彭泽鲫♀×兴国红鲤♂四倍体子代与彭泽鲫子代生长性能比较
2021-02-10张庆飞操文杰廖寿力魏怡飞王卫民
张庆飞,操文杰,廖寿力,魏怡飞,王卫民
华中农业大学水产学院/农业动物遗传育种与繁育教育部重点实验室/农业农村部淡水生物繁育重点实验室, 武汉 430070
彭泽鲫(Carassiusauratuspengsenensis)是自1983年选育7代得到的具有抗病和抗逆能力强、生长速度快、营养价值高的经济养殖品种,属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲫属(Carassius)。兴国红鲤(Cyprinuscarpiouarsinguonensis)属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲤属(Cyprinus),是“江西三红”之一。由于兴国红鲤食性广、抗逆、抗病性强、杂交亲和度高[1],且易于与鲫进行杂交产生具有杂种优势的新品种,因而常用作杂交亲本[2]。杂交育种是培育新品种常用的方式,杂交子代的生长性能也是养殖界关注的重点问题[3]。
现阶段鲫种类繁多,在全国各地均有养殖,但经年累代养殖、近亲交配、养殖环境恶化造成种质资源退化、抗病抗逆性衰退、生长速度减慢等多种问题,新品种的开发和培育势在必行[4]。已有研究表明,不同种类鲫的生长性能具有明显差异,杂交品种鲫的生长速度通常更快[5]。由于彭泽鲫是公认的三倍体,且生殖方式为天然雌核发育[6],杂交育种的方式并不适用。但近年来有研究者发现利用不同种类的精子刺激三倍体银鲫(两性型雌核发育)、彭泽鲫雌核发育也会对子代生长性能等造成影响[7]。笔者所在课题组前期用4种精子刺激彭泽鲫雌核发育,结果在兴国红鲤精子刺激彭泽鲫雌核发育的子代中检测到了极少数四倍体,经过产前培育进一步繁殖得到子代。经检测,其子代均为四倍体。我们认为该四倍体发生了精卵结合,即发生了远缘杂交,并且制作了染色体核型模式图、扩增其5S rDNA序列证明其为杂交品种(另文发表)。通常情况下,远缘杂交会结合亲本的优良性状,表现出杂种优势[8],育种潜力较高,但是否能形成自交繁衍的两性可育品种仍需对其进行进一步研究。
本研究以彭泽鲫♀×兴国红鲤♂四倍体子代和彭泽鲫子代为研究对象,通过180 d的饲养试验,分析2种子代生长性能的差异,旨在为彭泽鲫新品种的选育、可持续健康养殖等相关研究提供理论依据。
1 材料与方法
1.1 试验鱼
将笔者所在课题组前期在兴国红鲤刺激彭泽鲫雌核发育的子代中发现的四倍体,经过产前培育,于2020年5月进行人工繁殖得到四倍体子代;彭泽鲫亲本取自江西省水产科学研究所,于同一时期进行人工繁殖得到彭泽鲫子代。
1.2 试验设计与饲养管理
2020年5月挑选健康无病且性腺发育良好的2种亲本,使用地欧酮和促黄体素释放激素类似物A2进行催产并进行人工授精得到受精卵,置于孵化池孵化。待鱼苗孵出可以平游时,转移至水泥池暂养。于2020年8月9日取体型大小相似的经过倍性鉴定的彭泽鲫♀×兴国红鲤♂四倍体子代以及彭泽鲫雌核发育子代各300尾,分为3个平行组,每组100尾,置于华中农业大学南湖水产养殖基地1 m3圆桶内饲养。试验周期180 d,每个平行组均进行标准化养殖,进行定质、定量、定时和定位投喂,且定期吸污换水。
1.3 四倍体子代倍性鉴定
以彭泽鲫子代为内参,通过流式细胞仪检测四倍体子代倍性。分别取抗凝血剂10 μL置于2 mL离心管中,加入500 μL DAPI染液,再加入500 μL PBS缓冲液,用48 μm滤膜过滤,避光染色5 min后,上机检测。
1.4 数据收集与统计分析
在试验0、30、60、90、120、150、180 d时,各平行组随机挑选20尾鱼进行体质量、体长数据测量,根据公式计算绝对增重率、绝对增长率、增积量和肥满度:
绝对增重率(g/d)=(W2-W1)/(t2-t1)
绝对增长率(cm/d)=(L2-L1)/(t2-t1)
增积量(g·cm/d)=(W2-W1)(L2-L1)/(t2-t1)2
肥满度(g/cm3)=W/L3×100
式中,W与L分别代表体质量(g)与体长(cm),W1、W2和L1、L2分别为时间t1、t2时的体质量和体长。
此外,拟合2种子代体质量与体长的幂函数曲线关系(W=aLb),其中,参数a为常数,表示生长条件因子;b为幂指数系数。
所有数据均以“平均值±标准差”表示,数据采用Excel 2016与SPSS 22.0软件分析,采用独立样本t检验分析差异显著性,以α=0.05作为差异显著水平。
2 结果与分析
2.1 子代倍性鉴定结果
利用流式细胞仪检测血液混样所得结果见图1。横坐标表示红细胞荧光值,纵坐标表示细胞数目。彭泽鲫雌核发育子代平均荧光值为305.24±3.42,杂交四倍体子代荧光值为403.79±2.18,二者DNA相对含量比为0.76,即四倍体子代仍为四倍体。
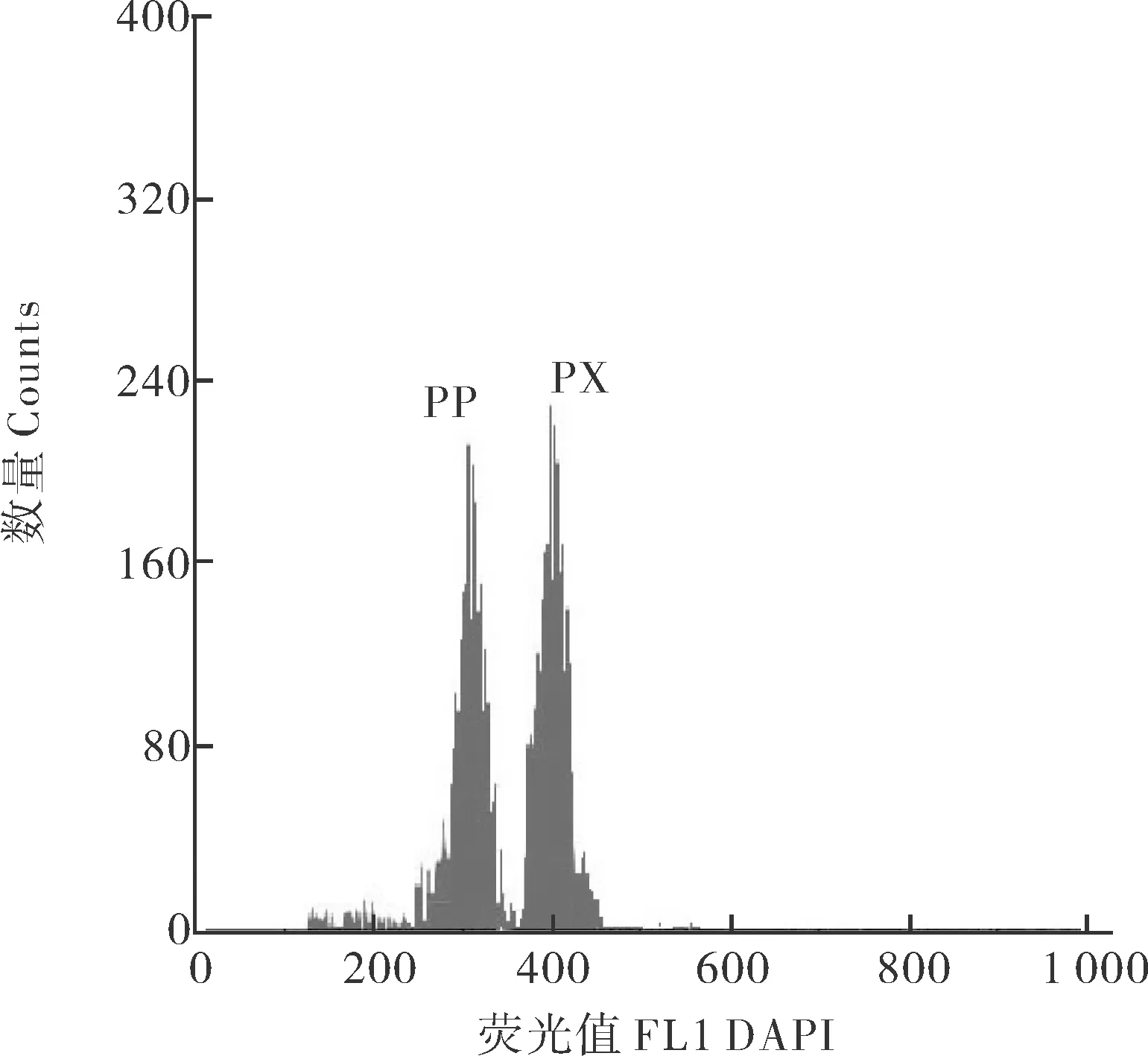
PP为彭泽鲫雌核发育子代 Offspring of Carassius auratus pengsenensis; PX为彭泽鲫与兴国红鲤杂交四倍体子代 Offspring of the tetraploid.
2.2 体质量、体长数据
彭泽鲫♀×兴国红鲤♂四倍体子代和彭泽鲫雌核发育子代平均体质量、平均体长数据见表1。结果显示,试验开始时二者体质量平均值分别为3.08±0.33、3.19±0.35 g,体长平均值分别为4.61±0.16、4.69±0.15 cm,均无显著性差异。但在随后的180 d饲养过程中,彭泽鲫♀×兴国红鲤♂四倍体子代体质量、体长均与彭泽鲫子代有极显著差异。在180 d时,四倍体子代体质量、体长分别高出彭泽鲫子代30.18%、7.70%,表明彭泽鲫♀×兴国红鲤♂四倍体子代具有明显的生长优势。
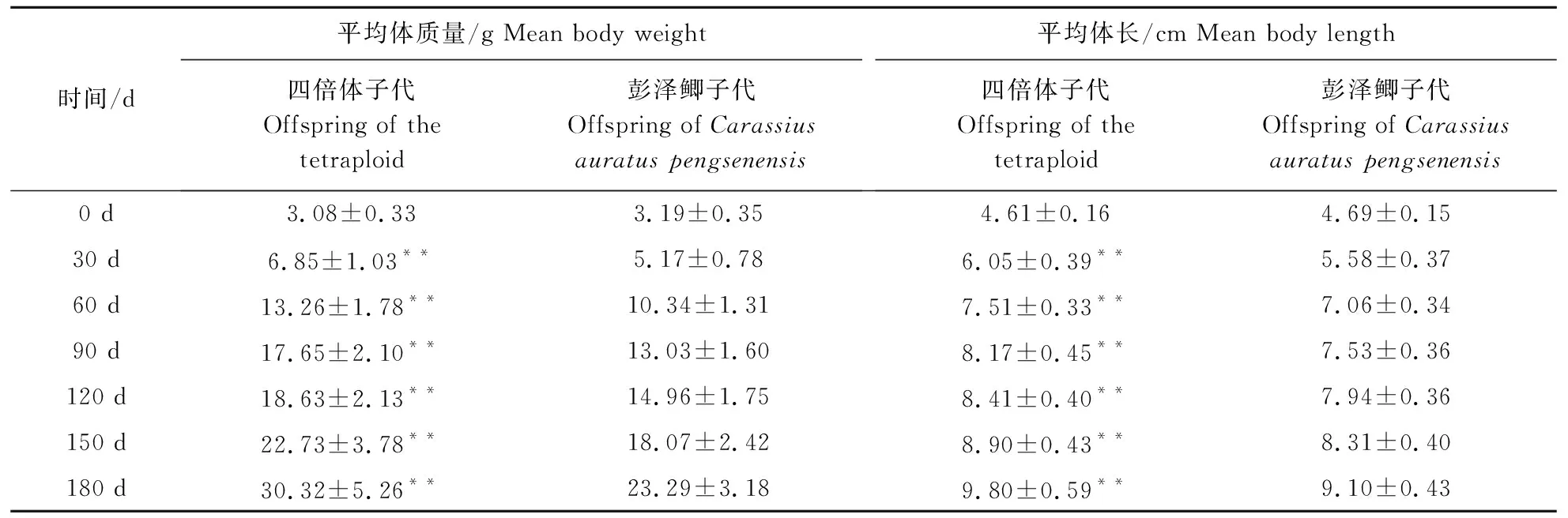
表1 2种子代各时期平均体质量、平均体长 Table 1 Average body weight and body length of two generations at different periods
2.3 绝对增重率、绝对增长率比较
彭泽鲫♀×兴国红鲤♂四倍体子代及彭泽鲫雌核发育子代各时期绝对增重率和绝对增长率结果如图2所示。2种子代绝对增重率均呈现先升高、再降低、再升高的趋势,在90~120 d时降到最小值。此外,在0~90、150~180 d时,四倍体子代极显著高于彭泽鲫子代;120~150 d时显著高于彭泽鲫子代。此外,绝对增长率也呈现出先增高、再降低、再升高的趋势,且90~120 d时彭泽鲫子代绝对增长率高于四倍体子代。二者的绝对增长率仅在0~30 d、60~90 d时表现出差异,30~60 d达到最高值,90~180 d过程中均无显著差异。
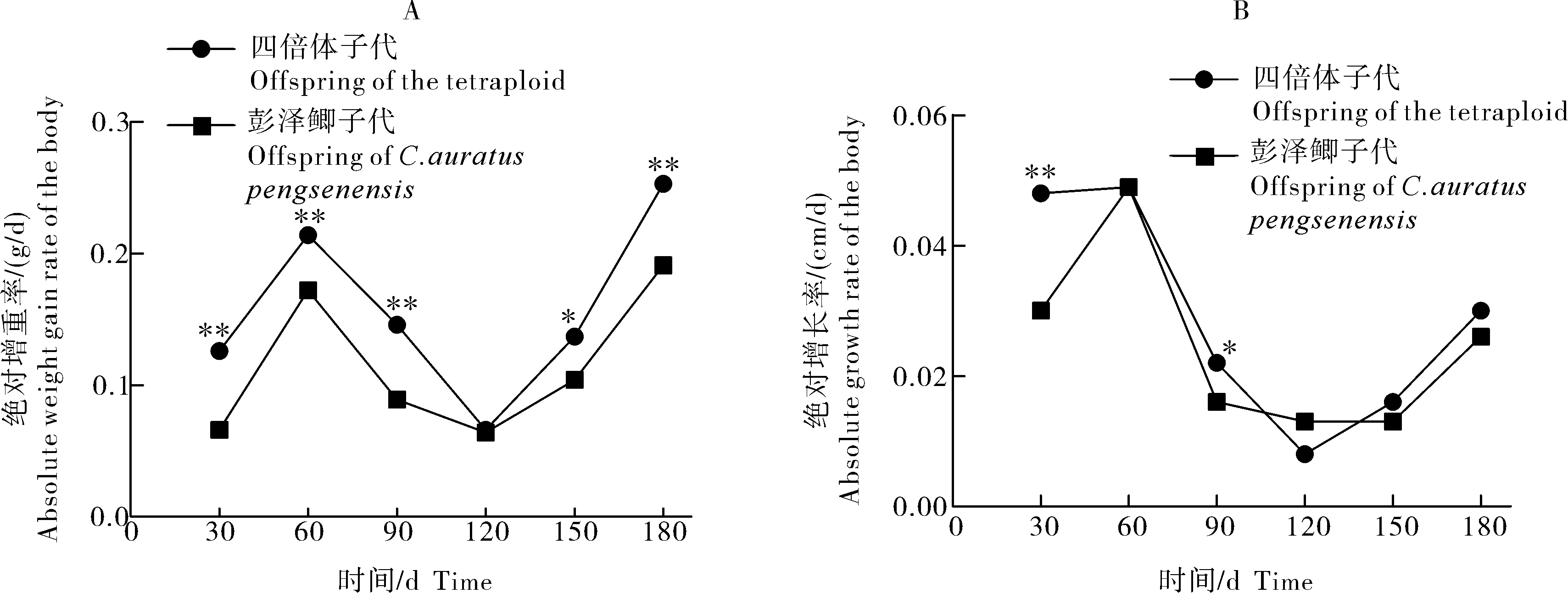
A:绝对增重率;B:绝对增长率。*代表差异显著(P<0.05);**代表差异极显著(P<0.01)。A:Absolute weight gain rate of the body;B:Absolute growth rate of the body. *represents significant difference (P<0.05); **represents extremely significant difference (P<0.01).
2.4 增积量及肥满度
彭泽鲫♀×兴国红鲤♂四倍体子代和彭泽鲫雌核发育子代各时期增积量和肥满度如表2所示。由表2可见:在180 d饲养过程中,增积量也呈现先升高、再降低、后升高的趋势,这与绝对增重率、绝对增长率的趋势相同;在60 d时,2种子代增积量达到最大值,分别为0.010 5±0.001 2、0.008 6±0.001 7 (g·cm)/d;且0~30 d、60~90 d,四倍体子代增积量极显著高于彭泽鲫子代;30~60 d、150~180 d四倍体子代增积量显著高于彭泽鲫子代。在0~30 d,二者肥满度无显著差异(P>0.05),即二者体型较为相似;30~180 d期间,四倍体子代肥满度与彭泽鲫子代有极显著差异,表明四倍体子代较为肥胖;在0~180 d期间,四倍体子代与彭泽鲫子代肥满度变化不大,最大值分别为3.277 7±0.078 3、3.125 6±0.038 5 g/cm3。
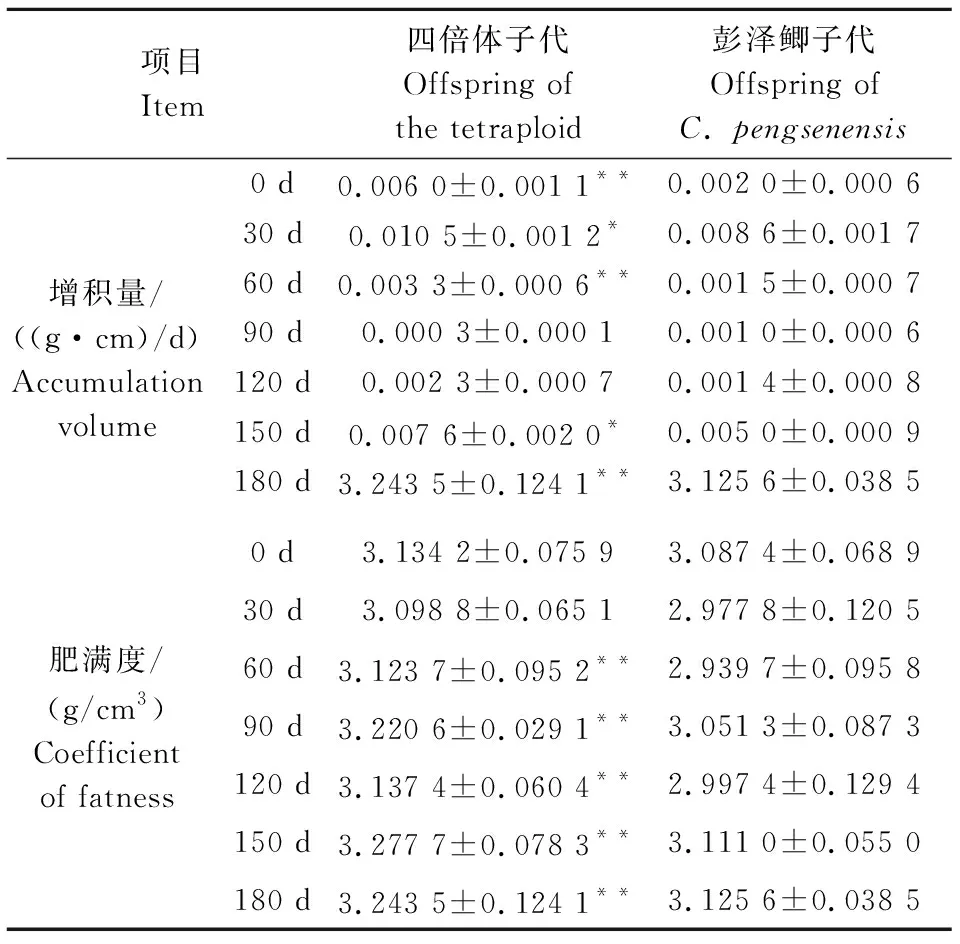
表2 2种子代各时期增积量和肥满度 Table 2 Accumulation volume and coefficient of fatness
2.5 体质量与体长关系
通过180 d体质量、体长数据的收集,拟合二者生长曲线见图3。生长方程分别为彭泽鲫♀×兴国红鲤♂杂交四倍体子代W=0.0320L2.9915(R2=0.977 9);彭泽鲫自交子代W=0.0307L2.9941(R2=0.982 7)。
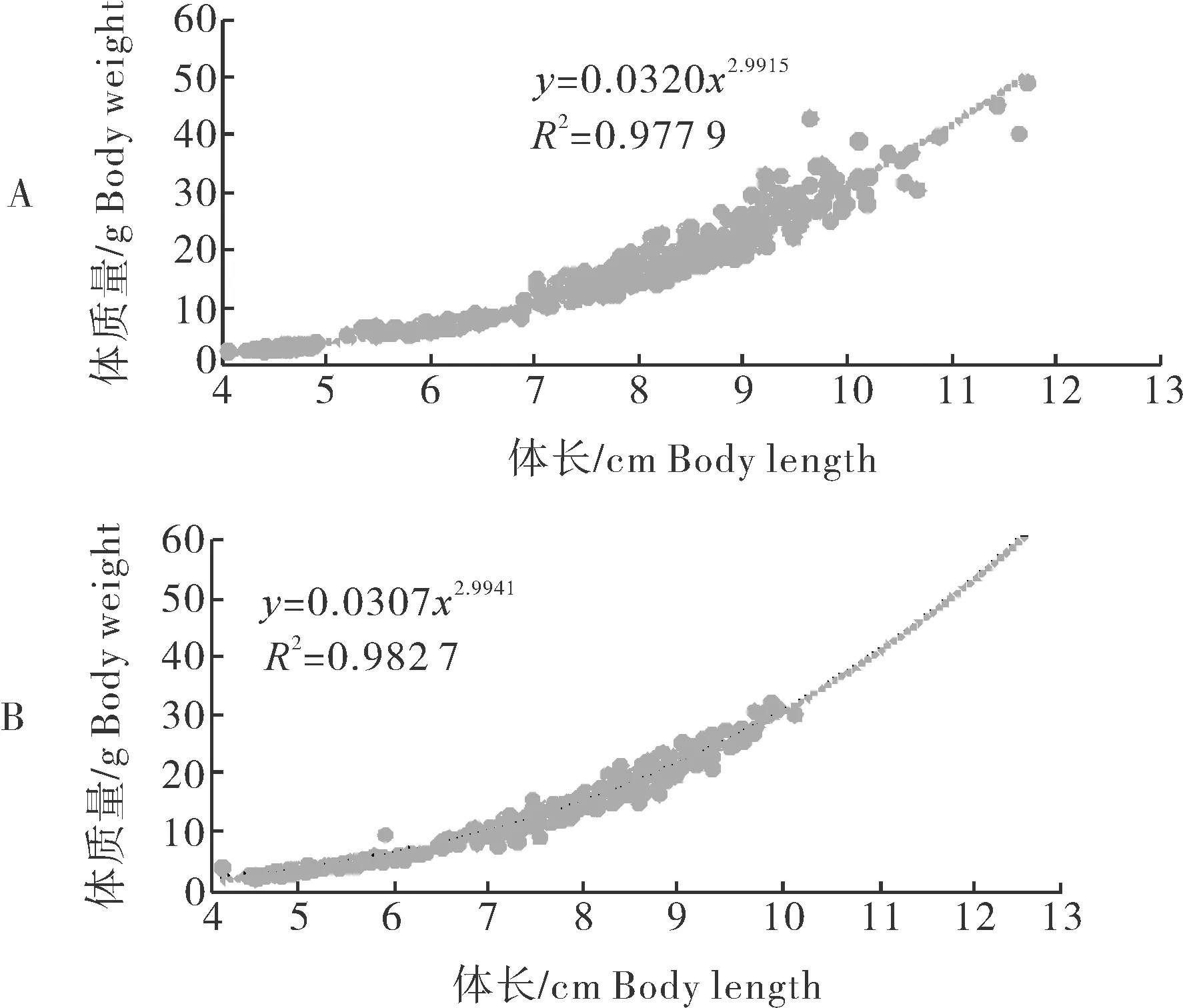
A:杂交四倍体子代; B:彭泽鲫自交子代。A:Offspring of the tetraploid;B:Offspring of Cauratus pengsenensis.
3 讨 论
通常情况下,杂交子代均能表现出一定的生长优势[9-10]。在本试验中,笔者在兴国红鲤精子刺激彭泽鲫雌核发育的子代中发现了四倍体,进一步人工繁殖得到其子代。为了评估其生长性能,以彭泽鲫自交子代作为对照,分析2种子代早期生长性能差异,为彭泽鲫新品种开发提供数据支持。在本试验中,初始体质量、体长会对结果造成一定影响。为了消除这一因素造成的误差,本试验尽量挑选体型、规格整齐的育苗,对初始数据进行差异性分析,结果显示二者差异不显著,影响较小。
二者绝对增重率、绝对增长率呈现相同趋势,原因主要是温度变化。有研究表明鱼类的生长速度与其能量吸收及转化有关,水温可以通过影响酶活进而影响鱼类的新陈代谢,导致生长速率的不同[11]。本试验中60~120 d时处于10-11月,水温降低,鱼类摄食减慢,生长代谢缓慢,导致鱼类生长缓慢。待加设加热棒以后,摄食恢复,绝对增重率、绝对增长率又逐渐升高。这一结果与周淼等[12]研究结果一致。
增积量表示鱼类体质量日增长量与体长日增长量之积[13],能够反映鱼类的增长速度。通过比较2种子代各时期增积量发现:增积量变化趋势与绝对增重率相似,均呈现先升高、再降低、后升高的趋势,且在60 d时二者增积量达到最高值且差异显著(P<0.05),这些现象都是由水温变化进而改变鱼类摄食所引起的,与绝对增重率、绝对增长率结果一致。
肥满度是衡量鱼类肥满程度的指标,通常情况下可反映鱼类可食程度,此外肥满度也可从侧面反映鱼类的营养状况与生长环境条件[14]。在本试验中,由于2种子代并未达到性成熟,因而性腺发育对肥满度的影响较小,结果较为可靠。在30 d之前,二者肥满度差异不显著,但在30~180 d时,四倍体子代肥满度极显著高于彭泽鲫子代,这说明四倍体子代摄食较多,鱼体可食部分更多、营养更丰富。由以上结果也推测在30~60 d内,四倍体子代进入快速生长阶段。
通常情况下,硬骨鱼类的体质量与体长符合幂函数关系(W=aLb),我们称之为生长曲线,它能够准确地描述鱼类的生长过程。其中R2值越接近1表示曲线拟合程度越好,b值反映鱼类生长发育的不均匀性。此外,有研究者认为b值也能反映鱼类生长环境和营养吸收的好坏,以及生长的阶段性[15-16]。体质量与体长的关系伴随着鱼类的生长而改变,在幼鱼阶段呈负异速生长,随着鱼体的生长速度趋于平缓,b值逐渐接近3。例如,七带石斑鱼在仔稚幼鱼阶段,其体质量与体长的关系式为W=1×10-5L3.5345(R2=0.981 9),而在10~14月龄阶段其生长曲线关系为W=0.436L2.055(R2=0.959)[17]。此外,王雪辉等[18]通过统计4目16科25属共计43种鱼的生长曲线发现同一阶段不同种类的鱼生长关系有差异;同种鱼在不同阶段体质量与体长关系也有差异。另有研究表明同一物种不同群体的鱼类在相同发育阶段其体质量与体长关系相似,如1~10月龄养殖大黄鱼(Larimichthyscrocea)F2体质量与体长的关系式为W=0.0569L3.0496(R2=0.998 9),野外捕捞大黄鱼F1的关系式为W=0.0565L3.0365(R2= 0.998 4)[19];周淼等[12]在彭泽鲫异精雌核发育子代中也有相同结果。本试验中2种子代R2值分别为0.977 9和0.982 7,接近1,曲线拟合度较高;b值分别为2.991 5和2.994 1,均接近3.0,呈现等速生长,符合von Bertalanffy生长方程。由此可知,2种子代的生长差异不是由于外界环境条件和投喂策略等人为因素引起的,主要原因是2种子代能量吸收和转化之间的差异。这与刘尊雷等[20]对小黄鱼生长模型研究中的结果一致。此外,生长曲线通常呈“S”形,具有2个拐点,即经过第一拐点后生长速度急剧加快,第二拐点后生长速度变缓慢,通常第二拐点在性腺发育阶段[21]。由于本试验中2种子代均未达到性成熟,根据绝对增长率和绝对增重率以及增积量、肥满度的变化,我们推测2种子代第一个拐点可能在30~60 d内。
在本研究的180 d养殖过程中,彭泽鲫♀×兴国红鲤♂四倍体子代相较于彭泽鲫雌核发育子代表现出较好的生长优势,体质量、体长的增长均极显著高于彭泽鲫雌核发育子代,这可为彭泽鲫新品种的培育提供数据支持。有关彭泽鲫♀×兴国红鲤♂四倍体子代在形态、抗病性、抗逆性等优势有待进一步研究。
