猪ALAS1基因的多态性与产仔数性状的关联分析
2021-01-27刘林清李庆岗苏世广王重龙
刘林清,李庆岗,苏世广,周 梅,张 威,王重龙
(安徽省农业科学院畜牧兽医研究所,猪分子数量遗传学安徽省农业科学院重点实验室,畜禽产品安全工程安徽省重点实验室,安徽 合肥 230031)
ALAS1基因,一个吡哆醛5'磷酸(PLP)限速酶,它属于α-氧基胺合成酶家族。ALAS1基因参与血红素生物合成的第一步反应,并且对血红素的合成起到限速酶的作用,以维持细胞内血红素的水平[1]。有研究发现,在ALAS1基因的5'侧翼序列存在两个cAMP应答元件(CRE)识别序列,cAMP调节信号转导通路在卵巢卵泡发育和黄体化过程中起到主要作用,并可诱导ALAS1基因的表达[2]。还有研究者们发现,ALAS1基因还在卵巢中参与调节与排卵过程有关的线粒体细胞色素P450和类固醇代谢及类固醇激素的产生[3-6]。此外,Lawrence等[7]在小鼠卵巢中注射HCG(人绒毛膜促性腺激素)0~39 h后发现,ALAS1基因在早期排卵过程中在卵泡组织中上调表达,并且可能参与卵泡黄体化过程。这些研究结果进一步暗示了ALAS1基因在卵巢卵泡发育及排卵过程后都发挥着作用。
为此,本研究建立了ALAS1基因的PCR-MspIRFLP分型技术,并在3个中外猪种中进行多态性分型,还在两个猪群中检测该位点的多态性并与产仔数性状进行关联分析,以期为提高猪产仔数提供一个有用的分子标记。
1 材料与方法
1.1 材料
1.1.1 试验动物及组织采集 试验地点在安徽省科鑫养猪育种有限公司;试验动物:于2014年7月采集58头淮猪、75头长白猪及85头淮猪新品系Ⅱ系猪的耳组织放置于75%酒精的离心管中,-20 ℃保存备用;饲养管理条件和饲粮营养水平一致。
1.2 方法
1.2.1 DNA提取 采用苯酚-氯仿抽提法,TE溶解,-20 ℃冷冻保存。
1.2.2 引物的设计与合成 根据ALAS1基因DNA序列,应用Premier 5.0软件在多态位点(第9内含子)两侧,设计1对引物,突变位点可用限制性内切酶MspI识别,引物由上海生工生物公司合成。引物信息如下:ALAS1基因的上游引物:5'-CACACCCCGCAGATGATGAC-3',下游引物:5'-AAATAGAAGTGCAGAGCCCAGC-3',片段长度为359 bp,退火温度为62.5 ℃。
1.2.3 PCR扩增 PCR反应体系为25 μL,其中PCR×2Taqmix 12.5 μL,10 mmol/μL的引物各0.5 μL,DNA模板1 μL(DNA约100 ng),加ddH2O至25 μL。PCR反应条件为第1步95 ℃变性5 min;第2步95 ℃变性45 s;第3步62.5 ℃复性40 s;第4步72 ℃延伸45 s;重复第2至第4步35个循环,之后再72 ℃延伸10 min,最后降温至4 ℃保存。PCR扩增产物用浓度为1.5%的琼脂糖凝胶电泳,核酸染料染色,凝胶成像系统中观察,可见到片段大小为359 bp的清晰条带。
1.2.4 限制性内切酶酶切(RFLP)ALAS1基因的PCR产物用MspI内切酶酶切,反应体系为:10×Buffer 1 μL,MspI酶0.20 μL(20 U/μL),加PCR产物至10 μL;37 ℃反应8~12 h;然后,酶切产物在2%琼脂糖凝胶中电泳,核酸染料染色,凝胶成像系统中观察。
1.3 数据处理分析
1.3.1 统计分析的软件 采用SAS 8.0统计软件(SAS Institute Inc, Version 8 Edition)的GLM程序进行标记方差分析,并进行显著性检验。
1.3.2 基因效应统计分析模型 依据Liu等[8]所述方法建立的单标记回归统计模型,除了猪个体为随机效应以外,其他因素均为固定效应。所采用模型如下:Y=平均值+基因型+胎次效应+残差,其中Y为性状表型值。
2 结果与分析
2.1 猪ALAS1基因的多态位点检测
PCR扩增片段长度为359 bp,经检测与预期结果一致(图1),片段本身就存在一个MspI酶切位点,经MspI酶切后产生3种基因型(图2)。当多态位点为CC基因型时,则造成了2个MspI酶切位点,PCR产物经MspI消化后产生3个片段(277 bp+35 bp+47 bp),记为等位基因C;当多态位点为TT基因型时,则一个酶切位点消失,PCR产物经MspI消化后产生1个片段(324 bp+35 bp),记作等位基因T。
2.2 猪ALAS1基因在不同猪群体中的基因型和等位基因的频率分布
由表1可见,ALAS1基因MspI酶切位点在所检测的3个猪群中,国内地方品种淮猪猪群中C等位基因占优势,而从国外引进的长白猪和自主培育的淮猪新品系Ⅱ猪群中T等位基因频率较高。
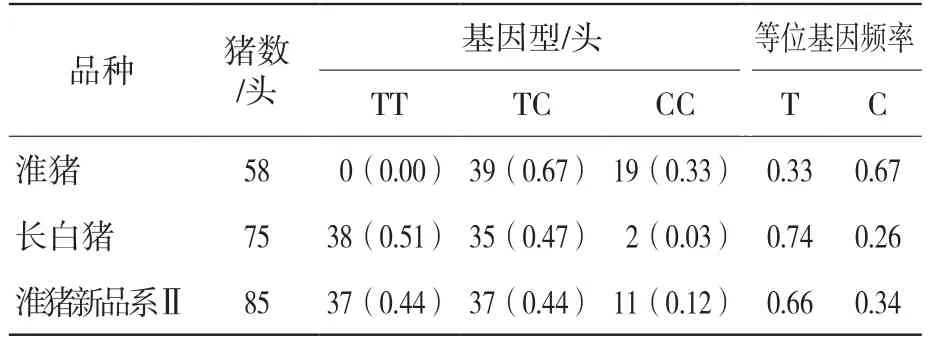
表1 ALAS1基因PCR-MspI-RFLP基因型频率和等位基因频率分布
2.3 猪ALAS1基因MspI多态位点基因型与猪产仔数性状的关联分析
应用基因效应统计分析模型,采用SAS统计软件的GLM程序分别在长白猪和淮猪新品系Ⅱ猪群体中进行猪ALAS1基因PCR-MspI-RFLP多态位点与产仔数性状的关联分析。
在长白猪和淮猪新品系Ⅱ猪群体中,所有胎次产仔数均呈现CC>TC>TT的趋势,其中CC型个体和TC型个体的所有胎次产仔数分别显著高于TT型个体(P<0.05),详见表2。统计结果显示,CC型母猪具有较高的产仔数,C等位基因为优势等位基因。
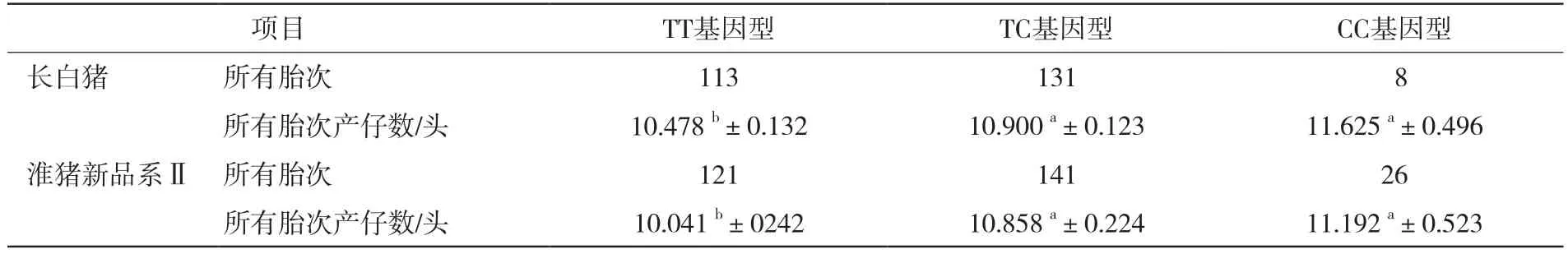
表2 ALAS1基因PCR-MspI-RFLP基因型与长白猪、淮猪新品系Ⅱ产仔数性状的统计分析
3 讨论
淮猪是我国地方猪种,具有生长速度慢、繁殖力高等特性。长白猪是引进的外来猪种,其具有生长速度快、繁殖力不高等特性。而淮猪新品系Ⅱ是以安徽省优良地方品种淮猪为母本,以国外优质瘦肉型品种长白猪、大白猪为父本,经过杂交育种组建基础群,然后运用标记辅助选择BLUP估计育种值进行群体继代选育的。ALAS1基因,一个吡哆醛5'磷酸(PLP)限速酶,它属于α-氧基胺合成酶家族。笔者前期采用RTPCR方法的分析猪ALAS1基因在用PMSG(孕马血清促性腺激素)和HCG(人绒毛膜促性腺激素)处理卵巢颗粒细胞不同时间后的表达情况,结果表明,ALAS1基因在卵泡颗粒细胞生长、发育等过程中瞬时表达[9],这与其他研究者的结果是一致的[7]。这些研究结果进一步显示了ALAS1基因在卵巢卵泡发育及排卵过程后都发挥着作用。所以,我们选择ALAS1基因作为研究猪产仔数性状的候选基因。
本研究发现,ALAS1基因MspI酶切位点在所检测的3个猪群中,国内地方品种淮猪猪群中C等位基因占优势,而从国外引进的长白猪和自主培育的淮猪新品系Ⅱ猪群中T等位基因频率较高。并且在长白猪和淮猪新品系Ⅱ猪群体中,所有胎次产仔数均呈现CC>TC>TT的趋势。其中,CC型个体和TC型个体的所有胎次产仔数分别显著高于TT型个体(P<0.05),从以上结果表明,CC型母猪具有较高的产仔数,C等位基因为优势等位基因。因此选择CC型个体留种可提高群体的产仔数,在今后选育中可适当提高等位基因C的频率。
