大丽轮枝菌淀粉酶VdAmy-1的致病功能*
2021-01-22王莹蕾赵钰莹王小叶刘子翔徐云巧
王莹蕾, 赵钰莹, 刘 旭, 王小叶, 刘子翔 徐云巧, 朱 妍, 庄 靖, 田 李
(曲阜师范大学生命科学学院,273165,山东省曲阜市)
大丽轮枝菌是引发多种农作物(包括棉花、茄子等重要经济作物)产生黄萎病的土传性病原真菌,在世界范围内造成重大经济损失[1,2].长期的致病机理研究发现,大丽轮枝菌胞外分泌蛋白在侵染过程发挥重要作用,是最终引起寄主植株发病和死亡的主要因素[3].胞外蛋白大致分为两类:一是扮演毒力因子或者参与调控寄主免疫反应的效应蛋白.如蛋白谱发现的小分子量富含半胱氨酸类蛋白VdSCP7,可诱导植物的免疫反应[4];比较基因组学发现的几丁质结合模体蛋白LysM抑制几丁质诱导的植物免疫反应[5].另外一类是具有酶活的分泌型蛋白.分泌蛋白中的大量酶类可以降解寄主植物的细胞壁和细胞内物质,这不仅有利于病原真菌的侵入与扩展,也为病原菌的生存提供了必要的碳源.根据酶作用的底物,细胞壁降解酶可分为果胶酶、纤维素酶等种类.目前,研究人员通过基因敲除实验发现果胶酶裂解酶VdPL3.1/3[6],纤维素酶VdEG1/3[7]均参与大丽轮枝菌的致病性.这可能与大丽轮枝菌需要降解寄主富含果胶和纤维素的细胞壁,从而进入维管束的生活史特点密切相关[8].淀粉是植物细胞内的重要碳源,大丽轮枝菌如何利用寄主的淀粉作为其营养物质一直不明确.本研究选取了一个大丽轮枝菌淀粉酶,利用基因敲除技术对其功能进行了初步研究.
1 材料与方法
1.1 大丽轮枝菌Vd991淀粉酶基因VdAmy-1的克隆
根据酵母菌淀粉酶蛋白序列,在NCBI大丽轮枝菌基因组数据库中进行BLASTp搜索.根据数据库中该蛋白的编码基因序列,设计可扩增该基因全长的引物Amy-F/R (见表1).分别以大丽轮枝菌Vd991基因组DNA和cDNA为模板,扩增该基因的全长序列和编码框序列.PCR产物送交上海生工生物工程有限公司测序.
1.2 VdAmy-1信号肽功能验证
PCR扩增编码信号肽的基因序列,将扩增产物连接到EcoRⅠ和XhoⅠ双酶切的pSUC2载体(该载体含有缺失信号肽的蔗糖转化酶基因SUC).利用聚乙二醇/醋酸锂(PEG/LiAc)法将重组载体转入YTK12酵母菌株(该酵母菌株缺失蔗糖转化酶基因SUC,在以棉子糖为唯一碳源的培养基中不能生长).将上述酵母菌培养在以棉子糖为唯一碳源的YPRAA培养基 (1% yeast extract,2% peptone,2% raffinose,2 μg/Μl antimicyn A),通过观察酵母是否生长,判断蔗糖转化酶SUC是否可有效分泌.
1.3 VdAmy-1基因敲除突变体的构建
按照高效大丽轮枝菌基因敲除方法进行[9].简要来说,使用引物K1-K6分别扩增VdAmy-1基因上游、下游和潮霉素抗性基因片段,融合 PCR方法构建基因敲除载体,然后通过农杆菌介导的方法转化大丽轮枝菌.在添加有50 μg/mL头孢霉素、30 μg/mL潮霉素和50 μmol/L F2dU的PDA培养基中筛选基因敲除转化子,并利用VdAmy-1基因内部引物K7-K8对其进行PCR分子验证.将野生型VdAmy-1基因重新导入突变体细胞,得到回补菌株.本文所使用的引物见表1.

表1 本研究中使用的引物
1.4 突变体菌株利用淀粉能力分析
分别取5 μL浓度为1×106个/mL野生型菌株、突变体菌株和功能互补菌株的孢子点接至1%淀粉作为唯一碳源的固体查比克盐(NaNO32 g/L,KCl 0.50 g/L,FeSO40.01 g/L,KH2PO41.0 g/L,MgSO4·7H2O 0.50 g/L) 培养基上(以丰富培养基PDA为对照),25 ℃恒温培养箱中培养9 d并拍照,根据菌落大小定性推测各菌株利用淀粉能力的差异.
1.5 致病力检测
在棉苗子叶期,将棉株连根拔起.将根在无菌水中冲洗,然后置于大丽轮枝菌的孢子悬浮液中5 min.每个菌株接种10株棉苗,共设3个重复.接菌后的第4周拍照并调查病情指数[10].
2 结果与讨论
2.1 大丽轮枝菌VdAmy-1基因的序列分析
根据酵母菌淀粉酶蛋白序列,在大丽轮枝菌 Vd991基因组数据库中进行 BLASTp 搜索,得到大丽轮枝菌Vd991中的同源蛋白,Genbank编号为VEDA_08865,本研究中命名为VdAmy-1.基因测序结果显示,该基因全长1967 bp,含有1个内含子,编码635个氨基酸残基.大丽轮枝菌VdAmy-1蛋白与酵母菌淀粉酶蛋白具有76%的氨基酸序列同源性,其中C端的淀粉酶结构域的同源性高达91%.蛋白质保守功能域预测表明,在该蛋白的 N端有典型的信号肽,C端有1个保守的淀粉酶(carbohydrate-binding module,family 20)结构域,说明VdAmy-1蛋白是一个分泌型淀粉酶.
2.2 VdAmy-1的N端信号肽具有分泌活性
本研究利用酵母信号肽捕获系统对其信号肽活性进行验证[11].YTK12 酵母菌株为蔗糖转化酶SUC缺陷性菌株,不能够在只含有棉籽糖作为唯一碳源的培养基YPRAA中生长.pSUC2质粒中含有缺失信号肽的SUC2编码基因,只有在该基因前插入编码信号肽的核酸序列,并将重组pSUC2质粒转化入YTK12菌株中,这样蔗糖转化酶才可以正常分泌至胞外,从而使YTK12菌株可以利用棉籽糖作为碳源生长.
信号肽预测程序 SignalP 5.0显示VdAmy-1蛋白质N端前21个氨基酸为信号肽序列.本研究将VdAmy-1编码信号肽的核苷酸序列连接到 pSUC2 载体中,转入YTK12酵母细胞中进行信号肽活性验证.结果显示,YTK12菌株和含有pSUC2空载体的菌株不能够在YPRAA培养基中生长,而含有VdAmy-1信号肽的酵母菌株可以在YPRAA培养基中正常生长(图1),说明该蛋白的信号肽具有分泌活性,VdAmy-1为大丽轮枝菌分泌型蛋白.

图1 酵母信号肽捕获系统验证VdAmy-1信号肽活性.已知的信号肽Avr1b作为阳性对照
2.3 获得大丽轮枝菌突变体菌株KO和功能回补菌株EC
通过农杆菌介导的方法,将基因敲除载体导入野生型菌株(Wild Type,WT)进行遗传转化,共得到26个转化子.选取随机挑选其中2个候选基因敲除的转化子,对其进行 PCR验证并命名为KO-1/2.为进一步证明突变体菌株KO-1/2相对于野生型的表型变化是由于VdAmy-1基因缺失所造成的,本研究将VdAmy-1基因重新导入KO-1/2,构建功能回补菌株EC-1/2.PCR验证显示(图2),以野生型菌株基因组作为模板,利用VdAmy-1基因内部引物K7-K8,可成功扩增出0.7 kb的VdAmy-1基因片段.以突变体菌株KO-1/2基因组作为模板,不能扩增出VdAmy-1基因所对应的条带,回补菌株EC-1/2又重新扩增出VdAmy-1基因所对应的条带(图2).上述结果说明已经成功构建了突变体和功能回补菌株.
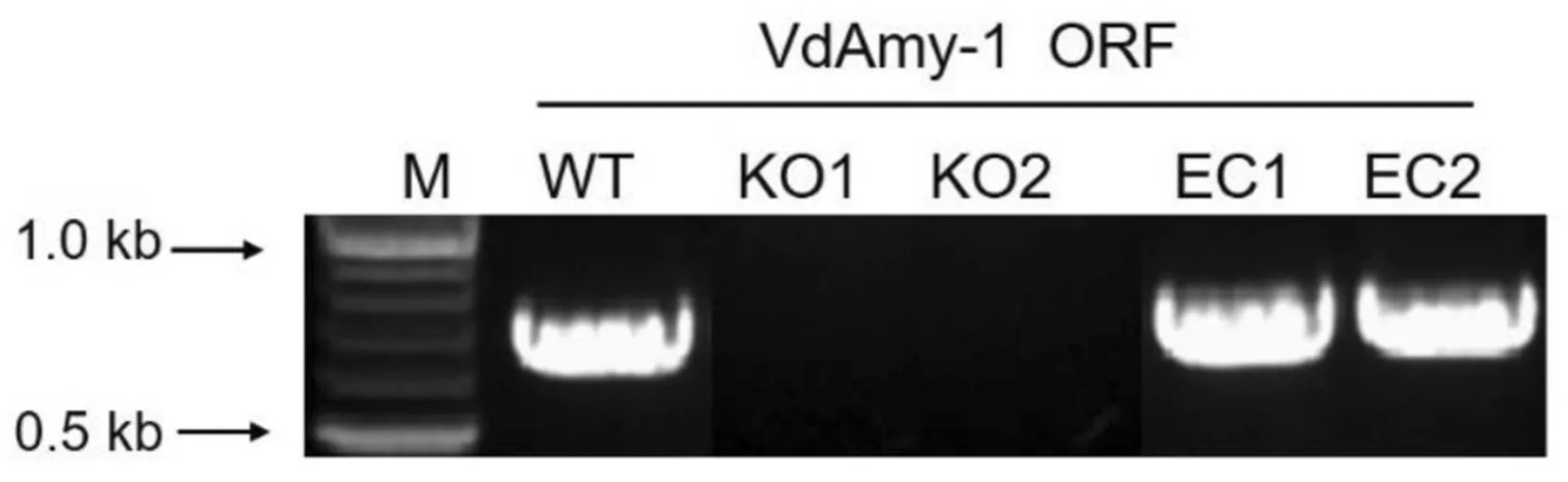
图2 大丽轮枝菌突变体菌株(KO-1/2)和功能回补菌株(EC-1/2)的构建
2.4 突变体菌株利用淀粉能力减弱
野生型菌株WT和突变体菌株KO-1/2在丰富培养基PDA上均生长良好,可形成大量的白色气生菌丝,无黑色微菌核生成,并且形成大小相似的菌落.说明VdAmy-1的缺失不影响大丽轮枝菌的基本生长能力(图3).与之相比,突变体KO-1/2在以淀粉为唯一碳源的培养基上的菌落直径明显小于野生型菌株,仅有野生型菌株的52%.功能互补菌株EC-1/2可弥补突变体菌株的上述缺陷(图3).
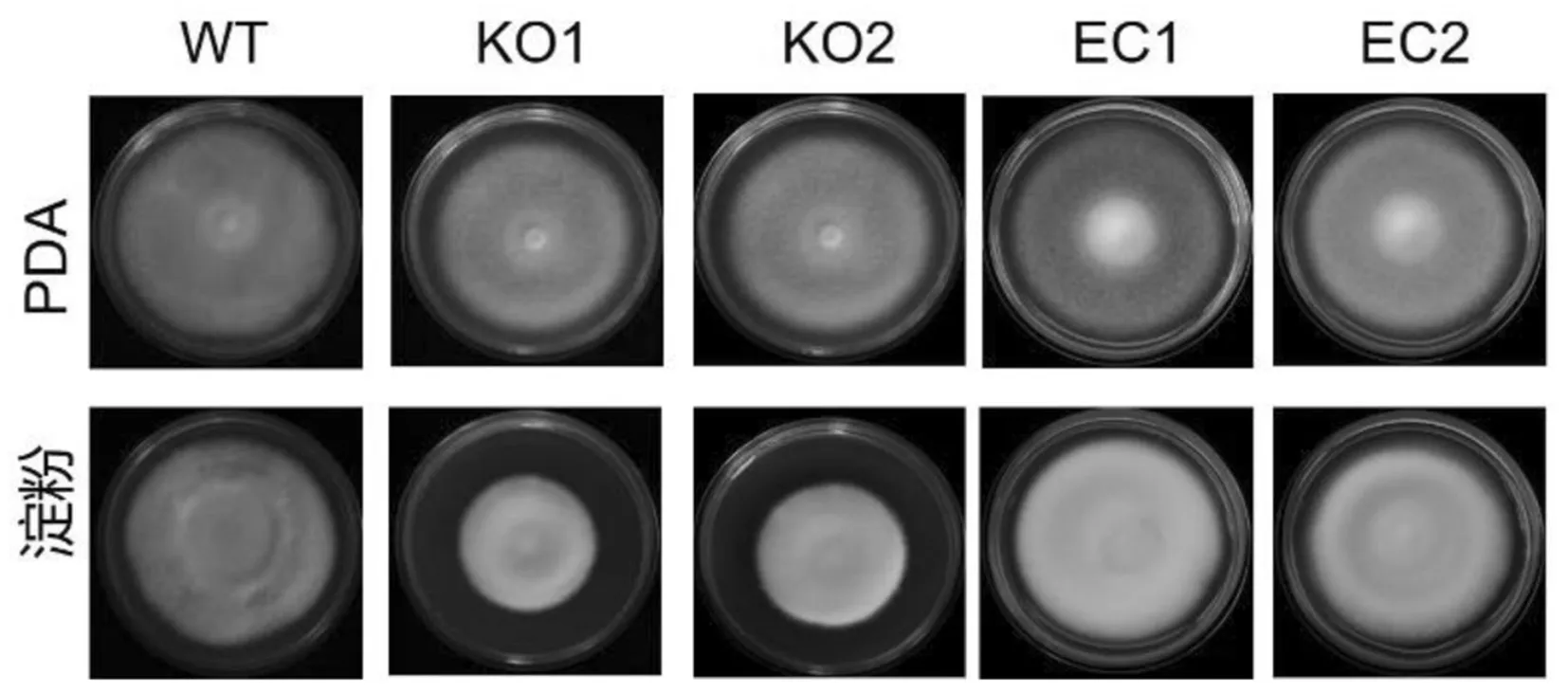
图3 VdAmy-1的缺失减弱菌株利用淀粉能力
以上实验说明突变体KO-1/2利用淀粉能力明显减弱,VdAmy-1是大丽轮枝菌重要的淀粉酶.同时,突变体菌株并没有完全丧失在淀粉培养基上的生长能力,提示大丽轮枝菌中还可能存在其他的淀粉酶编码基因,它们共同行使利用淀粉的功能.
2.5 突变体菌株致病性下降
致病性实验表明,接种野生型菌株的棉株表现出典型的叶片萎蔫、褪绿、坏死和植株株高变矮的黄萎病发病症状.与之相比,突变体菌株所侵染的棉苗发病明显减轻,仅表现出微弱的黄萎病症状.病情指数统计显示,突变体菌株造成的致病性仅有野生型菌株的41%.回补菌株所侵染的棉苗的发病症状与野生型相似(图4).以上结果说明淀粉酶VdAmy-1是大丽轮枝菌重要的致病因子.推测原因,可能是由于淀粉是病原菌重要的碳源,淀粉利用能力的下降可能造成大丽轮枝菌在寄主中的生长受限,从而使菌株致病性下降.

图4 VdAmy-1的缺失减弱菌株的致病性
3 结 论
VdAmy-1基因的缺失不影响大丽轮枝菌的基本生长,但是减弱了其利用淀粉的能力,降低了对寄主棉花的致病性.上述研究表明VdAmy-1是大丽轮枝菌一个重要的分泌型淀粉酶,扮演毒力因子的角色.本研究进一步揭示了大丽轮枝菌如何利用寄主的营养物质,为其生物防控提供了理论基础.
