素信号通路、结核、色氨酸代谢、赖氨酸降解和胃酸分泌等相关信号通路,见表1。
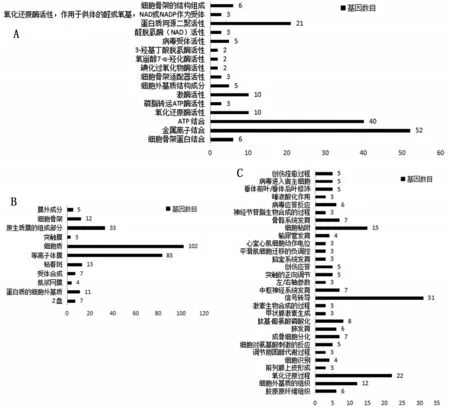
图2 共同差异表达基因的GO富集分析

表1 DGEs的KEGG通路分析
2.3 共同差异表达基因的PPI网络图和显著基因模块分析 构建共同差异表达基因的PPI 网络,其中上调基因有103 个,下调基因有225 个。并使用Cy‐toscape对PPI网络图获得最显著的基因模块(图3),本模块涉及的基因均是上调基因。
2.4 关键基因的筛选及分析 应用Cytoscape(版本3.6.1)软件的插件cytoHubba 对差异基因筛选出节点度>10 的关键基因,共筛选出19 个关键基因,其中上调基因有18 个,分别是AURKA、CDC6、WDHD1、OIP5、MCM4、NCAPH、RAD51AP1、CEN‐PF、FEN1、CDC7、NEK2、DSCC1、PAX6、KIF18A、CASP1、ICAM1、COL1A1、COL3A1,下调基因有1个,为FMOD。应用DAVID 对19个关键基因进行GO 富集分析及KEGG 通路富集分析,筛选出基因数≥5 的GO 富集分析,结果显示关键基因主要富集在蛋白结合、核浆、细胞核和细胞周期、DNA 复制,见表2、表3。应用UCSC 数据库对关键基因构建的层次聚类图结果显示,PAX6 和FMOD 在正常组织样本中高表达,CDC6 和NEK2 在肿瘤组织中高表达,关键基因可以基本区分甲状腺癌样本和正常样本(图4)。

表2 关键基因的GO富集
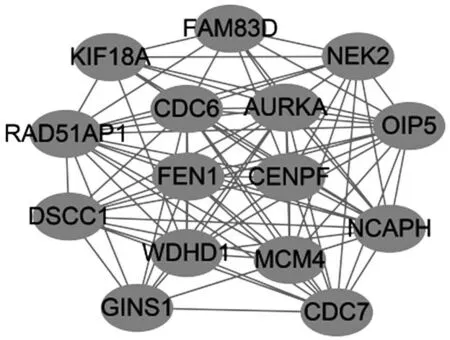
图3 PPI网络图中最显著模块图

表3 关键基因的KEGG通路分析

图4 层次聚类图
3 讨论
本研究通过对3个GSE基因芯片进行生物信息学分析,筛选得到328 个正常甲状腺组织与ATC 差异表达基因,其中上调基因有101 个,下调基因有227 个。对共同差异表达基因进行富集分析发现,基因主要参与金属离子结合、信号转导、氧化还原、细胞粘附等生物过程。金属离子可影响癌细胞端粒酶活性,而高水平的端粒酶与致癌转化相关[4]。信号转导异常是参与肿瘤发生发展过程的重要环节,而细胞间粘附降低则在癌细胞的侵袭过程和迁移过程中起到重要作用。细胞内氧化还原平衡的维持是细胞存活的关键,而肿瘤细胞内则因为抗氧化酶的过度表达,从而促进肿瘤的发生发展[5]。KEGG 富集分析表明,差异基因主要参与神经营养素信号通路和结核相关通路。有研究表明,神经营养因子参与肿瘤的增殖存活、迁移、转移和化疗耐药等过程[6]。结核杆菌感染与恶性肿瘤的发生有一定的相关性[7]。
共同差异表达基因通过构建PPI 网络图及分析,筛选出AURKA、CDC6、WDHD1、OIP5、MCM4、NCAPH、RAD51AP1、CENPF、FEN1、CDC7、NEK2、DSCC1、PAX6、KIF18A、CASP1、ICAM1、COL1A1、COL3A1、FMOD 19 个关键基因。关键基因富集分析显示,基因主要参与蛋白结合、细胞分裂、DNA 复制等过程。AURKA 是一种丝氨酸/苏氨酸激酶,通过在体内与中心体结合来调控有丝分裂,在有丝分裂进展的多个步骤中起着至关重要的作用,包括细胞周期G1 期的微管稳定性、染色体排列和分离以及胞质分裂,且经常在各种类型的人类癌症中异常表达[8]。有研究表明,AURKA 在ATC 中高表达,提示其可能在ATC 中发挥相关作用[9]。细胞周期调节因子在ATC 中有着重要作用。其中包括主要参与细胞周期进展或染色体不稳定的几个基因的表达:CDC6、MCM4、WDHD1、CDC7、CENPF、NEK2、DSCC1。CDC6属于ATP酶家族成员,有研究表明,CDC6 在甲状腺未分化癌、髓样癌、乳头状癌中阳性表达,且能反映出甲状腺癌细胞的增生程度,可辅助鉴别甲状腺腺瘤[10]。高水平表达的MCM4 与食管癌发生及病理分期有关[11]。抑制WDHD1 的表达可抑制肺癌和食管癌细胞的生长[12]。CDC7 和MCM4 在子宫内膜腺癌中有高表达的报道[13]。CENPF 全称着丝粒蛋白F,与细胞周期密切相关。CENPF 的高表达与乳腺癌预后差和肿瘤骨转移相关[14]。NEK2 属于NEK 蛋白激酶家族,参与细胞的有丝分裂,促进中心体成熟并维持其结构。有研究表明,在甲状腺肿瘤中NEK2 蛋白高表达[15]。DSSC1在结肠癌中过表达,且结肠癌细胞中DSSC1的高表达可降低患者的生存率[16]。根据这些结果,笔者推测这些细胞周期调节因子可能也在ATC 的发生、转移和预后中发挥重要作用。研究证实,OIP5在胶质母细胞瘤[17]、膀胱癌[18]、肾癌[19]等多种癌症中高表达,促进癌症的生长和转移,影响患者的预后。研究表明,PAX6 与恶性肿瘤的化疗耐药有关[20]。有研究发现,CASP1 与乳腺癌、肝癌和胰腺癌的免疫浸润程度相关[21]。COL1A1 在乳腺癌中高表达,且水平与癌症转移、生存率相关[22]。ICAM1,细胞间黏附分子1,可促进甲状腺癌细胞的增殖,且甲状腺乳头状癌(PTC)和ATC 都与ICAM-1 表达增加有关,ICAM-1 表达与不良预后密切相关[23-24]。NCAPH、RAD51AP1、FEN1、KIF18A、COL3A1、FMOD 目前在甲状腺癌中尚未有相关研究。但FMOD 被认为是淋巴瘤、白血病、平滑肌瘤等恶性肿瘤的一种新的肿瘤相关抗原[25]。
从数据库UCSC对关键基因的分析结果可以看出,PAX6 和FMOD 在正常组织样本中高表达。但层次聚类结果中显示PAX6 属于下调基因,与Cyto‐scape 筛选结果相反,原因可能与数据库UCSC 没有将甲状腺癌分类有关。数据库UCSC显示的结果是所有类型的甲状腺癌,而ATC 只占甲状腺癌1%~2%,从而导致PAX6 基因表达水平与Cytoscape 筛选结果相反。其余的关键基因基本上在正常组织低表达,在肿瘤组织中高表达。关键基因可以基本区分甲状腺癌样本和正常样本,进一步说明关键基因对ATC 是有明确影响的。但目前关键基因在ATC 中的研究还较少,很大原因可能是该病是罕见病导致样本数目少。但是绝大数关键基因在多种肿瘤中都有相关报道,与肿瘤的发生、进展、转移和耐药密切相关,因此我们推测关键基因也可能与ATC有着密切关系。
综上所述,本研究通过对ATC 的数据芯片的生物信息学分析,初步筛选出AURKA、MCM4、ICAM1、PAX6、FMOD 等19个基因在正常与癌组织中表达有明显差异,并探讨了关键基因参与的信号通路,有望成为ATC 诊断和治疗的潜在靶点。但由于本研究未对分析结果进行实验验证,因此,在未来的工作中,需要进一步的扩大样本数量并开展实验验证这些基因在ATC中的生物学功能。
猜你喜欢
杂志排行

广西医科大学学报的其它文章
- Effect of miR-200a-3p targeting transthyretin(TTR)on the biological behavior of placental trophoblasts in hypertensive disorder complicating pregnancy
- 沙利度胺对重型β地中海贫血患者红系细胞γ珠蛋白基因表达及分化的作用研究*
- CEMIP调控Src/AKT/STAT3信号通路对非小细胞肺癌上皮间质转化及细胞侵袭、迁移的影响*
- 乳铁蛋白对正畸复发移动过程中牙周改建的影响*
- 秦皮苷对脂多糖诱导的软骨细胞氧化应激和炎症因子的影响*
- miR-133b-3p对大鼠脑缺血/再灌注损伤的影响及其作用机制*