激素性股骨头坏死mRNA与非编码RNA差异表达谱及竞争性内源RNA调控网络分析
2021-01-22李嘉程杨曦梁学振阎博昭许波李刚
李嘉程,杨曦,梁学振,阎博昭,许波,李刚
1山东中医药大学第一临床医学院,济南 250014;2新疆军区总医院检验科,乌鲁木齐 830000;3山东中医药大学附属医院骨科,济南 250014
激素性股骨头坏死(steroid-induced osteonecrosis of the femoral head,SONFH)是骨科常见的难治性疾病之一。长期服用激素会对股骨头产生一定的影响,最终加重股骨头的缺血,引起成骨-破骨代谢异常,进而导致塌陷。塌陷是股骨头软骨下骨力学属性失败的重要特征,是决定SONFH治疗方案最重要的因素[1],目前SONFH发病趋于年轻化且治疗困难,已成为导致青壮年髋关节残疾的最常见原因之一[2]。因此,深入了解SONFH发生发展机制,寻找新的分子标志物以提高其早期诊断率、深入探究其发病的分子机制以提高药物的治疗效果,对于改善SONFH患者的预后具有重要意义。迄今为止,大量非编码RNA(non-coding RNAs,ncRNAs)已被发现参与控制基因表达和激活途径,从而影响SONFH的进展。鉴于长链非编码RNA(long noncoding,lncRNAs)自身表达的丰富性和组织表达的特异性,组织中表达失调的lncRNAs可作为SONFH早期诊断和预后判断的分子标志物。此外,研究表明,作为miRNAs的竞争性内源RNA(competitive endogenous RNAs,ceRNA),lncRNAs在疾病的发生和发展中发挥关键作用,是人类多种疾病的重要调节因子。然而,大多数lncRNAs在SONFH发生中的生物学作用和临床意义尚不完全清楚。为此,本研究通过生物信息学方法分析SONFH的相关差异表达基因,构建lncRNA、miRNA及mRNA之间的ceRNA调控网络,探讨差异表达基因的生物学功能及潜在机制。
1 材料与方法
1.1 数据来源 在高通量测序SRA数据库(https://www.ncbi.nlm.nih.gov/sra)检索SONFH相关去核糖体的转录组测序数据集SRP091890,由Huashan Hospital,Fudan University上传(https://www.ncbi.nlm.nih.gov/sra/SRX2255530),分析数据可得到lncRNA和mRNA的表达谱。在GEO数据库(https://www.ncbi.nlm.nih.gov/geo/)检索SONFH相关的miRNA芯片,获得编号为GSE89587的芯片原始数据和编号为GPL21439的芯片基因注释文件,该芯片数据的原始文件包含20个血清样本,由Zhang等[3]上传,分析数据可得到差异表达的miRNAs数据。
1.2 差异表达基因筛选 应用R 语言程序包(http://bioconductor.org/packages/release/bioc/html/edgeR.html)对上述数据进行背景校正和矩阵数据归一化处理,利用Limma包分析不同芯片原始数据的差异基因,数据结果运用log2进行转化,显著差异基因的筛选条件为P<0.05且差异倍数(fold change,FC)≥1.5;运用plot包和heatmap2包分别绘制芯片原始的火山图和聚类图。
1.3 SONFH相关miRNA-DEG调控网络的构建 使用miRDB(http://mirdb.org/)[4]、miRTarbase(http://mirtarbase.mbc.nctu.edu.tw/php/index.php)[5]和TargetScan(http://www.targetscan.org/vert_72/)[6]等数据库检索miRNA靶向的mRNAs。为了提高搜索结果的有效性,纳入两个及以上数据库中包含miRNA靶向的mRNAs,以构建miRNA-mRNA调控网络,并利用Cytoscape 3.6.1软件对上述结果进行可视化映射。
1.4 SONFH相关lncRNAs与lncRNA-miRNA-mRNA调控关系的预测 利用starBase(http://starbase.sysu.edu.cn /index.php)[7]数据库预测SONFH相关miRNA调控的lncR NA s,与通过基因芯片分析得出的lncRNA-DEG取交集,以构架SONFH可能的lncRNA相关ceRNA网络(图1),利用clusterProfiler包[8]对上述差异mRNAs进行功能富集分析。
2 结 果
2.1 SONFH相关的差异表达mRNAs、miRNAs及lncRNAs 利用R语言程序包中的limma包分析差异表达的mRNAs、miRNAs及lncRNAs,共筛选得到422个显著差异mRNAs,其中192个表达上调、230个表达下调;22个显著差异miRNAs,其中10个表达上调、12个表达下调;416个显著差异lncRNAs,其中148个表达上调、268个表达下调。所有差异表达mRNAs、miRNAs及lncRNAs的火山图和聚类图如图2-4所示。
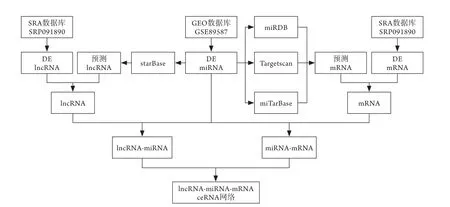
图1 SONFH相关的lncRNA-miRNA-mRNA ceRNA调控网络流程Fig.1 Flow chart of lncRNA-miRNA-mRNA ceRNA regulation network related to SONFH SONFH. 激素性股骨头坏死
2.2 预测与差异表达miRNAs相关联的mRNAs 利用TargetScan、miRTarBase、miRDB数据库预测与上述差异表达miRNAs相关联的mRNAs,至少2个数据库预测得到的mRNAs纳入研究,最后得到8324个预测miRNA-mRNA(图5),去重后得到1993个mRNAs,与通过基因芯片分析得出的差异mRNAs取交集得到46个mRNAs。映射差异表达的mRNAs和miRNAs预测的mRNAs,构建mRNA-miRNA 网络。
2.3 预测与差异表达miRNAs关联的lncRNAs及lncRNA-miRNA-mRNA调控关系的构建 利用starBase数据库预测与SONFH相关miRNAs调控的lncRNAs 196个,与通过基因芯片分析得出的差异lncRNAs取交集得到3个lncRNAs。映射差异表达的lncRNAs和miRNAs预测的lncRNAs,根据lncRNAmiRNA上下调关系构建网络,仅VASH1-AS1符合条件,将其与mRNA-miRNA网络构建lncRNA-miRNAmRNA调控网络,如图6所示。
2.4 差异表达基因的GO富集分析和KEGG通路分析 利用clusterProfiler包对上述差异mRNAs进行GO富集分析,结果显示,6条生物学进程被显著富集,包括氮化合物代谢过程、分解代谢过程、生物合成过程及代谢过程的正负调控等(图7)。但KEGG信号通路未被显著富集。
3 讨 论
股骨头坏死(osteonecrosis of femoral head,ONFH)是骨科常见的难治性疾病之一。流行病学调查显示,日本每年有2500~3000例新发病例,美国为1.5万~2万,韩国为1万~1.5万,我国为10万~ 20万。我国2012年一项多中心调查研究显示,约26.35%的男性和55.75%的女性非创伤性ONFH患者是由激素引起的。长期服用激素会对股骨头产生一定的影响[9]:①促进血小板生成,增高血液凝固性和黏滞度,在末梢小动脉炎的基础上发生动脉栓塞;②加重髓内静脉淤滞,增高骨内压,引起骨内循环障碍;③降低病变股骨头软骨下骨微血管密度,导致血管通透性异常和血管新生能力变差等。上述病理变化最终加重股骨头缺血[10],引起骨形成动态平衡体系的破坏,进而导致塌陷,而塌陷是股骨头软骨下骨力学属性失败的重要特征,最终导致髋关节残疾。但SONFH发生的具体机制目前尚不清楚,阐明SONFH相关基因及其参与ONFH进展的分子机制至关重要。
近年来,lncRNAs的相关研究引起了各个领域的关注。大量证据表明,lncRNAs在疾病发生和发展中起着非常重要的调控作用,但lncRNAs与SONFH关系的研究较少。本研究在SONFH患者和正常对照之间共筛选出422个显著差异mRNAs、 22个显著差异miRNAs和416个显著差异lncRNAs,经过与预测的mRNAs和lncRNAs去交集后,构建了一个VASH1-AS1/miRNA/mRNA相关的ceRNA调控网络。
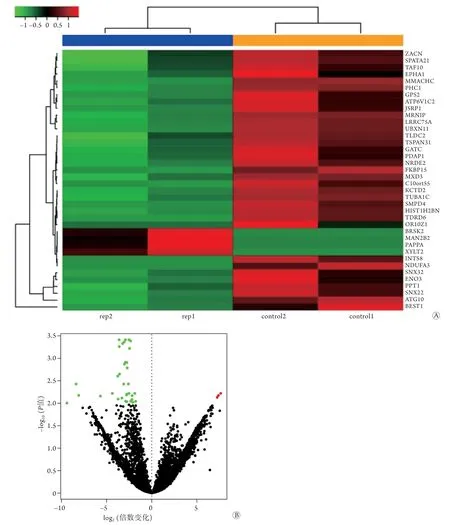
图2 SONFH相关的差异表达mRNAs的聚类图(A)和火山图(B)Fig.2 Volcano diagram and clustering diagram of mRNAs differently expressed in SONFH SONFH. 激素性股骨头坏死;蓝色示SONFH;橙色示对照组;红色示上调基因;绿色示下调基因
VASH1-AS1为VASH-1(vasohibin-1)的反义RNA,是最近发现的能够通过负反馈调节血管生成的调控因子[11-12],在血管内皮细胞特异性表达,且其表达随血管生成刺激因子如血管内皮生长因子(VEGF)和碱性成纤维细胞生长因子(bFGF)的表达而增加[13]。VASH-1的特殊性在于其以负反馈调节的方式抑制血管生成,特异性作用于血管内皮细胞,是目前唯一有科学研究依据的、专一抑制血管新生的抑制因子[14]。而VASH1-AS1是VASH-1的反义RNA,与VASH-1特异性互补结合,抑制其翻译、表达,阻断该基因的功能。近年来研究发现,血管新生在哺乳动物器官发育、组织修复和再生等过程中发挥着关键的作用,而且与多种高发疾病如心肌梗死、骨坏死、肿瘤及视网膜病变等的发生和发展密切相关[15]。VASH1-AS1可通过抑制VASH-1而发挥促进血管生成的作用。
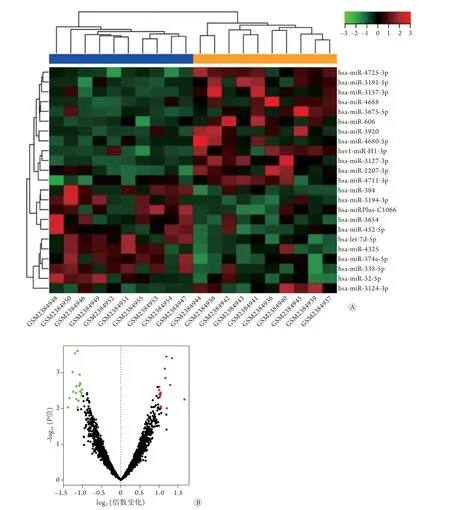
图3 SONFH相关的差异表达miRNAs的聚类图(A)和火山图(B)Fig.3 Volcano diagram and clustering diagram of miRNAs differently expressed in SONFH SONFH. 激素性股骨头坏死;蓝色示SONFH;橙色示对照组;红色示上调基因;绿色示下调基因
let-7最早在线虫中发现,负责调节细胞的增殖和分化,随后有研究表明,let-7家族miRNAs在哺乳动物中的序列和功能都高度保守。let-7家族miRNAs作用广泛,能够参与调控干细胞发育、凋亡、分化、免疫应答、肿瘤发生及转移等多种生理病理过程[16]。徐小隔等[17]发现,let-7d在骨髓间充质干细胞向神经细胞的分化中起着重要作用。上调SHR/WKY-VSMCs、HCASMCs等血管平滑肌细胞内let-7d的表达可有效抑制KRAS蛋白的表达,并能抑制血管平滑肌细胞的增殖[18]。
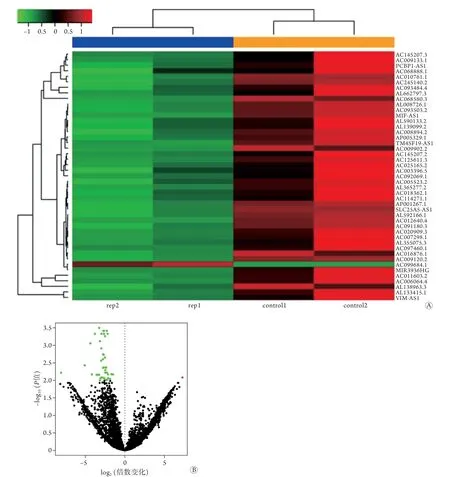
图4 SONFH相关的差异表达lncRNAs的聚类图(A)和火山图(B)Fig.4 Volcano diagram and clustering diagram of lncRNAs differently expressed in SONFH SONFH. 激素性股骨头坏死;蓝色示SONFH;橙色示对照组;红色示上调基因;绿色示下调基因
SLC5A6是一种钠依赖的多种维生素转运蛋白(SMVT),介导细胞对生物素、泛酸和硫辛酸的摄取[19]。Uchida等[20]证实,人脑微血管中SLC5A6 mRNA和蛋白均呈高表达,且SLC5A6对人脑毛细血管内皮细胞摄取生物素和泛酸起主要作用,提示SLC5A6是脑毛细血管内皮细胞摄取生物素和泛酸转运的主要因子。
NQO2基因位于6号染色体的6pte-q12片段,可降低细胞内氧自由基(reactive oxygen species,ROS)的水平,抑制细胞因子ERK1/2的磷酸化,从而抑制细胞增殖。部分研究表明,NQO2高表达可能会激活大脑中的某些化学物质,导致氧化应激,引起神经元损伤[21]。也有研究表明,NQO2可防止醌诱导的皮肤癌变[22]。Yu等[23]发现,NQO2表达增加可降低乳腺癌发生的可能性,尤其是具有野生型p53的乳腺组织。目前,通过检索文献尚未发现hsamir-3194-3p和THAP6的单独研究。
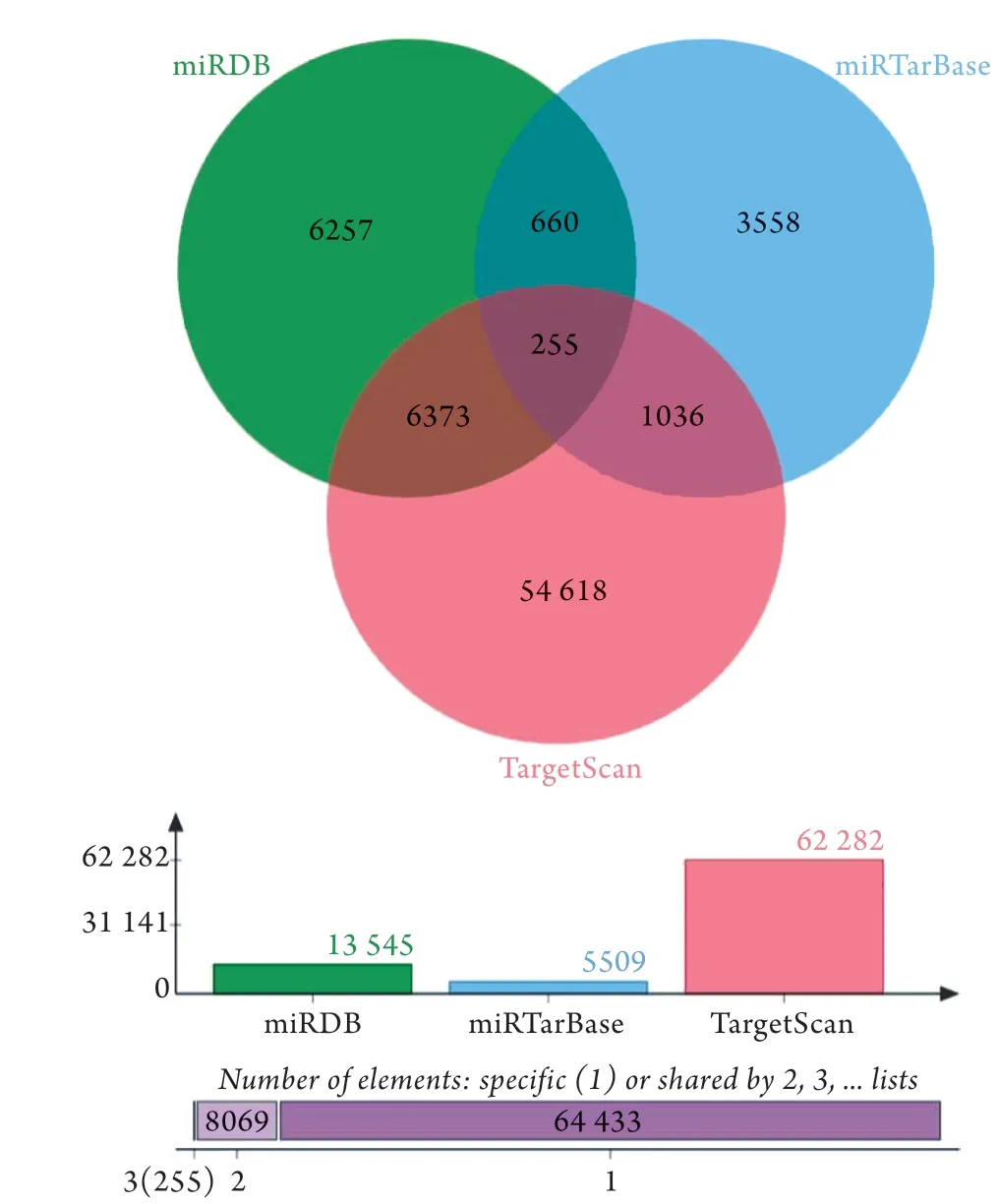
图5 差异表达miRNAs预测的mRNAs Veen图 Fig.5 Veen plot of mRNAs predicted by differentially expressed miRNAs
骨坏死的发生发展是多因素、多环节、多阶段、多基因参与的动态过程[24]。既往研究一般聚集于骨细胞,从基因、分子、表观表型等方面阐述骨坏死的发生发展过程,而忽略了骨骼血管微环境的影响。骨骼血管微环境是由调节骨生长、骨骼动态平衡的多种基质细胞及其所分泌的血管生成因子构成的。骨修复过程中的血管化是骨再生的重要调控因素,“血管新生-骨形成耦联”机制是调控骨重建修复的关键机制[25],而VASH1-AS1、has-let-7d-5p和NQO2可能作为调控因子,通过调控网络参与调节“血管新生-骨形成耦联”机制,在SONFH发病过程中发挥了关键作用。
本研究存在一定的局限性:①纳入样本量有限,可能导致筛选的lncRNAs、miRNAs和mRNAs的数量有所偏差,未来需扩大研究样本量进行证实;②本研究结果仅涉及SONFH血液标本,需要进一步研究其他类型的ONFH(如创伤性ONFH)和其他来源(如骨组织)的样本以更加准确地阐释ONFH的生理病理过程。
综上所述,本研究通过整合分析SONFH血液中lncRNAs、miRNAs和mRNAs的表达差异,成功构建了lncRNAs相关的ceRNA调控网络,发现VASH1-AS1可能通过has-let-7d-5p或hsa-mir-3194-3p间接调控SLC5A6、NQO2和THAP6,从而参与SONFH的发病过程,其中VASH1-AS1、has-let-7d-5p和NQO2可能作为调控因子,通过调控网络参与调节“血管新生-骨形成耦联”机制,在SONFH发病过程中发挥关键作用。这一发现为SONFH的作用机制研究提供了新思路,为其治疗提供了潜在靶点,但仍需进一步研究验证这一调控效应。
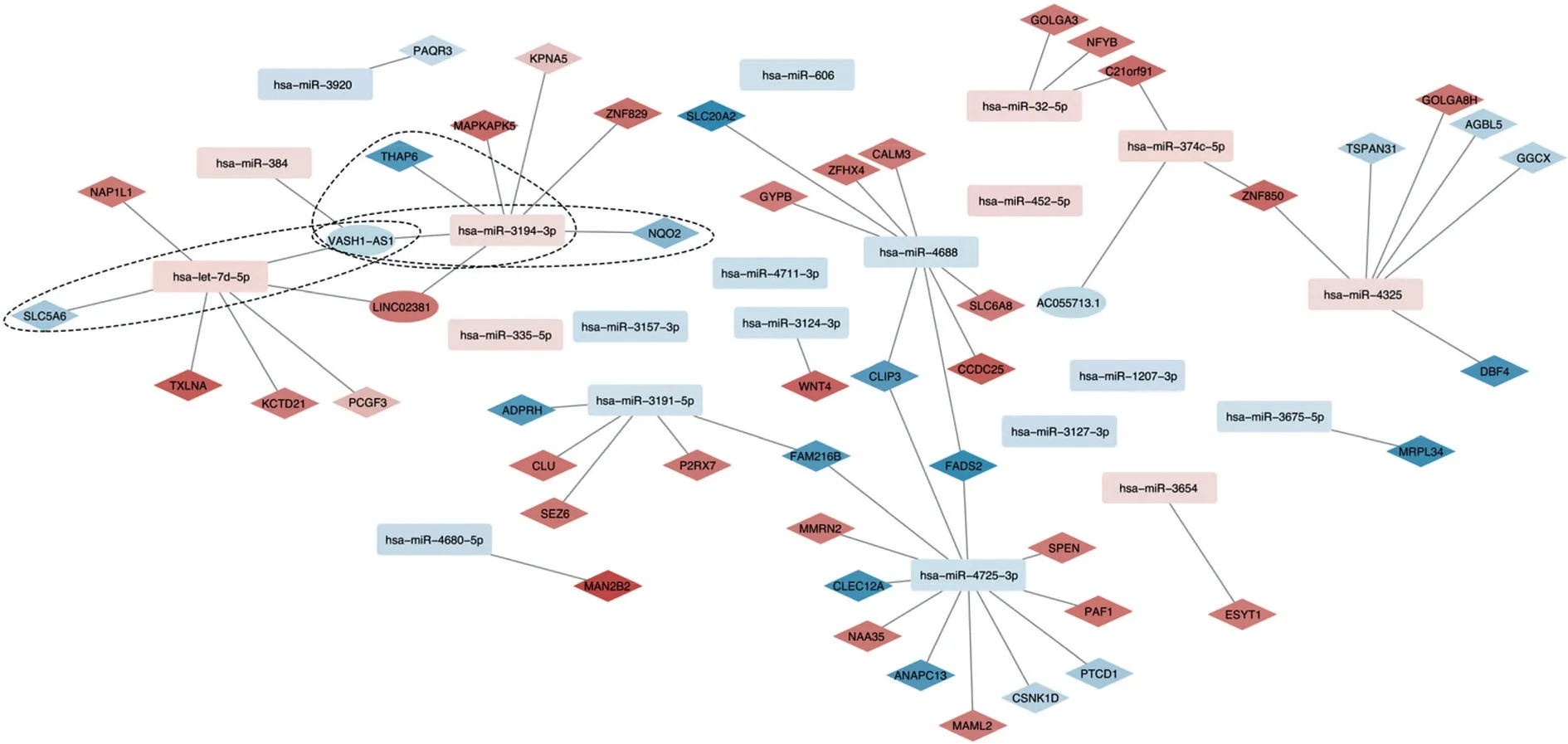
图6 SONFH相关的lncRNA-miRNA-mRNA调控网络Fig.6 SONFH-related lncRNA-miRNA-mRNA regulatory network
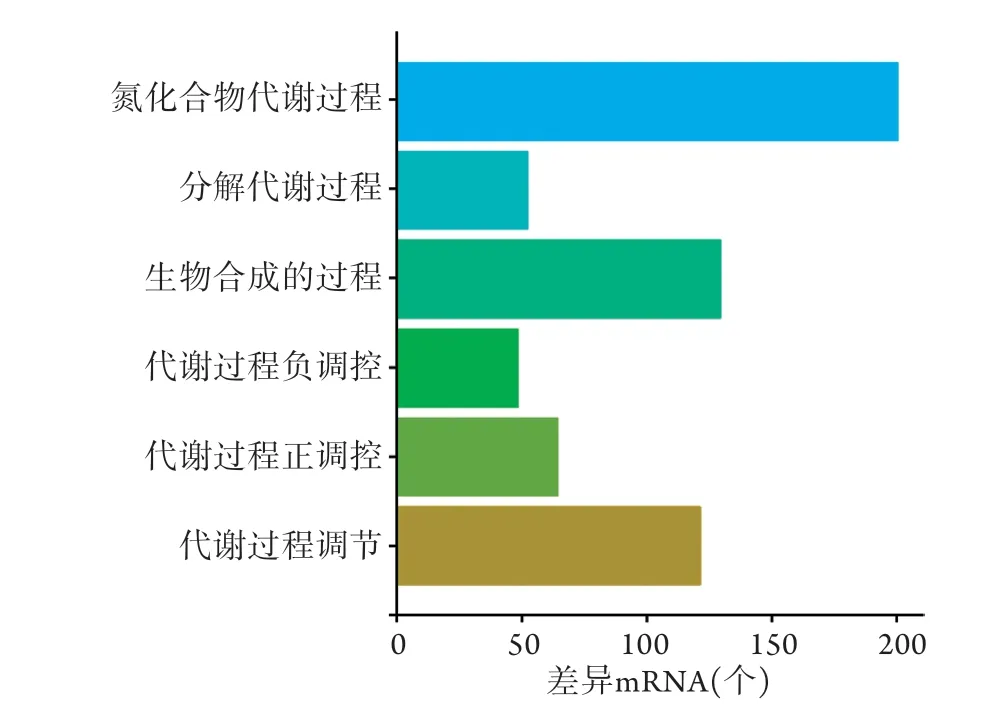
图7 SONFH相关的差异表达mRNAs的GO功能富集分析Fig.7 GO functional enrichment analysis of SONFH-related differentially expressed mRNAs
