包装冷却肉中微生物腐败及其挥发性气味的研究进展
2021-01-20杨啸吟张一敏梁荣蓉朱立贤
杨啸吟,张一敏,梁荣蓉,朱立贤,罗 欣
(山东农业大学食品科学与工程学院,山东 泰安 271018)
肉与肉制品具有较高的营养价值、水分活度(≥0.85)以及适宜的pH值(5.5~6.5),使其极易发生微生物腐败,这在很大程度上造成了全球每年超过20%的肉品浪费,相当于7 500万 头肉牛的产肉量[1]。肉的腐败变质通常伴随异味、变色、发黏等现象,其中微生物生长代谢所产生的挥发性有机物(volatile organic compounds,VOCs)是导致肉品腐败异味的重要来源。
包装气体成分和温度是影响微生物生长的最重要外在因素,不同气体成分不仅会影响冷却肉中的菌群演替过程,还会使细菌发生不同类型的代谢活动,从而最终决定了肉品腐败时所释放的VOCs种类和含量[2]。包装肉的腐败通常是由某些占据种群优势的特定腐败菌(specific spoilage organisms,SSOs)引发,这些细菌代谢产生的醇类、醛类、酮类、硫化物、酯类和有机酸是肉品呈现异味的主要VOCs,也是标记肉品贮藏期间气味变化的重要化学标签[3]。因此,系统研究不同包装冷却肉的腐败气味成分,从菌群结构上阐释其特定VOCs形成的外在原因,并从底物利用和代谢途径角度分析SSOs产生VOCs的内在原因,对利用特征气味分子观测肉品腐败进程以及抑制肉品腐败具有指导意义。
1 不同包装冷却肉的微生物群落及其生成的VOCs
1.1 透氧托盘包装
鲜切肉中的初始微生物主要源自动物的皮毛、粪便、内脏,以及屠宰加工环境和人为操作的交叉污染,其中不动杆菌、假单胞菌、索丝菌、黄杆菌、嗜冷杆菌、梭状杆菌、莫拉氏菌、葡萄球菌、微球菌、乳酸菌和肠杆菌是最常见的污染菌,检测到的初始VOCs主要有己醛、1-辛烯-3-醇、2-丁酮、2,5-辛二酮等[4-5]。
由于透氧托盘包装内的气体成分始终与空气较为相似,在这种低温有氧环境下,一旦菌落总数超过8(lg(CFU/cm2))就会导致肉品产生异味。乙偶姻(3-羟基-2-丁酮)、二乙酰(2,3-丁二酮)、2-乙基己醇、3-甲基-1-丁醇、2-丁酮、2-甲基丁酸、乙酸乙酯等是典型的VOCs,这主要归因于假单胞菌的生长代谢[5-7],这些物质可以作为假单胞菌引发托盘包装肉品腐败气味的关键VOCs。常见的假单胞菌菌种有莓实假单胞菌、荧光假单胞菌、隆德假单胞菌和恶臭假单胞菌,莓实假单胞菌和恶臭假单胞菌多为牛肉中的优势腐败菌,而猪肉和禽肉中莓实假单胞菌和荧光假单胞菌则更为常见[8-9]。莓实假单胞菌生长过程中会释放大量甲酯、乙酯(约占总酯类82%)、2-乙基己醇、丁醇、3-甲基-1-丁醇等VOCs,荧光假单胞菌生长过程中会产生戊酮、辛酮、乙偶姻、丁醇等VOCs,隆德假单胞菌生长过程中则会释放大量1-十一碳烯、5-甲基-2-己酮和甲基-2-丁烯酸等VOCs以及氨气,而恶臭假单胞菌生长过程中会产生二硫化碳和2-丁酮、辛酮等酮类VOCs[10-13]。另外,热杀环丝菌也是该包装下的优势腐败菌,伴随着它们的生长,肉中的2-甲基丙醇、乙偶姻、二乙酰、2,3-丁二醇、2-甲基丁酸和3-甲基丁酸等VOCs含量会显著增加,其中两种甲基丁酸具有酸臭味,是热杀环丝菌有氧繁殖的特殊代谢产物[3,14-15]。表1为不同包装冷却肉中的常见细菌种类。

表1 不同包装冷却肉中的常见细菌种类[16]Table 1 Bacterial species commonly found in chilled meat under different packaging conditions[16]
1.2 真空包装
真空包装促进了厌氧菌和兼性厌氧菌的生长,乳酸菌(弯曲乳杆菌、清酒乳杆菌、低温乳杆菌、明串珠菌等)、肠杆菌(液化沙雷菌、水生拉恩氏菌、蜂房哈夫尼菌等)、热杀环丝菌、梭菌和腐败希瓦菌是真空包装牛肉、猪肉、羊肉、禽肉的常见腐败菌(表1)。在真空包装牛肉贮藏后期,由于乳酸菌(乳杆菌、乳球菌和肉食杆菌)、梭状芽胞杆菌(奥吉氏梭菌和腐化梭菌)以及肠杆菌(沙雷氏菌和蜂房哈夫尼菌)的快速增殖,产生了大量CO2以及醇类(丁醇、3-甲基-1-丁醇、2,3-丁二醇等)、醛类(己醛、庚醛、苯甲醛等)等VOCs,进而引起包装胀袋现象[17]。Mansur等[6]发现丁酸、乙酸和2-丁酮是真空包装牛肉腐败进程中的代表性VOCs,它们与低温乳杆菌的生长密切相关。进一步研究发现戊酸、乙醇、3-甲基-1-丁醇和2,3-丁二醇也可成为真空包装牛肉腐败的重要标记物[18]。值得注意的是,丁酸含量与乳酸菌和梭菌数量都呈显著正相关,而这两种细菌是导致真空包装牛肉后期腐败的优势菌[6-7,17],因此可把丁酸作为该包装体系下肉品腐败的潜在特征VOCs。采用Nisin活性真空包装则可以有效抑制乳酸菌的生长,在这种包装环境下肉食杆菌生长所释放的丁酸、己酸和庚酸这些羧酸类物质是牛肉酸败气味的主要来源[19]。不同于透氧袋装羊肉所检测到的大量己醛、庚醛和壬醛物质(表2),真空无氧环境促进了乳酸菌的繁殖,其代谢产生的3-甲基丁醛和乙偶姻是真空包装羊肉腐败异味的重要VOCs[20-21]。此外,腐败希瓦菌的生长不仅会产生H2S等恶臭气味物质,还会造成肉品绿变[22]。
1.3 气调包装
1.3.1 高氧气调包装
高氧气调包装(体积分数6 0%~8 0% O2配合20%~40% CO2)可赋予红肉诱人的鲜红色,是目前最流行的气调包装方式。热杀索丝菌和假单胞菌通常是该包装冷却牛肉的SSOs,而且假单胞菌在后期表现出比热杀环丝菌更好的环境适应性,其中乙偶姻、2,3-丁二醇、辛酸乙酯和己酸这些VOCs与热杀环丝菌的代谢密切相关[7,28]。但有学者却发现乳酸菌才是引发高氧气调包装牛肉腐败的主要原因,这在高氧气调包装猪肉上也得到了验证,贮藏前10 d冷生明串珠菌一直是优势菌种,到第14天肉品腐败时鱼乳球菌则发展为SSOs,此过程中乙醇、乙酸、二乙酰和乙偶姻的含量随腐败加剧而快速升高[25,29]。同样,Jääskeläinen等[2]也报道冷生明串珠菌和伴气明串珠球菌是造成高氧包装牛肉黄油异味的SSOs,它们生长过程中会伴随二乙酰、乙偶姻、壬醛、1-辛烯-3-醇和己酸含量的显著增加。鉴于高氧气调包装肉的菌群结构差异是导致出现不同VOCs组成结果的一个重要原因,因此有必要对该包装下的菌群结构和VOCs变化开展进一步研究,但也发现乙偶姻和己酸是不同菌群产生的共同VOCs,可作为未来预测该包装牛肉腐败程度的潜在标记物。

表2 导致不同包装冷却肉异味的醇类、醛类、酮类挥发物种类及优势腐败菌Table 2 Alcohols, aldehydes, and ketones produced by spoilage bacteria during storage of chilled meat in different packages

续表2
除热杀索丝菌和乳酸菌外,Nieminen等[23]在高氧气调包装猪肉SSOs中分离到了发光杆菌,特别是明亮发光杆菌,该细菌的增殖导致乙偶姻、双乙酰和3-甲基-1-丁醇这些异味分子的产生。Jääskeläinen等[26]则在接种了伴气明串珠球菌的高氧包装猪肉中检测到乙偶姻、双乙酰、2,3-丁二醇、乙酸、己醛、庚醛的快速产生,这些VOCs赋予猪肉黄油异味。另有学者在高氧气调包装鸡肉中接种了荧光假单胞菌,发现贮藏10 d内烃类物质最多(17 种)、其他依次为醛类(8 种)、硫化物(6 种)、酯类(6 种)、酮类(4 种)和醇类(3 种),其中戊-1-烯、二甲硫醚、乙偶姻、乙酸乙酯和硫代乙酸甲酯是荧光假单胞菌代谢的主要VOCs[27]。而对接种热杀环丝菌的高氧气调包装鸡肉,二乙酰、二甲硫醚、乙偶姻、3-甲基-1-丁醇和乙酸是该包装体系下的特征VOCs[24]。鉴于同一包装条件下不同菌种生长过程中会产生相应的挥发性指纹标记物,而挥发物组学方法目前已在肉品掺假鉴定上得到成功应用[30],因此能否借助该技术对包装肉品中的不同SSOs进行持续监测值得深入研究。
1.3.2 低氧气调包装
低氧气调包装(O2≤30%(体积分数,下同))可赋予肉品更长的货架期,其中以O2体积分数不超10%的低氧气调包装较为典型,乳酸菌、肠杆菌和热杀环丝菌是该包装体系下的优势腐败菌[31]。但近期研究结果发现:低氧气调包装被认为会限制假单胞菌这种严格好氧菌的生长,然而部分韦氏假单胞菌、莓实假单胞菌和隆德假单胞菌在无氧环境中生长良好,且能抵御高体积分数的CO2,其产生的VOCs是无氧气调包装牛肉腐败气味的重要来源[32]。即便是乳酸菌,也不一定能适应这种低O2高CO2浓度的贮藏环境,Nieminen等[33]指出80% O2有助于猪肉中明串珠菌的生长,无氧环境则促进了乳杆菌的生长,至于CO2较少的无氧包装则会加剧乳球菌的生长。
一氧化碳气调包装(69.6% N2/0.4% CO/30% CO2)是常用的低氧气调包装方式,研究发现十二烷、癸醛和乙偶姻是该包装牛肉中最典型的VOCs[34]。而在40% CO2/30% O2/30% N2包装牛肉贮藏期间,热杀环丝菌和乳酸菌是优势菌群,它们代谢产生的乙酸和2-羟基丙酸加剧了肉品酸败气味的产生,且己醇和乙酸乙酯快速增加[5,35]。对于80% CO2/20% N2包装鸡肉而言,气单胞菌、乳球菌、沙雷菌和希瓦菌则是导致其贮藏后期产生异味的优势腐败菌[8]。Tománková等[36]通过比较70% Ar/30% CO2和高氧气调包装鸡肉的VOCs差异,发现除氨气和五甲基庚烷这两种共同VOCs外,二甲硫醚、二甲二硫醚、二甲三硫醚和H2S是Ar包装鸡肉特有的VOCs(表3),这与该包装下会产生更多的乳酸菌和大肠菌群有关。
2 不同包装肉中微生物生成VOCs的底物
2.1 底物种类
不同包装冷却肉腐败散发的VOCs不仅取决于微生物的种类,还在很大程度上取决于微生物代谢所利用的底物种类和含量。碳水化合物是绝大多数细菌生长所需的首要营养物质,一旦肉中的葡萄糖被耗尽,细菌转而利用肉中或自身代谢产生的乳酸、葡萄糖酸、6-磷酸葡萄糖、丙酮酸、乙醇、乙酸盐等物质,后续还将分解利用核酸以及蛋白质、脂肪中的氨基酸、脂肪酸和甘油。不同包装下的优势菌在此过程中利用的底物不同,因而产生的代谢物也不同,通常有氧气调包装肉的腐败气味比无氧气调包装肉更难闻[37]。葡萄糖是细菌产生3-甲基丁醇、2-甲基丙醇、乙醇、乙酸、丁酸、2-甲基丁酸、乙偶姻和二乙酰的前体物质,正是由于糖代谢对乳酸菌生长至关重要,因此在真空包装肉中才能检测到比托盘包装肉更多的醇类和羧酸类物质[3,37];游离氨基酸则是某些酶促脱羧脱氨作用的底物,可生成硫化物、支链脂肪酸、甲酯和氨等,而脂肪被细菌酶解后所释放的挥发性脂肪酸,以及脂肪酸代谢产生的醛类、酮类、酯类等物质同样是肉品腐败气味的重要成分[4]。
2.2 底物利用顺序
不同包装中细菌对底物的利用顺序也不同。假单胞菌在有氧气调包装肉中利用的底物依次为葡萄糖、葡萄糖-6-磷酸、乳酸、丙酮酸、葡萄糖酸、葡萄糖酸盐和氨基酸,而在无氧气调包装中则是葡萄糖、葡萄糖-6-磷酸、丙酮酸或葡萄糖酸盐、乙酸、氨基酸,并不能利用乳酸[22,37]。由于假单胞菌分泌的外源酶拥有更强的蛋白质和脂质水解能力,使得它们能更快地渗入肉品内部获得更丰富的能量来源,从而在有氧条件下表现出比其他细菌更强的种群优势和腐败潜力,此过程会释放大量的2-壬酮、2-庚酮、乙酸、3-甲基-1-丁醇、2-乙酸辛烯酯[38]。热杀环丝菌是兼性厌氧菌,在有氧包装或葡萄糖稀缺的肉中会通过异型发酵依次利用葡萄糖、葡萄糖-6-磷酸、氨基酸、核糖和甘油,乙偶姻就是该菌高氧气调包装下进行葡萄糖代谢的终端产物,而典型的2-甲基丁酸和3-甲基丁酸则分别来自其对异亮氨酸和亮氨酸的降解;在无氧或葡萄糖充足环境下,热杀环丝菌则只能通过同型发酵将葡萄糖和葡萄糖-6-磷酸转化为甲酸、乳酸和乙酸这些异味物质[4,22,37,39]。
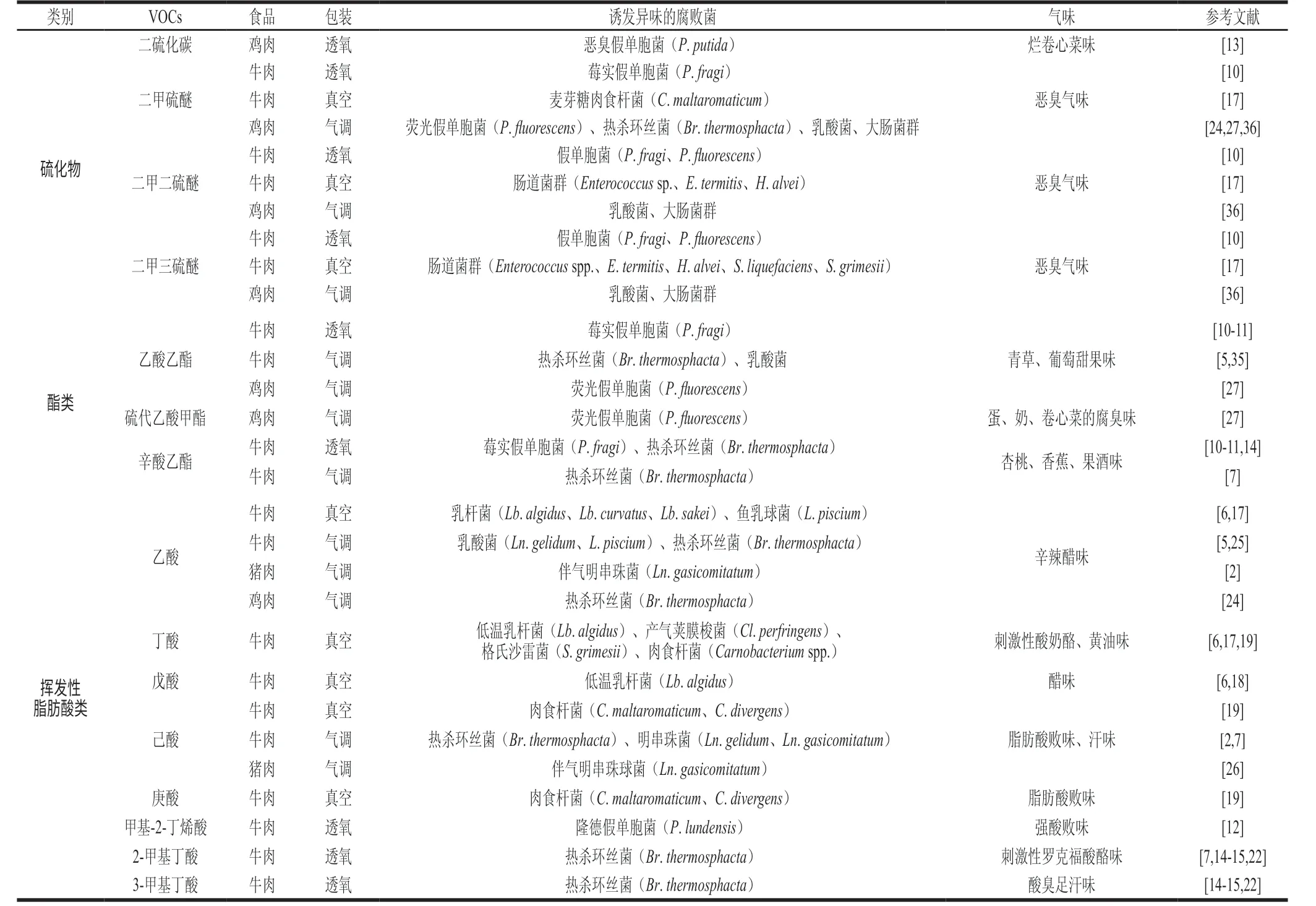
表3 导致不同包装冷却肉异味的硫酸物、酯类、脂肪酸挥发物种类及优势腐败菌Table 3 Sulfur compounds, esters, and fatty acids produced by spoilage bacteria during storage of chilled meat in different packages
无论有氧还是无氧包装,乳酸菌都先利用葡萄糖和葡萄糖-6-磷酸进行代谢,不同的是,无氧包装后期乳酸菌还能利用肉中的氨基酸,某些清酒乳杆菌和肠膜明串珠菌就可将组氨酸、酪氨酸、精氨酸、赖氨酸水解成氨气和组胺、酪胺、精胺、尸胺等生物胺[40]。相较于真空包装,高氧包装促进了乳酸菌的异型发酵,生成乳酸、乙酸、CO2,并伴随着乙醇和乙醛等中间产物的产生,而且一旦葡萄糖耗尽,高浓度的CO2还会促使乳酸菌以乳酸为底物将其氧化为乙酸[2,41-42]。与乳酸菌类似,肠杆菌生长也是先利用葡萄糖和葡萄糖-6-磷酸,在有氧气调包装后期会利用乳酸和氨基酸,而在无氧气调包装中则利用乙酸。无氧包装中,液化沙雷菌和蜂房哈夫尼菌等肠杆菌可将葡萄糖发酵为含有乙酸、甲酸、乳酸、琥珀酸和乙醇的混合物,一旦它们的数量达到6~7(lg(CFU/g)),就会分解氨基酸产生大量硫化物、胺类和支链酯类,散发出让消费者难以接受的气味,而且这些肠杆菌在高pH值的异质肉中更容易生长[43]。作为造成真空包装肉品异味的另一大腐败菌——梭菌(奥吉氏梭菌、腐化梭菌、产气荚膜梭菌等)则通过代谢葡萄糖和葡萄糖-6-磷酸来生成乙醇、3-甲基-1-丁醇和丁酸等VOCs,并产生CO2和H2导致包装膨胀[17,44]。
3 不同包装肉中微生物生成VOCs的主要代谢途径
细菌为应对特定包装环境所开展的代谢方式并不相同,这将最终影响肉的腐败进程。例如气调包装会诱发乳酸菌异型发酵,通过磷酸戊糖途径释放乙醇和乙醛等异味VOCs,而牛至精油则能激发乳酸菌同型发酵,通过糖酵解生成乳酸[3,41]。从代谢类型转换角度来看,向气调包装肉中添加牛至精油将有利于抑制包装肉品的异味产生,因此了解不同包装微生物合成VOCs的代谢途径十分必要。
3.1 醇类产生途径
醇类的产生涉及氨基酸降解以及酮类、醛类的还原等多个代谢通路,因此,在不同包装肉中都能检测到此类物质,其中比较有代表性的是2,3-丁二醇、3-甲基-1-丁醇、1-辛烯-3-醇、2-乙基己醇、丁醇、己醇和1-辛烯-3-醇(表2),假单胞菌、热杀环丝菌和肉食杆菌被认为是肉中醇类物质的主要合成者[4,45]。在高氧气调包装中,2,3-丁二醇可由广布肉食杆菌和热杀环丝菌通过丙酮酸代谢途径还原乙偶姻合成(图1),而3-甲基-1-丁醇也是热杀环丝菌的有氧代谢产物,可由其水解氨基酸产生[24]。
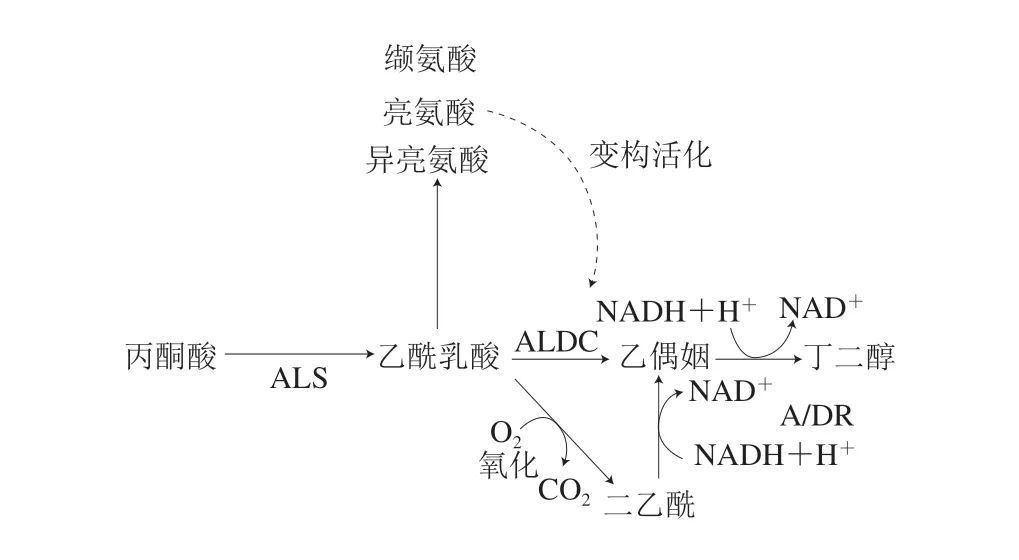
图1 细菌的丙酮酸代谢途径及相关挥发性代谢产物Fig.1 Bacterial pyruvate metabolism pathways associated with volatile metabolites
3.2 醛类产生途径
醛类的产生主要源于三酸甘油酯的水解、不饱和脂肪酸β-氧化或脂质的自动氧化,以及氨基酸转氨作用等途径,因此,醛类(尤其是短链醛类物质)通常具有脂肪酸败的气味,假单胞菌、热杀环丝菌、肉食杆菌和肠杆菌是肉中醛类物质的主要贡献者[4,46]。己醛、壬醛、庚醛、苯甲醛和3-甲基丁醛是最常见的醛类,在不同包装肉中都有检出(表2)。作为衡量包装牛肉货架期的重要指标,己醛、壬醛等醛类物质可通过亚油酸等不饱和脂肪酸的氧化代谢产生[2,34]。甲基丁醛则是乳酸菌利用支链脂肪酸(如亮氨酸)生成3-甲基-1-丁醇、2-甲基-1-丁醇或2-甲基丙烷的中间产物[47]。
3.3 酮类产生途径
酮类可以来自脂肪酸的自动氧化以及酶促α-或β-氧化,也可以通过革兰氏阴性菌水解脂质、降解烷烃以及对醇类脱氢完成,酮类物质在以假单胞菌、热杀环丝菌、乳酸菌、肠杆菌为主的包装肉中都能检出,其中乙偶姻和二乙酰是不同包装肉腐败所产生的最重要酮类物质[5]。在细菌的酶促丙酮酸代谢通路中,丙酮酸先被转化为乙酰乳酸,经进一步氧化后生成二乙酰,而二乙酰则可被二乙酰还原酶还原为乙偶姻[39](图1)。
3.4 硫化物产生途径
无论是在有氧还是无氧包装肉中,微生物(假单胞菌、肠杆菌、乳酸菌等)降解含硫氨基酸(甲硫氨酸和半胱氨酸)都会释放硫化物,甲硫氨酸代谢是这些细菌产生二甲硫醚、二甲二硫醚和二甲三硫醚等常见硫化物的主要途径:甲硫氨酸先被分解为甲硫醇,再经氧化后成为具有恶臭味的高分子硫化物[22]。
3.5 酯类产生途径
乙酸乙酯、硫代乙酸甲酯、丁酸乙酯和辛酸乙酯是包装肉中比较常见的挥发性酯类,赋予了腐败肉水果异味,其中莓实假单胞菌、荧光假单胞菌和热杀环丝菌被发现是有氧气调包装肉中酯类物质的重要制造者(表3)。这些酯类一方面来自细菌自身合成醇类和羧酸类物质的酯化作用;另一方面也可通过醇类与摄入脂肪酸的酯化合成[48]。
3.6 挥发性脂肪酸产生途径
导致肉品酸败异味的VOCs多为一些挥发性脂肪酸,它们可通过微生物碳水化合物代谢(氧化酮、醛、酯类)或水解脂质和氨基酸等途径生成[49]。乙酸、丁酸和己酸这些直链脂肪酸在不同包装肉中都有发现,而甲基丁酸这类支链脂肪酸则只在有氧包装肉中才被发现。乙酸是乳酸菌异型发酵的重要代谢产物,而在真空包装肉中,梭菌则会分别利用葡萄糖或氨基酸为底物进行丁酸发酵或Stickland反应并最终生成丁酸[50]。此外,肉食杆菌和热杀环丝菌的代谢也会释放大量挥发性脂肪酸(表3)。
4 生鲜肉微生物腐败气味的检测与鉴定
4.1 色谱串联质谱法
气相或液相色谱串联质谱法是目前最常规的VOCs检测手段,以顶空固相微萃取-气相色谱-质谱联用法最为常用,学者们用该技术对不同包装贮藏牛肉、猪肉、羊肉、禽肉的VOCs进行了大量研究[8,21,29,35]。传统色谱-质谱法检测样品需要长时间的前处理,这导致VOCs的测定被间隔化,所提供信息不能及时反映样品的原有状态。针对上述弊端,质子转移反应质谱技术应运而生,可实现肉品VOCs的实时监测,但此方法仍存在同分异构体分子难以区分的局限性,未来气相色谱技术如果能与质子转移反应飞行时间质谱技术相结合将能有效解决此问题[24]。
4.2 电子鼻技术
电子鼻利用传感器可将气味分子的化学信号转化为电信号,从而通过模拟人的嗅觉来描述食物的复杂气味,该技术能够依据样品的整体气味对肉品腐败程度进行高效精确评估,而且还能区分不同贮藏条件下不同物种肉的气味特征,具有很好的实用价值。但是当前的电子鼻技术还存在传感器寿命短、不能精确定性定量分析特征VOCs等缺点,这都很大程度上制约了其大规模推广[51]。未来电子鼻结合生物传感器实现VOCs的精确识别将是该技术的一大发展方向。
4.3 光谱技术
基于光谱识别的微生物代谢物无损快检技术是食品科学领域的前沿热点,以傅里叶变换红外光谱、拉曼光谱和高光谱成像技术为主流。高通量傅里叶变换红外光谱具有灵敏度、精度、分辨率高和波段宽等特点,Kosa等[52]采用该技术成功检测到产油真菌中的生物聚合物、多糖、脂肪酸等代谢物。拉曼光谱受水分子影响较小,因此更适合分析复杂环境下的细菌代谢物,Morelli等[53]就利用表面增强拉曼散射方法精确定量了大肠杆菌分泌的香豆酸和肉桂酸。高光谱成像技术目前已被用于监测藻类代谢物的变化积累,而该技术对腐败菌VOCs的鉴定则还有待深入研究[54]。
5 结 语
不同包装环境会改变肉的菌群结构,并激发SSOs启动相应的代谢通路,促使肉品腐败时释放出具有迥异气味的VOCs,而且不同包装优势菌的底物利用与代谢类型也会直接影响肉品VOCs的组成和含量,这能够最终决定肉品的腐败程度。因此,掌握特定包装肉中不同腐败菌的演替过程及其潜在VOCs标记物的种类和合成机制意义重大,将有利于采取靶向措施调控和监测肉品微生物腐败进程。
为抑制肉品腐败气味的产生,未来可基于不同包装SSOs的生长特性开发新型活性包装:通过缓释特定的天然植物提取物来改善菌群结构,并调节肉中SSOs的代谢途径,使其朝可产生更弱异味VOCs的方向发展。鉴于微生物脱羧生化反应是其生成异味VOCs的重要途径,而CO2作为该代谢过程的重要产物会抑制脱羧反应的进行[55],因此,通过开发具有CO2缓释功能的活性包装来抑制肉品异味也将成为可能。另外,随着食物组学技术的快速发展,可将基因组学、转录组学、蛋白质组学、代谢组学等手段与挥发物组学进行有机结合,从而高通量精确识别不同包装肉的特征腐败气味分子,探寻出更多能够标记肉品腐败的指纹挥发物,并在此基础上利用大数据、生物电子传感器等新兴技术综合开发可用于预测肉品货架期的智能包装系统,最终实现对冷却肉全链条流通的实时监控。
