冷驯化期间4种楸树叶片叶绿素荧光参数变化研究
2021-02-03魏兰波董佳慧钱稷邸葆
魏兰波,董佳慧,钱稷,邸葆
(河北农业大学 园艺学院, 河北 保定 071001)
光合作用对低温的反应十分敏感,低温胁迫会使叶绿体基质片层松散,类囊体膨大,造成叶绿体双层膜完整性丧失,最终表现为叶绿素含量下降[1]。相关研究表明[2-4],叶绿素含量的高低与园林树木的抗寒性具有明显的相关性,抗寒性强的植物叶片叶绿素含量高于抗寒性弱的植物叶片叶绿素含量,而较高的叶绿素含量有利于产生可溶性糖等渗透调节物质,进而降低细胞冰点,增强植株的抗寒性[5]。当叶片吸收过多的光能时,可以启动自身保护机制,通过非光化学猝灭等形式耗散掉过多的光能,但当不能及时有效的加以利用或耗散时,植物就会受到强光的胁迫,发生光抑制现象。低温可以增加发生光抑制的可能性,主要原因是低温抑制了受损D1蛋白降解酶等相关的酶活性,严重抑制了光系统的修复。有的学者提出,叶绿素荧光参数与低温胁迫密切相关,其可用于植物抗寒性鉴定,其中Fv/Fm可作为植物抗寒性的鉴定指标之一,可扩展应用于叶片之外的其他绿色组织器官[6-7]。
楸树(Catalpaspp.)是我国所特有的乡土树种,其不仅是珍贵的用材树种,也是主要的园林绿化树种。然而目前对楸树的研究主要集中在种质资源的调查、收集,速生丰产栽培等技术方面,而关于温度对楸树生长影响的报道较少[8-9]。本研究以自然条件下生长的4种楸树为试材,研究冷驯化期间叶片叶绿素荧光参数的变化规律,旨在为楸树抗寒性理论研究及园林应用提供依据。
综上,这一时期环境保护工作的进展与经济增长基本持平,在某种程度上,各类环境管理制度体系的建设甚至超越了相应的经济发展阶段。这一阶段中国的经济发展仍有较强的中央计划属性,对在环境保护上重走西方国家“先污染、后治理”的道路始终保持着警惕,虽然“摸着石头过河”,但对于先保护、后发展有着强烈的愿望。[注] 李志青:《从经济发展的视角看环境保护40年》,《检察风云》,2018年第13期,第34-35页。20世纪80年代,经济增长和工业化尤其是一些地区乡镇工业的兴起开始给生态环境造成一定的破坏,环境污染问题日益严重,中央政府对环境问题开始高度警觉。但这一时期的环境污染问题多以局部和点状污染为主。
1 材料与方法
1.1 试验材料
选取南北不同纬度的4种1 a生嫁接楸树苗,遗传水平包括:春季发芽早、秋季生长停止晚(滇楸Catalpafargesiif.duclorxii);春季发芽晚、秋季生长停止早(灰楸CatalpafargesiiBur.);春季发芽早、秋季生长停止早(梓楸CatalpaovataG. Don);春季发芽晚、秋季生长停止晚(金丝CatalpabungeiC. A. Mey)。将4种楸树种植于河北农业大学标本园(河北省保定市,北纬38°50′,东经115°26′),试验采用完全随机区组设计,3次重复。
在4种楸树进入冷驯化期间于9月20日开始采样,选取中部位置发育良好且厚度均匀的叶片,每隔15 d采样1次,直至落叶。
1.2 测定指标
抗寒性测定采用电解质渗透法(EL)[10],并利用Logistic方程计算半致死温度。
本文从业务逻辑的角度出发,初步建立指标体系。然而在实际应用中,这些指标是否对个人逾期(在数据集中该指标符号为y)有显著的预测作用,或者说,这些指标与因变量y是否相互独立,则需进一步验证;若独立,则意味着这些指标无法起到评价个人信用状况的作用。此外,从另一个角度理解筛选变量的优点,是可以有效地降低数据集的维度,提高鲁棒性,更多的变量在很大程度上意味着收集变量数据所需成本的增加、模型运行速度的下降、模型训练成本的增加。
正确、合理的思维教学策略与方法是培养学生思维能力的保障.一般地,常用的思维教学的策略可分为以讲课为基础的讲授策略,以事实为基础的问答策略和以思维为基础的问答策略3种.2种典型的思维教学方法分别为:一是以直接讲授为主,以典型问题为例讲解思维过程、方法和规则;二是以思维技能训练为主,在具体学科的教学中教某种思维技能,以综合性项目的实现更好地培养学生的创新精神、创新品质和创新思维[1,9-10].
冷驯化期间楸树叶片ΦPSⅡ变化见图3。
1.3 统计分析
植物通过光合作用利用光能转化合成有机质,而植物的光合器官对低温较为敏感,低温会导致植物光合系统相关酶活性的降低、光合电子传递速率下降,引发光抑制现象,消弱植物利用光能的能力,但植物可以在一定范围内通过热耗散等反应保护自身光合机构[12]。
2 结果与分析
2.1 冷驯化期间楸树叶片抗寒性变化
由图2可知,冷驯化期间,4种楸树叶片的PSⅡ最大光化学效率Fv/Fm值整体均表现为先下降、后上升趋势。滇楸叶片Fv/Fm值在10月5日显著下降后(P<0.05),维持在较低水平;梓楸和灰楸叶片Fv/Fm值在4种楸树中一直处于较高水平,2种楸树一直较为接近,只是在9月20日梓楸显著低于灰楸(P<0.05),其他3个时期差异不显著;金丝楸叶片Fv/Fm值在10月5日显著下降了23.03%(P<0.05),并显著低于其他3种楸树(P<0.05),后期虽有回升,但整体处于较低水平。
由图1可知,9月20日4种楸树叶片的抗寒性较为接近,抗寒性最强与最弱之间仅相差1.16 ℃,但9月20日至10月5日期间灰楸、梓楸和金丝楸叶片抗寒性迅速增强,其中金丝楸抗寒性最强,达到-6.36 ℃,而灰楸叶片抗寒性变化不大,11月1日灰楸叶片抗寒性大幅增强,且远强于其他3种楸树。
2.2 冷驯化期间楸树叶片Fv/Fm变化
冷驯化期间楸树叶片Fv/Fm变化见图2。

图2 冷驯化期间楸树叶片Fv/Fm变化Figure 2 The variation of Fv/Fm in leaves of Catalpa spp. during cold hardening
冷驯化期间楸树叶片抗寒性变化见图1。
2.3 冷驯化期间楸树叶片ΦPSⅡ变化
叶绿素荧光参数测定采用贺立红等的方法[11],测定PS II最大光化学效率或原初光能转换效率(Fv/Fm)、非光化学猝灭(NPQ)、PS II非环式电子传递量子效率(ΦPSⅡ)以及PS II的非循环光合电子传递速率(ETR)。
冷驯化期间楸树叶片NPQ变化见图4。
2.4 冷驯化期间楸树叶片NPQ变化
由图3可知,冷驯化期间,金丝楸叶片光系统Ⅱ光合电子传递量子效率ΦPSⅡ值为下降趋势,而其他3种楸树为先上升、后下降趋势。金丝楸叶片ΦPSⅡ值除9月20日显著高于其他3种楸树外(P<0.05),其他3个时期均低于其他3种楸树;滇楸叶片ΦPSⅡ值最高值出现在10月5日,灰楸和梓楸叶片ΦPSⅡ值最高值出现在10月20日,11月1日3种楸树叶片ΦPSⅡ值均显著低于初始值(P<0.05)。
冷驯化期间楸树叶片ETR变化见图5。
2.5 冷驯化期间楸树叶片ETR变化
由图4可知,冷驯化期间,滇楸叶片的非光化学猝灭NPQ值均在9月20日最高,10月5日显著下降后(P<0.05),维持在较低水平;灰楸、梓楸和金丝楸叶片NPQ值均在10月5日显著下降后(P<0.05),又出现上升趋势,其中金丝楸11月1日达到最高值,不仅显著高于其他3种楸树(P<0.05),而且显著高于自身其他3个时期(P<0.05)。
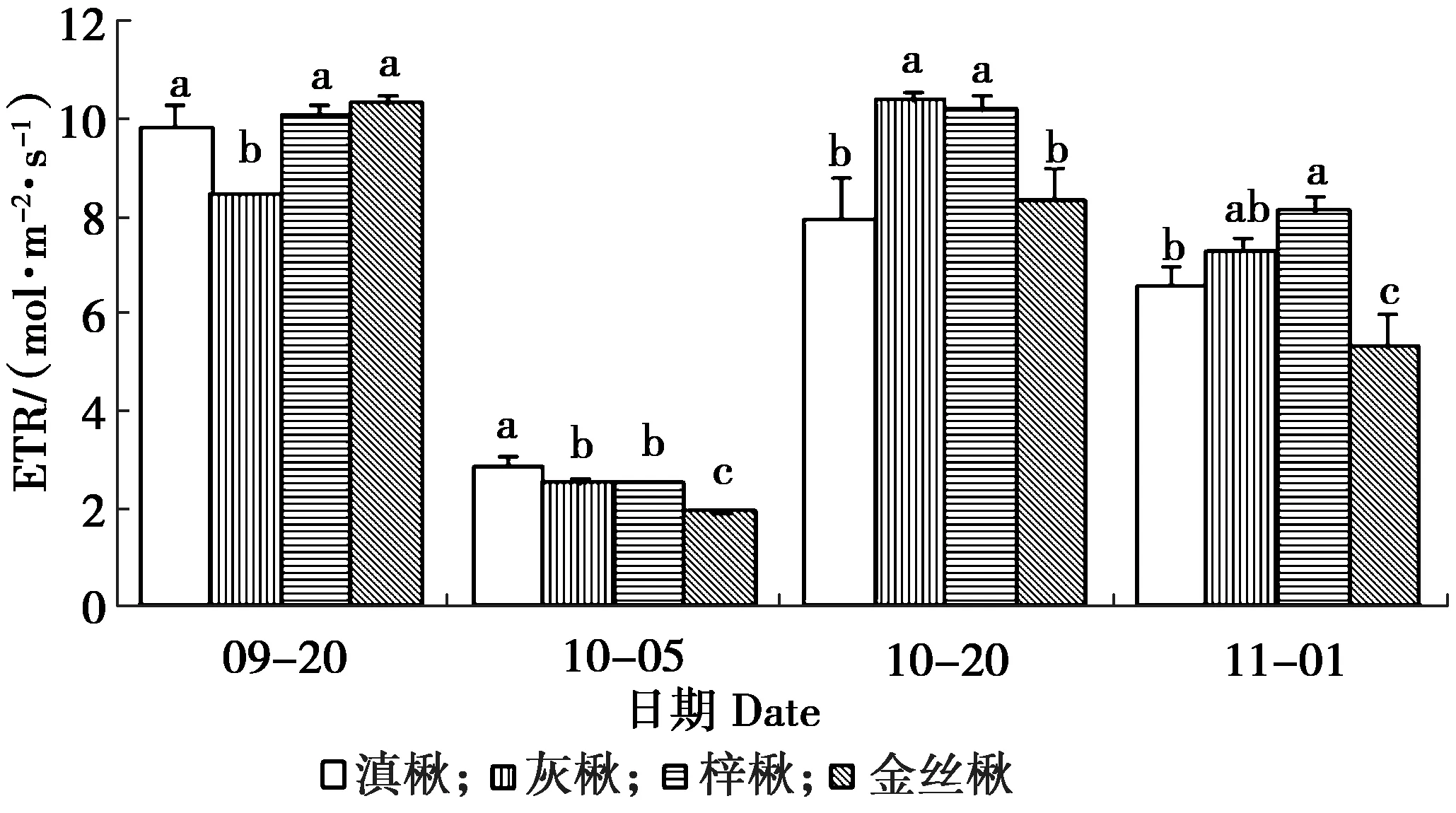
图5 冷驯化期间楸树叶片ETR变化Figure 5 The variation of ETR in leaves of Catalpa spp.during cold hardening
由图5可知,冷驯化期间,4种楸树叶的表观光合电子传递速率ETR值均为先下降、后上升趋势。4种楸树叶片ETR值均在10月5日显著下降到最低值(P<0.05),10月20日又显著上升(P<0.05),其中滇楸和金丝楸叶片ETR最高值出现在9月20日,而灰楸和梓楸最高值出现在10月20日。
3 结论与讨论
利用excel 2010软件进行数据分析整理,SPSS 13.0统计软件进行相对电导法的半致死温度计算以及显著性分析。
采用SPSS19.0统计软件统计分析,计量资料用t检验作组间比较,计数资料组间比较用x2检验,p<0.05表示组间差异存在统计学意义。
众多研究发现,在非胁迫条件下,不同种类和生态型的植物,Fv/Fm稳定在0.832±0.004左右,但低温胁迫会导致植物Fv/Fm的显著下降,而且抗寒性弱的种类比抗寒性强的种类下降更明显[13]。PSⅡ是植物对环境胁迫的最敏感的指示器,Fv/Fm反映PSⅡ反应中心最大光能转换效率,Fv/Fm的下降可以作为反映PSⅡ活性的可靠指标,其变化程度可用来鉴别植物抵抗逆境胁迫的能力[14-17]。王运涛等[18]在苜蓿中研究表明Fv/Fm的减低幅度与品种对低温的敏感性是一致的,Fv/Fm的下降说明低温已经对PSⅡ反应中心造成光化学伤害,Fv/Fm下降的越多说明受伤害程度越大。本研究与前人研究表现一致[19-20],随着冷驯化期间自然温度的降低,4种楸树Fv/Fm整体表现为下降趋势,但灰楸、梓楸和金丝楸10月20日Fv/Fm值有所回升,而滇楸Fv/Fm一直为下降,其中灰楸和梓楸一直具有较高值,尤其在10月5日后,均高于滇楸和金丝楸。同时Fv/Fm的表现趋势与ETR的表现趋势完全一致,表明随着冷驯化的深入,4种楸树均受到了低温的影响发生了光抑制,但4种楸树Fv/Fm降低幅度的差别表明4种楸树对冷驯化的敏感性不同。4种楸树的Fv/Fm与抗寒性存在较高的一致性,可作为楸树抗寒性强弱的鉴定指标。
“每个医生都拥有自己的数字证书和签章,可开具电子申请单和书写电子病历,各种信息都通过信息系统进行传递,并通过数字化病案管理系统实现了病案的电子化归档及管理,节省了工作时间,提高了工作效率。”
许大全[21]研究表明,低温会导致光合电子传递链PSⅡ的最终电子受体减少,会引起PSⅡ功能下降,表现为ETR的降低。王运涛等[18]发现低温胁迫后抗寒性强的苜蓿有效电子传递和光化学效率高,光抑制程度低。本研究结果与王运涛等人研究结果一致,在冷驯化期间4种楸树的PSⅡ整体均呈逐渐下降趋势,但滇楸和梓楸前期PSⅡ有所提高,可能是在冷驯化初期,2种楸树经过低温的诱导,抗性有所上升,但随着冷驯化的深入,4种楸树PSⅡ均表现为下降,但灰楸和梓楸在后期PSⅡ较高,此趋势与灰楸和梓楸在后期具有较高的ETR和Fv/Fm较为一致,表明灰楸和梓楸PSⅡ受伤害程度较小,受低温影响小,抗寒性较强。
On the Need for Public Participation through the “Strange Buildings” Incidents________________LI Jiaqi et al 82
植物在低温胁迫情况下,为避免光合系统尤其是PSⅡ受到破坏,会通过光呼吸、荧光发射、叶黄素循环等途径来消耗植物自身不能利用的过剩光能。非光化学淬灭系数 NPQ表示PSⅡ吸收的光能中不能用于光合电子传递而以热的形式耗散的部分,是植物的一种自我保护机制[22]。本研究表明随着冷驯化的深入,4种楸树NPQ整体呈下降趋势,可能由于低温抑制了叶黄素循环的关键酶活性,影响了叶黄素循环,而在冷驯化后期灰楸、梓楸和金丝楸NPQ有所回升,表明这3种楸树PSⅡ反应中心仍具有一定的自我恢复能力,说明关键酶活性受损程度较轻,适应低温胁迫能力较好,滇楸NPQ一直表现为下降,说明其PSⅡ反应中心受伤害程度较高,已经超出其自身修复范围。
本研究中,4种楸树在自然低温胁迫下,PSⅡ最大光化学效率降低,PSⅡ潜在活性降低,非光化学淬灭降低,电子传递速率减慢,这些变化均表明楸树PSⅡ反应中心受到低温的影响,导致光合能力减弱,造成了光抑制。
