立地和龄组对四川省柏木人工林叶生态化学计量特征的影响
2021-03-02宋思梦
宋思梦 周 扬 张 健
(1. 四川民族学院农学院,横断山区生态修复与特色产业培育研究中心,康定 626001;2. 四川农业大学生态林业研究所,华西雨屏区生态环境监测站,成都 611130;3. 四川省林业生态工程重点实验室,长江上游生态安全协同创新中心,成都 611130)
柏木(Cupressus funebris)是我国亚热带地区主要造林树种之一,用途广,适应性强,具有极高经济、药用和园林观赏价值[1~2]。自20 世纪60 年代四川盆地荒山造林以来,柏木人工林在紫色砂泥岩发育的钙质紫色土区和碳酸盐类发育的石灰土区发挥了不可替代的生态功能[3],现主要分布在嘉陵江—渠江流域两岸的盆地北缘中低山区,已发展为该区域优势森林类型之一,在全省面积达204.47万hm2[1]。但柏木人工纯林带来多重效益的同时也面临着林地养分平衡较差[2]、生产力水平下降[3~5]及营养胁迫[1,6]等多种生态安全问题,生态系统服务功能整体较低。生态化学计量学(Ecological stoichiometry)是表征生态系统养分循环状态及植物与环境间协同变化规律的重要理论工具,可在一定程度上反映植物对结构性元素C 和限制性元素N、P 的平衡与利用能力[7~8]。C 是有机体干物质主要元素[9],N与植物光合效率密切相关,P是植物细胞分裂过程的必需元素[10],而C∶N,C∶P,N∶P不仅决定了有机体关键特征,更指示了其对有机质及矿物质元素等资源数量与种类的需求状况[9,11~12]。因此,开展柏木人工林生态化学计量特征的研究有助于认识区域尺度人工林养分需求利用水平,指导人们进行植物养分管理并促进柏木人工林可持续性经营。
立地是在一定空间范围内对林木生长有重大意义的环境条件综合体,包括地貌地形、土壤类型与理化性质等自然生境因子[13]。立地分类则是将相同立地类型归类合并,同一类中的林分拥有相近或相似的生产潜力,可以采用相同手段抚育管理[14]。目前人们对立地的研究主要为立地分类方法[13,15]、立地类型划分[16~18]、立地指数编制[19~20],在立地质量评价方面多集中于立地与生产力[21]、林草适宜性[22~23]、植物生长及水分关系[24]等,少有研究将立地与植物化学计量互作关系联系在一起,特别是在区域尺度柏木人工林中未见报道。立地条件与生态系统植物营养元素的耦合关系决定了植物生长发育过程和营养水平[13],柏木人工林中不同立地条件林分生长表现因海拔[15]、坡向[16]、坡位及小地形[14,19]等不同,可能在C、N、P 元素分配格局上存在很大差异,且随林龄增加这种差异性可因生态系统组成、内部环境及养分供给水平的变化进行重组[21,23]。因此,本研究以四川省不同立地与龄组内柏木人工林为研究对象,测定分析柏木叶C、N、P化学计量特征,拟回答以下科学问题:①四川省柏木人工林叶C、N、P 含量及C∶N、C∶P、N∶P;②同一龄组内,立地对柏木人工林叶生态化学计量的影响;③同一立地内,龄组对柏木人工林叶生态化学计量的影响。探讨柏木在化学计量分配策略上的适生立地类型,为四川省柏木人工林栽培的养分管理、规划布局和生产指导提供依据。
1 材料和方法
1.1 研究区概况
研究区位于四川盆地的柏木人工林集中分布区域(29°22′~32°52′N,104°16′~106°45′E,海拔750~1 488 m),是长江上游重要的生态安全屏障和水土流失严重的生态脆弱区。气候属盆地亚热带湿润季风气候,四季分明。盆北(苍溪、剑阁、梓潼)、盆中(大英、旌阳、南部)、盆南(威远)年降雨量分别为800~1 000、900~950、1 000~1 100 mm,年日照时间分别为1 300~1 400、1 000~1 300、1 150~1 200 h,年均温分别为16~17℃、15~16℃、17~18℃,无霜期分别为220~260、270~290、320~350 d。研究区地貌多为深丘、中丘、浅丘和中低山,地质为中生代陆相红层,以浅变质砂岩、硅质岩、页岩和泥岩为主。土壤类型多为酸性紫色土与石灰性紫色土等典型幼年岩性土,层次尚未发育完整,浅薄且肥力不高。原生植物群落(中亚热带常绿阔叶林)由于受长期干扰破坏已不复存在,现有植被常见为柏木、马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)与油松(Pinus tabuliformis)等人工林。柏木人工林林分结构单一且群落构成简单,柏木为其绝对优势种,林下植被较少,成丛明显,灌木主要为铁仔(Myrsine africana)、马桑(Coriaria nepalensis)与火棘(Pyracantha fortuneana)等,草本主要为莎草(Cyrerus microiria)、荩草(Arthraxon hispidus)、圆果雀稗(Paspalum orbiculare)、竹叶草(Oplismenus compositus)与白茅(Imperata cylindrica)等。
1.2 样地设置与野外调查
基于四川省201 个县(局)森林资源二类调查资料(2014 年),2015—2016 年在柏木人工林主产区进行样线踏查并走访林业部门工作人员与国营林场护林员。2017年,为保证采样条件一致性,按照典型抽样与空间代替时间的方法,在各样点选择林木分布较均匀(株行距2 m×3 m或2.5 m×2.5 m)、林相整齐、群落结构相近、经营管理措施相似的柏木人工林为研究对象。共设置标准地70 块(标准地数量依据柏木人工林在我省各区分布面积权重确定),样地面积为400 m2(20 m × 20 m),结合林场经营档案、GPS(Q star 5)及树木生长锥(Haglof CO500)等确定标准地经纬度、海拔、地貌等地形信息及林龄、郁闭度、株高、胸径等林分特征,详见附录Ⅰ。依据国家林业局2003 年与2014 年分别发布的《森林资源规划设计调查主要技术规定》与《国家森林资源连续清查技术规定》对研究区柏木人工林龄组划分:幼龄林≤20 年,中龄林21~40 年,近熟林41~60年。
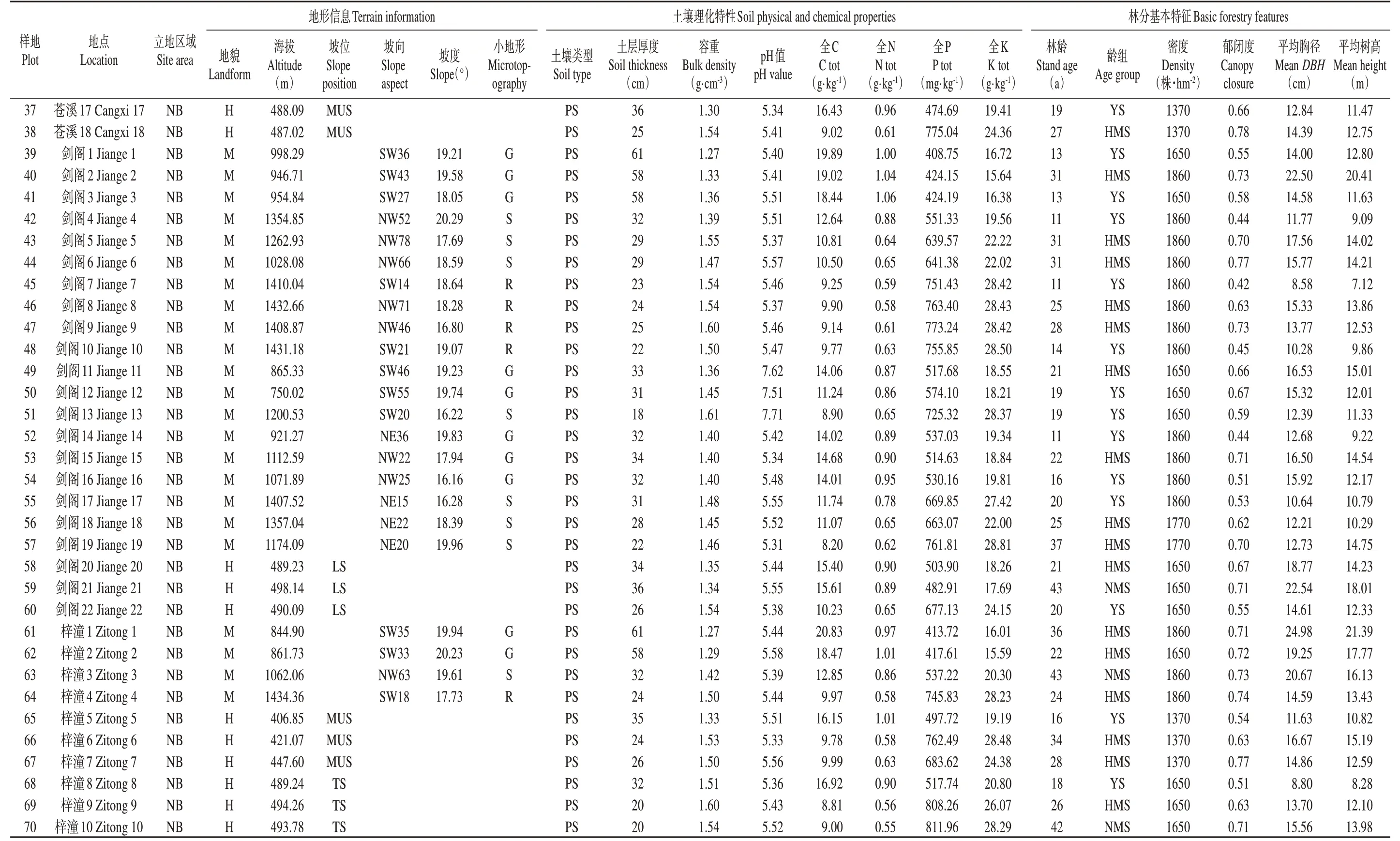
续附录Ⅰ Continued appendix Ⅰ
1.3 样品采集与测定
根据植物生长状况统计结果,在各样地内按对角线法选择长势优良、均匀一致且无损向阳的5株典型标准木,用高枝剪采取树冠东南西北4个方位及上中下3 个部位健康完整的成熟叶片各5 份,每份不少于10 g。将同一样地采集的柏木叶片混匀后放置于装有适量干燥剂的信封袋,做好标记后带回实验室。同时,在各样地内按“S”型路线挖取土壤剖面3 个,测定土壤厚度,用土钻分别采集0~10、10~20 cm 层土样,混合均匀后装入塑料自封袋;用环刀在距土表向下10、20 cm处采取原状土,设置2 次重复。将柏木叶片用蒸馏水清洗去除杂质,经105℃杀青、85℃烘干至恒重后粉碎过筛(100 目)备用;将土壤样品摊晾风干并剔除凋落叶、粗根及土壤动物残体等杂物,研磨过筛(100目)备用。叶样全C 含量测定采用重铬酸钾氧化—外加热法(GB 7657-87),经H2SO4-H2O2消煮后叶全N 和全P 含量测定分别采用半微量凯氏定氮法(LY/T 1269-1999)和钼锑抗比色法(LY/T 1270-1999)。土样水分物理性质测定采用环刀法(LY/T 1215-1999),pH 测定采用电位法(GB 7859-87),土壤全碳、全氮、全磷和全钾含量测定参考鲍士旦[25]主编的《土壤农化分析》,分别使用重铬酸钾氧化—外加热法、凯氏定氮法、钼锑抗比色法和NaOH碱熔—火焰光度法。
1.4 柏木人工林立地类型划分
由柏木实际生长表现及野外调查结果对土层厚度分级:厚层土>60 cm、中层土30~60 cm、薄层土<30 cm,将坡位、坡向、小地形等定性因子分级赋值转为定量因子。根据综合多因素与主导因素相结合的原则,将各变量用R 型聚类降维(度量标准选用pearson 系数,聚类方法选择最远元素,见表1)确定聚类因子后,对研究区柏木人工林进行系统聚类分析和谱系图绘制,事后通过单因素方差分析(one-way ANOVA)和野外经验检验立地分类结果的合理性与有效性[15,20]。

表1 柏木人工林立地类型小区变量相似性矩阵Table 1 Similarity matrix of variable cluster in site type districts of C.funebris plantations
1.5 数据处理
利用Excel 2010 与SPSS Statistics 24.0 软件对数据进行正态分布检验和统计分析,采用Origin 9.0与Canoco for Windows 4.5 软件对结果进行图形绘制。计算各立地与龄组序列柏木人工林叶片化学计量特征的平均值(mean)与标准误差(SE),图表数据均为“平均值(mean)±标准误差(SE)”。利用双因素方差分析法(two-way ANOVA)检验立地、龄组及其交互作用对柏木人工林叶片化学计量特征的影响。利用单因素方差分析法分别比较同一龄组不同立地、同一立地不同龄组的叶片化学计量特征,方差齐性(非齐性)则用LSD 法(Tamhane’s T2
法)进行显著性多重比较。将14 个立地因子与龄组为自变量,柏木叶C、N、P 含量及C∶N、C∶P、N∶P为因变量做PCA 分析,检验环境对柏木叶片化学计量特征相对贡献。
2 结果与分析
2.1 四川省柏木人工林立地分类
结合野外系统调查与立地分类原则,在研究区选择对水热分配占主导的地貌因子来划分丘陵和山地2个立地类型小区;各小区内选择对立地质量有显著影响的土壤类型因子来划分酸性紫色土组和石灰性紫色土组;丘陵各组内因山丘低矮、坡面平缓,故舍弃海拔,选择坡位因子划分坡下部、坡中上部、坡顶台地亚组,山地酸性紫色土组内选择对水热条件再分配的坡向因子来划分阳坡和阴坡亚组。各组/亚组系统聚类时类间距离采用离差平方和方法且以5为阈值进行立地类型划分来保证分类科学性和生产实用性(见图1)。由表2可知,整个研究区柏木人工林被划分为2个立地类型小区,4个立地类型组,8个立地类型亚组和17个立地类型,不同立地的土层厚度、容重和养分含量等各有不同。

表2 四川省柏木人工林立地类型划分结果Table 2 The result of site type classification of C.funebris plantations in Sichuan province
2.2 不同立地类型柏木人工林叶C、N、P含量
由表3 可知,四川省柏木人工林叶C、N、P含量分别为514.59~592.06、9.10~9.92、0.89~1.11 g·kg-1,平均值分别为550.55、9.39、0.95 g·kg-1。从立地类型小区来看(见表3),各龄组柏木叶C 含量均为山地小区显著高于丘陵小区,幼龄林叶N 含量为丘陵小区显著高于山地小区。从立地类型组来看(见图2A~C),丘陵和山地小区内各龄组柏木叶C含量均为酸性紫色土组显著高于石灰性紫色土组,叶N、P 含量均为丘陵石灰性紫色土组最高。从立地类型亚组来看(见图2D~F),各龄组柏木叶C 含量均为丘陵酸性紫色土坡下部亚组和山地酸性紫色土阳坡亚组最高,丘陵石灰性紫色土坡中上部亚组与坡顶台地亚组最低,叶N、P 含量相反。表4 显示,立地类型与龄组对柏木叶C、N、P含量有极显著影响(P<0.001);立地类型×龄组对叶C、N 含量有极显著影响(P<0.001),对叶P 含量有显著影响(P<0.05)。柏木叶C、N 和P 含量在不同立地类型中最高值是最低值的1.23、1.08 和1.23 倍(见表5)。叶C 含量在幼龄林与中龄林中均为k、n和p立地类型内较高,在近熟林中为k、p 和b 立地类型内较高;叶N、P 含量在各龄组中均为i、j 和e 立地类型内较高,表明柏木叶C 高积累量集中出现在丘陵厚土层或山地沟槽中,叶N、P反之。

表4 立地类型和龄组对柏木人工林叶C、N、P含量及化学计量比影响的双因素方差分析Table 4 Two-way ANOVA for the site type and age group on leaf C,N,P and the stoichiometry of C.funebris plantations
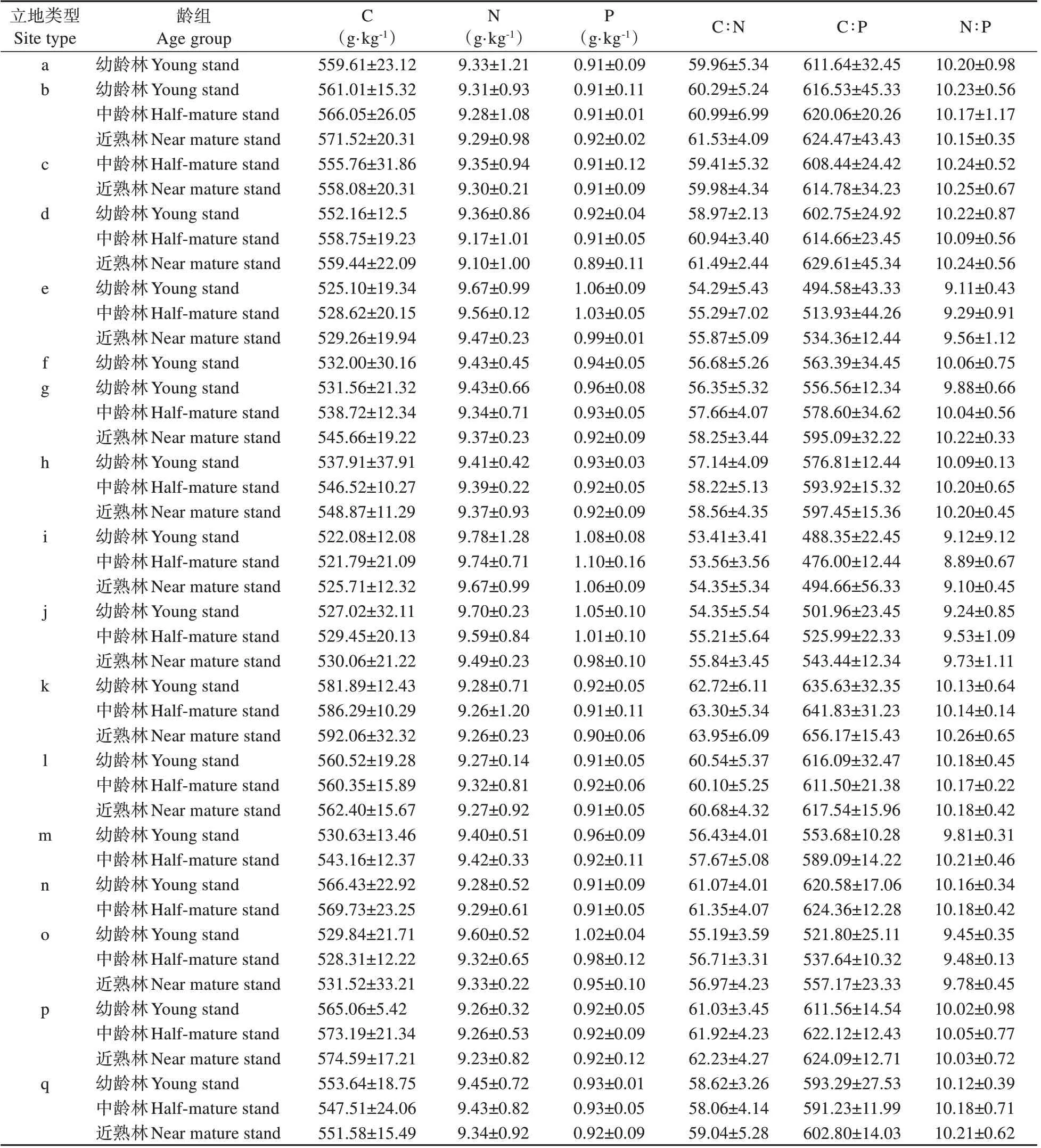
表5 不同立地类型间柏木人工林叶C、N、P含量及化学计量比Table 5 The leaf C,N,P contents and the stoichiometry of C.funebris plantations in different site types

表3 不同立地类型小区间柏木人工林叶C、N、P含量及化学计量比Table 3 The leaf C,N,P contents and the stoichiometry of C.funebris plantations in different site type districts
2.3 不同立地类型柏木人工林叶C∶N、C∶P、N∶P
由表3 可知,四川省柏木人工林叶C∶N、C∶P、N∶P 分 别 为52.18~63.95、465.94~656.17、8.78~10.30,平均值分别为58.70、583.68、9.93。从立地类型小区来看,各龄组柏木叶C∶N 和C∶P 均为山地小区显著高于丘陵小区。从立地类型组来看(见图2G~I),丘陵和山地小区内各龄组柏木叶C∶N 和C∶P 均为酸性紫色土组显著高于石灰性紫色土组,叶N∶P 为丘陵石灰性紫色土组最低;从立地类型亚组来看(见图2J~L),各龄组柏木叶C∶N、C∶P和N∶P均为丘陵酸性紫色土坡下部/中上部亚组和山地酸性紫色土阳坡亚组最高,在丘陵石灰性紫色土坡中上部亚组最低。立地类型与龄组及立地类型×龄组对柏木叶C∶N 和C∶P 均有极显著影响(P<0.01),立地类型与龄组对叶N∶P 均有显著影响(P<0.05)。柏木叶C∶N、C∶P 和N∶P 在不同立地类型中最高值是最低值的1.20、1.34 和1.13 倍(见表4~5)。叶C∶N和C∶P在幼龄林与中龄林均为k、p 和n 立地类型内较高,在近熟林均为k、p 和b 立地类型内较高;叶N∶P 在幼龄林为b、d 和a 立地类型内较高,在中龄林为c、m 和h 立地类型较高,在近熟林为k、c 和d 立地类型较高。C∶N、C∶P 可反映植物对生境中N、P 的需求利用水平,结果表明柏木叶在丘陵厚土层与山地沟槽中N、P 需求利用水平较高而在浅薄的坡中上部需求利用水平较低。随龄组增大,各立地类型组/亚组中柏木叶C含量、C∶N 和C∶P 差异显著且整体上升;叶N 和P含量普遍下降,幼龄林至中龄林变幅更大而后变幅减缓;N∶P 差异不显著但整体呈现缓慢上升的变化趋势。
2.4 环境因子对柏木人工林生态化学计量特征影响的PCA分析
对14 种立地因子及龄组与柏木人工林叶C、N、P 含量及C∶N、C∶P、N∶P 进行主成分分析,结果经Monte Carfo Test 检验。第1 排序轴和其余所有排序轴均显著(F=2.532,P=0.029;F=2.157,P=0.033),且前两个排序轴对柏木叶化学计量值的累积贡献率达95.5%,表征其可在绝大程度上反映柏木叶C、N、P含量及比值与环境之间的关系。其中第1 轴与土壤类型、坡位、pH、容重、土壤全磷及全钾呈显著正相关,与地貌、坡向、小地形、土层厚度、土壤全碳和全氮及龄组呈显著负相关;第2 轴与地貌、坡向、土层厚度、土壤全碳和全氮及龄组呈显著正相关,与土壤类型、坡位、小地形、pH、容重、土壤全磷和全钾呈显著负相关(见表6)。由图3 可知坡位除对柏木叶P 含量和N∶P 无显著作用外,对其他特征值影响较大,尤其是叶C含量和C∶N;其余环境因子对柏木叶片化学计量特征均有显著影响。
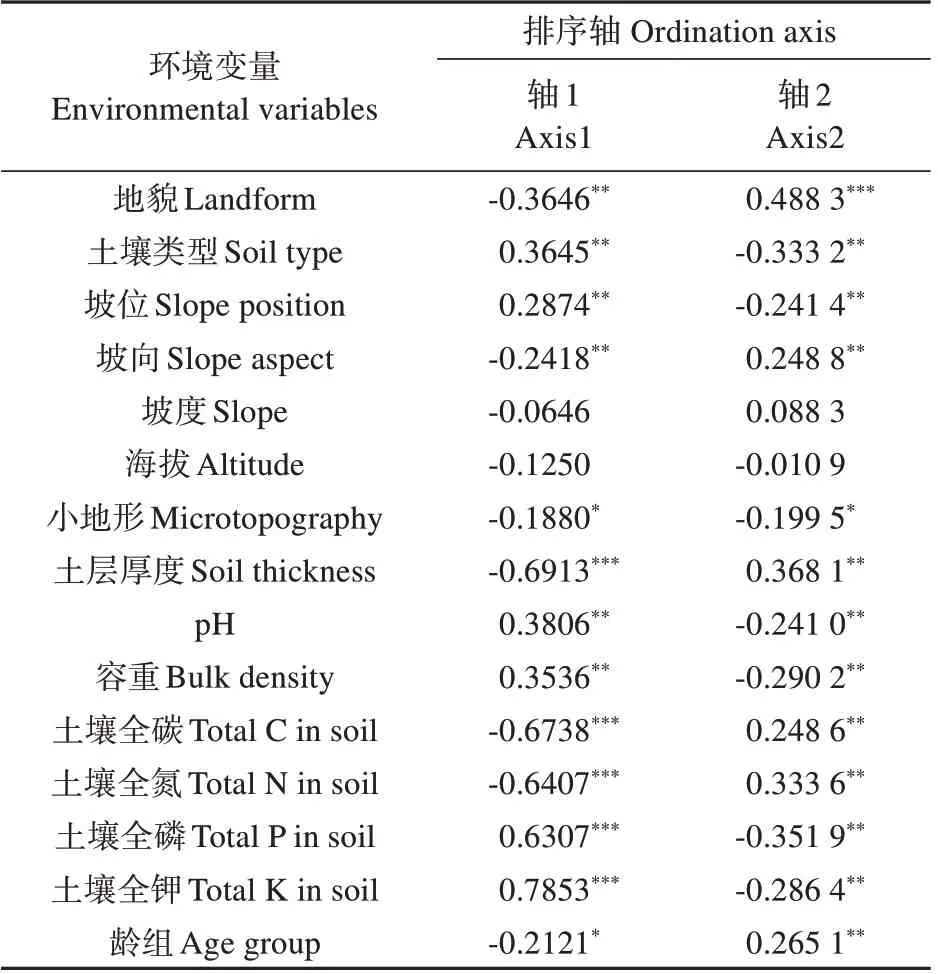
表6 环境变量与排序轴的相关系数Table 6 Correlation coefficients of environmental variables with ordination axes
3 讨论
3.1 四川省柏木人工林叶C、N、P 含量及比值特征
植物主要组成元素C、N、P及其比值能反映外部营养条件供应状况,表征个体对环境资源的需求水平[11]。四川省柏木人工林平均叶C 含量为550.55±18.08 g·kg-1,高 于 全 球492 种 陆 地 植 物(464.00±32.10 g·kg-1)[26]和中国东部南北样带102个优势种(480.1±25.3 g·kg-1)[27]叶片C 含量均值,说明其具有较高的C 积累水平与干物质合成能力。柏木人工林平均叶N、P 含量为9.39±0.20、0.95±0.12 g·kg-1,均低于全球(18.30、1.99 g·kg-1)[26]和全国(17.55、1.28 g·kg-1)[28]植物叶片N、P 含量,可能暗示了研究区柏木人工林生境中可获得性N、P 元素含量较低。一方面,紫色土本就含N 较少,加之植物对N 的吸收利用主要为可溶性硝态氮和铵态氮[12,26],容易在四川充沛的降雨量下发生淋溶流失;另一方面,研究区部分土壤酸化加速了Al 的原生/次生矿物风化[10~13],释放出大量Al3+,富铝化作用使土壤中Fe、Al活性升高,与P 结合形成难溶性Fe-P 和Al-P 等固定态P 化合物[27],化学性质稳定,难以被柏木利用,验证了我国亚热带土壤缺P 的普遍特征[28]。再者,就植物自身而言,柏木为常绿树种,相较阔叶树,常绿树叶寿命较长,凋落量小,可有效减少养分损失[11],故较低的营养需求使得柏木叶N、P吸收蓄积水平相对较低。
四川省柏木人工林平均叶C∶N、C∶P为58.70±3.04、583.68±44.59,远高于全球(23.80、300.90)[26]与中国森林生态系统(37.10、469.20)[28]植物的平均水平。柏木叶较高的C∶N、C∶P 是其高C 含量低N、P 含量的结果,也是常绿鳞形叶、针叶等窄叶树种富C 次生产物(蜡、树脂、单宁等)占比较高的直接体现[12,29~30]。N、P 养分的获得与利用常受自然条件调控,易成为陆地生态系统中的限制性元素,故人们常将N∶P 作为外界环境对植物养分供应的指示标准[11,13,31]。已有研究表明N∶P 大于16 时P元素为受限位点,小于14时N元素为受限位点[32]。本研究中四川省柏木人工林平均叶N∶P 为9.93±0.37,略低于全球森林生态系统植物N∶P平均水平(12.60)[26],远低于14,说明该地区柏木可能受N限制。研究区位于我国高N 沉降区域(华西雨屏区),一些原本受N 限制的物种出现“N 饱和现象”转为受P 或其他元素限制,但柏木仍受N 限制,可能是因为这种持续性N 输入反而引起部分地区“N过饱和”,生态系统固N 能力下降,输入N 量不及损失N量[33]。
3.2 立地类型对柏木人工林叶C、N、P 含量及比值的影响
植物生长发育与生境条件密切相关,有机体可通过调节C、N、P 的代谢和循环,尤其是计量比值格局,来反映其对环境变化的响应和适应性[30,32]。N∶P可表征植物受限状况;C∶N、C∶P可表征植物对C 的同化能力与对N、P 的利用效率,一般较高的C∶N、C∶P 对应较高的养分利用效率[28~29,31]。本研究发现,四川省柏木人工林叶C 含量与C∶N、C∶P在山地小区显著高于丘陵小区。环境可决定植物的化学组成,人类活动又可改变环境条件来间接影响植物C、N、P分布[10]。丘陵小区柏木林多由荒坡造林形成,位于农田、村庄等人口稠密区,由于频繁受多种人为活动干扰,生境N、P供应量被影响的同时树体C、N、P生物平衡受到破坏[8~9],林木积累的有机质和养分的流失(90%以上)远大于对生态系统的归还(10%以下)。且由于林下植被的破坏加剧了土壤侵蚀与养分流失,土壤肥力下降,特别是N、P 有效供应水平大幅下降[12,24,27],也会造成生境养分周转效率较人类干扰程度轻、植被保存相对完整的山地小区更低。就立地类型组而言,柏木人工林叶C含量与C∶N、C∶P在各小区内均为酸性紫色土组显著高于石灰性紫色土组。土壤酸碱度影响着各种元素有效性[30],柏木虽具有较强耐贫瘠耐干旱的能力,对钙质、石灰土适应力较强,但在酸性淋溶紫色土/暗紫色土中生长最佳。
就立地类型亚组而言,柏木人工林叶C 含量与C∶N、C∶P在丘陵小区各组内均为坡下部亚组显著高于坡中上部和坡顶台地亚组,在山地小区酸性紫色土组内为阳坡亚组显著高于阴坡亚组。表2、图2D,J,K 及PCA 分析可知坡位越靠下,土层越深厚,柏木叶C 积累量和N、P 利用效率越高。不同坡位由于受到地面起伏、人类干扰程度与风、水蚀等影响,表层土理化性质与局域微环境有所不同[23~24]。丘陵坡顶台地受人为干扰严重;坡中上部养分在被土壤固定前易在淋溶作用下渗透流失,在被土壤固定后易沿坡面径流冲刷流失[29],导致坡中上部柏木N 受限状况最严重;而坡下部(下垫面)由于在长期侵蚀过程中对岩石碎块碎屑等有截留固定的作用[21~23],使得其土层深厚,物质充分,养分相对富积,微生物总数最多、状况最好[24,31],更有利于柏木生长发育和对养分吸收利用。坡向影响光热与降水的分配,不同光照条件下柏木叶光合作用及养分代谢速率不同,对土壤矿物质元素的获得能力和利用效率也不同[22]。N是决定光合物质代谢能力的最关键因素,P是核糖RNA 的必需元素,植物在充足的光照资源中为满足自身生长所需能量,会增加对这两种元素的需求及利用效率[26]。阳坡太阳辐射均匀充足,柏木光合作用更强烈,加快N、P 周转速率的同时也加速对C 的合成与积累[12,24]。并且阳坡较阴坡的土温、气温更高,可能会影响喜光喜阳的柏木酶类活性、反应介质和膜系统流动性等,从而调节元素利用水平[14,28]。干热气候条件下柏木根系还会产生更多分泌物以提高根际螯合作用[12],进而提高其对土壤N、P 元素的选择性吸收利用效率。因此,在阳坡中柏木叶C 含量与C∶N、C∶P 更高。就立地类型而言,柏木人工林叶C 含量与C∶N、C∶P 在厚土层及沟槽类型显著高于中土层及坡地、脊梁类型,N、P 含量反之。表明厚土层及沟槽类型中柏木叶片吸收N、P 后利用效率较高,产出有机物质多。沟槽土层较厚,养分充足,特别是微生物养分周转条件与N 素供应容量良好[18],可促进柏木健康生长。
3.3 龄组对柏木人工林叶C、N、P 含量及比值的影响
植物在不同生长发育阶段与环境资源“供应—需求”间存在错配现象以调节其生理生态过程,可通过改变自身C∶N∶P来适应器官发育速率及生境条件的变化[26,34]。本研究发现,不同龄组间柏木人工林叶C含量、C∶N和C∶P差异显著且从幼龄林组到近熟林组整体呈不断上升趋势;N、P 含量普遍不断下降,幼龄林组至中龄林组变幅更大而后变幅减缓;N∶P 差异不显著但整体呈现缓慢上升的变化趋势。光合作用是植物获取能量和增长干物质的关键途径[28~29],柏木自幼龄林阶段进入相对速生期,光合效率提高,干物质合成量增大,C含量升高,快速消耗N、P 养分以健全营养器官发育,加速生命活动,因此初期的快速生长引起N、P 含量下降(尤其是N 含量),而C∶N 和C∶P 不断升高,且幼龄林至中龄林阶段更明显,Elser 等[32]也曾提出植物早期生长易受有效性N 限制的结论。进入近熟林后柏木代谢与合成速度变缓,N、P 需求有所下降,富C 结构性物质的提高使得C∶N、C∶P 持续上升。受造林时间影响,目前研究区极少存在林型与经营管理措施等方面与其他龄组实验样地相对一致的柏木成熟林标准地,故龄组对柏木叶化学计量的影响有待后期增加成熟林组的实验以持续深入分析。此外,本研究发现柏木人工林叶N、P 含量对龄组变化均有显著响应,而N∶P 在立地类型与龄组的交互影响下却保持着相对稳定,反映了植物能通过对自身C∶N∶P比例的调节来与其环境养分供应之间保持一种相对稳定的“元素动态平衡”格局[28,30]。
4 结论
立地类型与龄组显著影响四川省柏木人工林叶片生态化学计量特征。柏木人工林在山地阳坡沟槽酸性紫色土,或丘陵坡下部较深厚的酸性紫色土地区有较高的有机物合成积累能力及N、P 利用效率,而丘陵石灰性紫色土、浅薄的坡中上部酸性紫色土及山地阴坡坡面C积累不高,N、P相对稀缺。从幼龄林到近熟林阶段的各立地类型中柏木人工林叶C 含量、C∶N 和C∶P 不断上升,N、P 含量不断下降,受N 限制严重且生长前期更明显。为促使四川省柏木人工林健康生长并有更好的化学计量分配格局,建议减少人为干扰,自幼龄林时期施N 肥或混交有效N 存储中心的豆科(Leguminosae)植物,通过与根瘤菌共生固氮来提高系统N∶P水平、有效N 元素和土壤肥力,促进林地养分循环。
