外源乙烯的处理时机对甘薯抑芽及碳水化合物代谢的影响
2021-01-20段志蓉靳苗苗吴姗鸿代慧张敏
段志蓉,靳苗苗,吴姗鸿,代慧,张敏*
1(西南大学 食品科学学院,重庆,400715)2(西南大学 食品贮藏与物流研究中心,重庆,400715) 3(农业部农产品贮藏保鲜质量安全风险评估实验室(重庆),重庆,400715)
近年来,鲜食甘薯作为最常见的粗粮食品之一受到消费者的喜爱[1-2]。然而甘薯无生理休眠期,在16~35 ℃的环境中很容易发芽,采后发芽会导致块根内部营养物质迅速转移消耗于芽生长,使甘薯品质下降,寿命缩短,商品性受到破坏。而我国大部分销售流通环境又正好处于这个易发芽的温度区间,所以采用有效的抑芽手段十分必要。
乙烯作为一种天然气体激素存在于大部分植物中,根据激素平衡调节的原理,利用外源乙烯改变内源植物激素的平衡,可以延长休眠、抑制发芽。DANIELS-LAKE等[3]研究表明,在一定范围内,外源乙烯的处理浓度与马铃薯的休眠抑芽时间呈正相关。此外,碳水化合物是甘薯块根中存在量较大的一类化合物,在其生理过程中起到提供能源和信号传递等关键作用[4]。就甘薯等易发芽的园艺作物而言,采后碳水化合物代谢是影响其生长、休眠和发芽的关键[5-6]。有研究表明,较高的蔗糖含量有助于抑制芽的生长[7-8]。同时,磷酸蔗糖合成酶(sucrose phosphate synthase,SPS)、转化酶和淀粉酶在调节蔗糖水平方面起到了重要作用[1]。MURKUTE等[9]研究表明,外源乙烯处理可使洋葱总糖含量处于较高水平,并推测这可能是延长洋葱休眠期的原因之一。DAI等[10]研究表明,199.3 μL/L外源乙烯处理可提升马铃薯SPS、酸性转化酶(acid invertase,AI)的活性,进而有效抑制块茎发芽。
此外,BUFLER等[11]对何时使用外源乙烯的研究表明,在洋葱开始出现发芽迹象的时间点后再用外源乙烯处理,与全程乙烯处理一样能有效抑制发芽。如果能对处理时机的影响有很清楚的了解,那么实际生产中可以在综合考虑采摘时机、管理方式、成本控制等实际情况的前提下,选择最适宜的处理时机来达到最佳效果。然而,目前有关不同外源乙烯的处理时机对甘薯抑芽及碳水化合物代谢活动影响的文献未见报道,其调控机理尚不明确。本课题组前期研究表明,使用1 g/L外源乙烯处理甘薯在维持甘薯品质和抑芽方面都能得到最佳效果,所以本研究采用1 g/L外源乙烯,以甘薯开始发芽的时间点(即薯皮出现肉眼可见的芽点)为节点,研究节点前、节点后和储运全程乙烯处理对甘薯发芽及碳水化合物代谢的影响,以探究不同外源乙烯处理时机对甘薯常温储运期间的抑芽作用,并从碳水化合物代谢角度分析其调控机理,为实际应用提供参考。
1 材料与方法
1.1 材料与试剂
选用同一批次、大小合适且相对均匀、机械损伤少、无肉眼可见芽点、无病害和虫害的“红心王”甘薯,重庆市北碚区天生农贸市场。
标准邮政6号纸箱,260 mm×150 mm×180 mm,三层B瓦楞,山东冠县万林纸制品有限公司;2.5 g/包乙烯利(有效成分20%),山东营养源食品科技有限公司;尿苷5-二磷酸葡萄糖二钠、果糖、6-磷酸果糖二钠、D-无水葡萄糖,北京索莱宝科技有限公司,其他试剂均为分析纯。
1.2 仪器与设备
UV-2450PC型紫外可见分光光度计,日本岛津公司;HH-2型数显恒温水浴锅,常州奥华仪器有限公司;FA2004型电子天平,上海恒平科学仪器有限公司;H1650R型台式高速冷冻离心机,湖南湘仪公司;RXZ-8000型智能人工气候箱,宁波东南仪器有限公司。
1.3 实验方法
1.3.1 样品制备
将采购于农贸市场的甘薯按照材料要求筛选后,随机放入纸箱后称重,每箱1500 g左右。不放乙烯利的为对照组(CK组),乙烯利浓度为1 g/L(乙烯利质量与气体空间体积比)为全程处理组(A组)。将装箱后的甘薯放入气候箱中,模拟常温储运环境,(25±1)℃、相对湿度(90±5)%下贮藏18 d,每天观察发芽情况,观察到CK组甘薯出现发芽迹象时,将CK组中的一半甘薯取出,放入乙烯利1 g/L的纸箱后再封箱,为节点后处理组(B组);将A组中的一半甘薯取出,通风0.5 h放入无乙烯利的纸箱内再封箱,为节点前处理组(C组)。每3 d取样测定各项指标,每组随机取样3箱。
1.3.2 发芽率和发芽指数的测定
甘薯皮出现肉眼可见的芽点即被视为发芽[12],记录发芽甘薯个数和甘薯总数,同时参考PANKOMERA[13]的方法,观察每个甘薯的发芽数,记录为发芽级数:1级= 0芽;2级= 1~3芽;3级= 4~6芽;4级= 7~9芽;5级= 10芽及以上。按照公式(1)和(2)分别计算甘薯的发芽率和发芽指数:
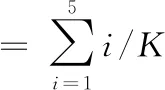
(1)

(2)
式中:i,发芽级数;K,甘薯总数,个。
1.3.3 呼吸强度的测定
参照曹建康等[14]的静置法,以mg/(kg·h)表示。
1.3.4 淀粉含量的测定
按GB 5009.9—2016[15]中酸水解法测定,以g/100 g表示。
1.3.5 蔗糖含量的测定
按GB 5009.8—2016[16]中酸水解-莱因-埃农氏法测定甘薯的蔗糖含量,以g/100 g表示。
1.3.6 还原糖和可溶性总糖含量的测定
按GB 5009.7—2016[17]中直接滴定法测定,以g/100 g表示。
1.3.7 α-淀粉酶和β-淀粉酶活性的测定
参照曹建康等[14]的方法,测定甘薯的α-淀粉酶和β-淀粉酶活性,酶活性以mg/(min·g)表示。
1.3.8 SPS、AI、中性转化酶(neutral invertase,NI)活性的测定
参考王惠聪等[18]的方法,测定甘薯的SPS、AI、NI活性,酶活性以mg/(g·h)表示。
1.4 数据分析
使用Microsoft Excel 2007计算各指标数据;IBM SPSS Statistics 22对指标进行显著性分析,P<0.05表示差异显著,P<0.01表示差异极显著;采用Origin Pro 8.6制图。
2 结果与分析
2.1 不同外源乙烯处理时机对甘薯发芽率和发芽指数的影响
尽管发芽的甘薯无毒,但发芽会使甘薯的呼吸强度增强,从而促进淀粉分解为小分子糖类,供给芽的生长,加速糖类的消耗导致块根品质急剧下降,进而失去食用价值[19]。如图1所示,CK组在3 d时观察到发芽迹象,6 d时发芽率高达(53.33±4.71)%,与处理组差异极显著(P<0.01)。A组直到18 d才观察到发芽现象,说明1 g/L乙烯利全程处理能有效推迟甘薯的初始发芽时间。节点后处理组(B组),在18 d内保持了较低的发芽率,且发芽指数无显著增长,与CK组差异极显著(P<0.01),说明节点后1 g/L乙烯利处理同样能够起到有效的抑芽作用。节点前处理组(C组),储运12 d时观察到发芽现象,与CK组相比,将甘薯发芽时间延迟了9 d,但开始发芽后芽苗迅速生长,18 d时发芽率和发芽指数分别高达(86.67±4.71)%、(2.77±0.29),与CK组差异不显著(P>0.05)。可见,节点前乙烯处理虽能将甘薯发芽时间延迟9 d,但开始发芽后芽苗迅速生长;节点后乙烯处理组发芽指数全程无明显增长;全程乙烯处理能将甘薯发芽时间延迟15 d。
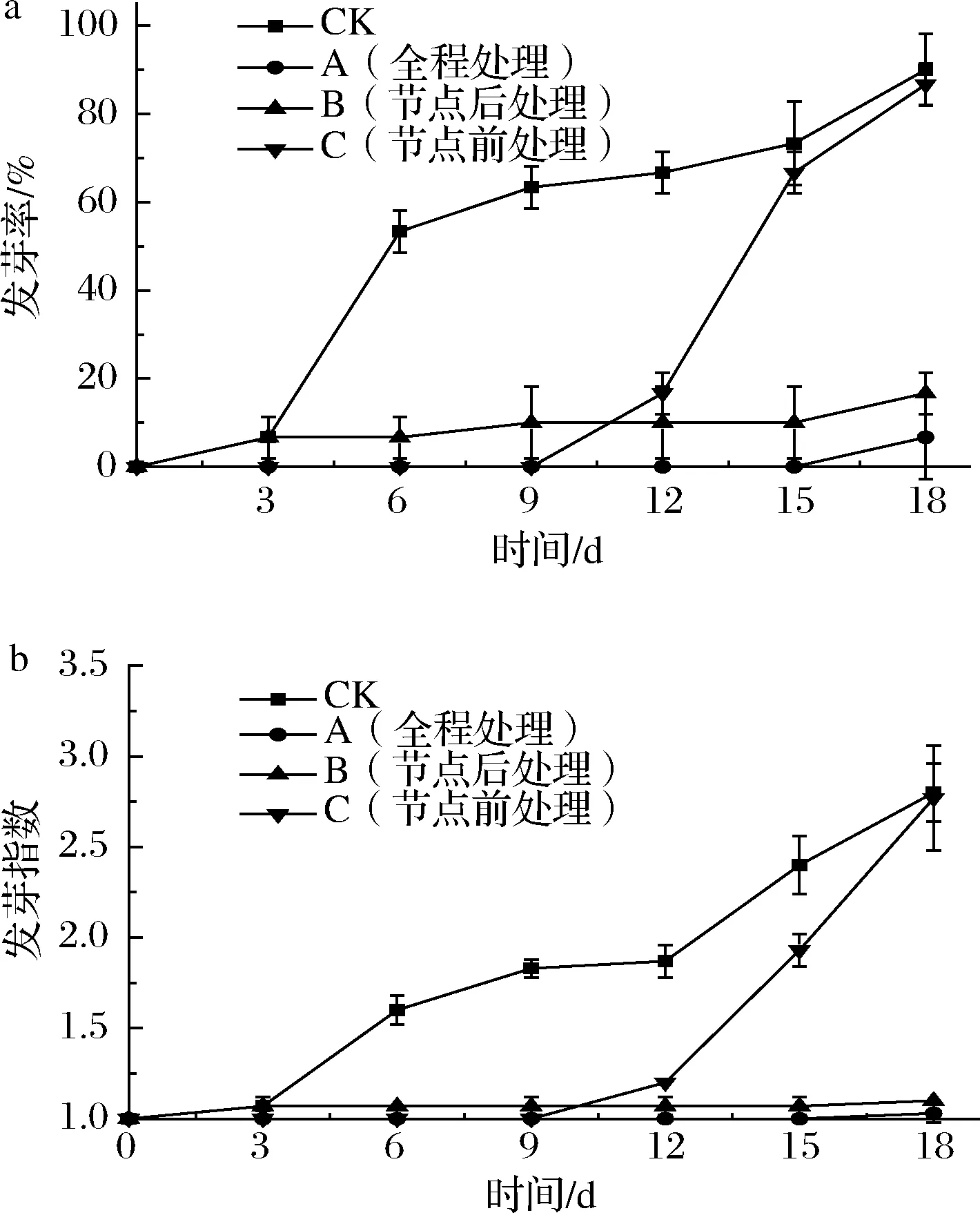
图1 不同外源乙烯处理时机对甘薯发芽率(a) 和发芽指数(b)的影响Fig.1 Effect of different exogenous ethylene treatment on germination rate (a) and germination index (b) of sweet potato
2.2 不同外源乙烯处理时机对甘薯呼吸强度的影响
呼吸作用的强度在极大程度上决定了甘薯采后的生理代谢强度,包括碳水化合物代谢程度。如图2所示,各组甘薯呼吸强度在0~3 d均有不同程度的上升。CK组在6 d后有一个先下降又上升的趋势。这是因为甘薯后期开始发芽增强了呼吸强度[20-21]。A组、C组呼吸强度在第3 天迅速升高至峰值(49.32±3.61)mg/(kg·h),比CK组峰值高出29.76%。C组在甘薯出现发芽迹象取出乙烯利后呼吸强度迅速降低,但在12 d后有所回升,这是由于此时C组发芽旺盛,使得甘薯呼吸强度升高,后期略高于CK组且差异不显著(P>0.05)。B组在甘薯出现发芽迹象放入1 g/L乙烯利后呼吸强度迅速升高,9 d时达到峰值后下降,这是由于甘薯在9 d时达到了呼吸跃变峰值[20-21]。B组储运末期虽呼吸强度有所下降,但仍然处于稳定的较高水平,显著高于CK组(P<0.05),与A组差异不显著(P>0.05)。说明外源乙烯处理会增强甘薯的呼吸强度,一方面是由于乙烯结合在细胞膜上提升了其通透性,从而加速气体交换,增强氧化作用[22];另一方面是由于乙烯可以诱导呼吸酶的活性增强,也可以通过促进呼吸酶mRNA的合成来增加呼吸酶的含量,进而增大呼吸强度[23-24]。可见,所有时机外源乙烯处理均会提高甘薯的呼吸强度,全程和节点后处理组在储运末期提升程度差异不大,节点前处理组甘薯在离开外源乙烯环境后呼吸强度下降,直到块根开始发芽时,呼吸强度再次升高。
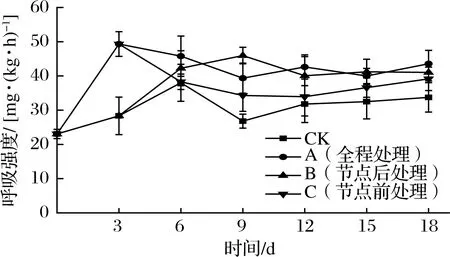
图2 不同外源乙烯处理时机对甘薯呼吸强度的影响Fig.2 Effect of different exogenous ethylene treatment on respiration rate of sweet potato
2.3 不同外源乙烯处理时机对甘薯淀粉含量的影响
植物的种子、根部和块茎中蕴藏着丰富的淀粉,甘薯中的淀粉大粒为卵形,小粒为圆形,淀粉降解是甘薯碳水化合物代谢的首要环节,淀粉降解为可溶性糖后参与到蔗糖、还原糖的转化累积以及呼吸作用中[25]。如图3所示,各组甘薯淀粉含量在前期有不同程度的上升。可能是由于环境等因素,储运前期甘薯的水分含量大幅度下降,导致淀粉相对含量增加[25];另外,可能发生的一系列复杂反应又合成了一部分淀粉[26]。储运0~3 d,CK组、B组淀粉含量显著高于A组、C组(P<0.05),这是由于外源乙烯处理增大了甘薯呼吸强度,使得乙烯处理组淀粉分解速度较快,从而淀粉含量相对较低。储运3~9 d,CK组、A组淀粉含量呈波动趋势,可能是由于甘薯块根为了保持生理代谢平衡,机体进行自我调节使得淀粉含量有所回升[26]。B组淀粉含量在3 d后下降速度逐渐加快。CK组、C组淀粉含量在9 d后下降幅度增大,这是由于此时甘薯发芽旺盛,块根消耗过多淀粉供给甘薯芽生长,导致块根淀粉加速分解。储运9~18 d,CK组淀粉含量极显著高于A组、C组(P<0.01)。在15~18 d期间CK组极显著高于B组(P<0.01)。可见所有外源乙烯处理均会加快块根淀粉含量下降速度,节点前处理组下降速度最快。

图3 不同外源乙烯处理时机对甘薯淀粉含量的影响Fig.3 Effect of different exogenous ethylene treatment on starch content of sweet potato
2.4 不同外源乙烯处理时机对甘薯可溶性总糖含量的影响
糖在甘薯块根中广泛参与到能量代谢和呼吸作用中,呈甜味的主要来源[27]。如图4所示,所有组甘薯可溶性总糖含量均先上升后下降。这是由于储运前期呼吸强度增大,加快了淀粉降解为葡萄糖等小分子糖类,之后又因芽生长被大量消耗[28]。储运0~3 d,A组、C组可溶性总糖含量显著高于CK组、B组(P<0.05)。这是由于外源乙烯处理促使淀粉水解为可溶性小分子糖,导致可溶性总糖含量明显增大[10]。CK组、C组可溶性总糖含量分别在6、12 d后明显下降;储运9~18 d,A组、B组可溶性总糖与CK组差异极显著(P<0.01),18 d时分别比CK组高出28.82%、25.14%。这是由于节点前处理组和CK组在后期发芽茂盛,消耗过多有机物所致。可见,节点后和全程1 g/L乙烯利处理均能有效延缓甘薯可溶性总糖含量的下降,使其维持较高水平,其中全程处理组效果最佳。另有研究表明,使用乙烯处理番荔枝[29]、马铃薯[5]、洋葱[30]、桃[31],其可溶性总糖含量高于未用乙烯处理组,与本研究结果相似。
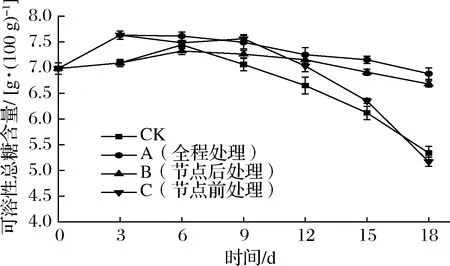
图4 不同外源乙烯处理时机对甘薯可溶性总糖含量的影响Fig.4 Effect of different exogenous ethylene treatment on soluble total sugar content of sweet potato
2.5 不同外源乙烯处理时机对甘薯还原糖含量的影响
甘薯内还原糖主要是葡萄糖、果糖和麦芽糖。储运0~3 d,A组、C组还原糖含量略微上升,与CK组、B组差异显著(P<0.05)。这是由于外源乙烯处理加速了甘薯中还原糖的生成[28]。3 d后,A组、B组还原糖含量明显下降。一方面是由于乙烯处理促使甘薯中部分还原糖转化为蔗糖;另一方面是由于还原糖参与细胞呼吸,被降解为CO2和H2O[25]。6 d时,CK组还原糖含量有所回升。这是由于甘薯块根为了准备供给发芽的营养,淀粉部分水解成还原糖后未完全立即参与代谢过程[28],还原糖在体内累积,相对含量上升。CK组、C组还原糖含量分别在6、12 d后快速下降,这是因为甘薯在此时发芽旺盛,加快了机体还原糖的消耗。18 d时,CK组显著高于各处理组(P<0.05)。可见,所有外源乙烯处理均会降低甘薯还原糖的积累。
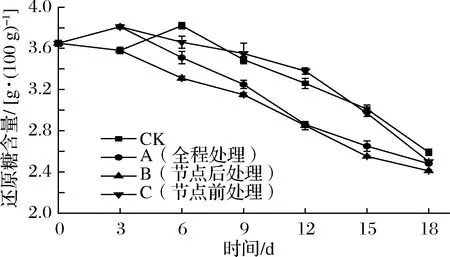
图5 不同外源乙烯处理时机对甘薯还原糖含量的影响Fig.5 Effect of different exogenous ethylene treatment on reducing sugar content of sweet potato
2.6 不同外源乙烯处理时机对甘薯蔗糖含量的影响
蔗糖是体内有机物运输的主要形式,而还原糖是光合作用的直接产物,是合成蔗糖的前体物质[32]。如图6所示,所有甘薯蔗糖含量均呈先升后降的趋势。贮藏0~3 d,A组、C组蔗糖含量上升幅度显著高于CK组、B组(P<0.05)。这是由于外源乙烯处理增大了甘薯中还原糖向蔗糖转化的速度,与郑剑英[33]的研究结果相似。CK组、C组蔗糖含量分别在6、9 d时达到峰值后持续下降,这是由于甘薯发芽旺盛,块根中部分蔗糖转化为单糖为发芽提供能量,其中C组峰值极显著高于CK组(P<0.01),但18 d时C组与CK组蔗糖含量差异不大(P>0.05)。而A组、B组蔗糖含量均在15 d达到峰值(4.5%、4.36%)后下降,18 d时分别比CK组高出59.64%、55.3%。可见,所有外源乙烯处理均会提升甘薯蔗糖含量,但节点前处理组在后期由于发芽旺盛会加速蔗糖消耗,节点后和全程处理组蔗糖含量在整个储运期间都维持在较高水平。有研究表明,较高的蔗糖含量有助于抑制芽的生长[7-8]。
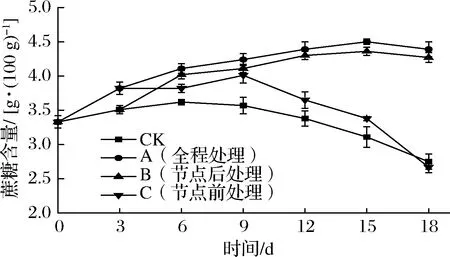
图6 不同外源乙烯处理时机对甘薯蔗糖含量的影响Fig.6 Effect of different exogenous ethylene treatment on surcose content of sweet potato
2.7 不同外源乙烯处理时机对甘薯α-淀粉酶和β-淀粉酶活性的影响
甘薯块根中的α-淀粉酶和β-淀粉酶能催化淀粉水解成麦芽糖和短支链低聚糖,其中,α-淀粉酶是作用于淀粉的α-1,4糖苷键,生成短链糊精和少量的麦芽糖和葡萄糖,β-淀粉酶是外酶,最终生成麦芽糖和限制糊精[34]。如图7所示,各组甘薯α-淀粉酶活性均呈上升趋势。储运0~3 d,外源乙烯处理组α-淀粉酶活性明显增强,与对照组差异显著(P<0.05),说明外源乙烯处理会增大甘薯α-淀粉酶活性。储运3~9 d,B组α-淀粉酶活性上升速度加快,C组α-淀粉酶活性上升趋势减缓,进一步说明离开乙烯环境会降低甘薯α-淀粉酶活性的上升幅度。CK组、C组α-淀粉酶活性分别在3、9 d后迅速上升,各组甘薯β-淀粉酶活性也呈上升趋势。储运0~3 d,外源乙烯处理组β-淀粉酶活性明显增强,与对照组差异显著(P<0.05),说明乙烯处理会增强甘薯β-淀粉酶活性。CK组、C组β-淀粉酶活性分别在第3天、第9天后迅速增强。有研究表明淀粉酶活性的增高是甘薯发芽期的正常生化反应[35]。可见所有外源乙烯处理和甘薯发芽均会诱导甘薯α-淀粉酶和β-淀粉酶活性显著增强。
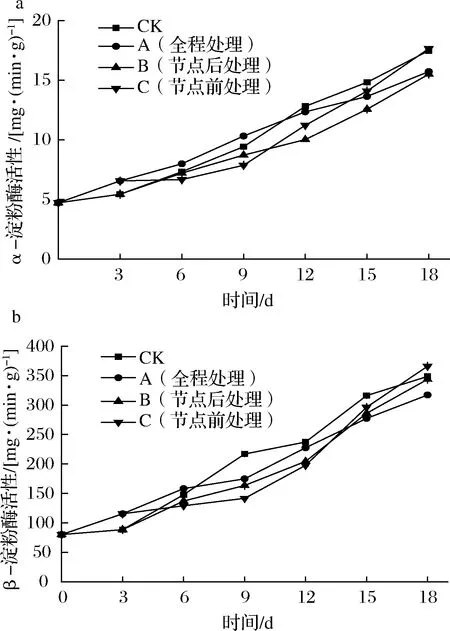
图7 不同外源乙烯处理时机对甘薯α-淀粉酶(a)和 β-淀粉酶(b)活性的影响Fig.7 Effect of different exogenous ethylene treatment on the activity of α-amylase(a) and β-amylase(b) activities in sweet potato
2.8 不同外源乙烯处理时机对甘薯SPS活性的影响
SPS是SPS-磷酸蔗糖磷酸化酶蔗糖合成系统中反应速度最慢的酶,可以调节整个途径的反应速度,因此SPS在调节蔗糖合成中起到非常重要的作用[1],有研究表明,乙烯处理可以提高菠萝蜜[36]、苹果[37]、马铃薯[5]的SPS活性。如图8所示,各组甘薯SPS活性总体均呈先升后降的趋势。CK组、C组SPS活性在第3 天达到峰值后呈波动下降。其中C组SPS活性在离开乙烯环境后明显下降,直到12 d出现了明显回升,可能是由于块根为储备更多蔗糖供给芽生长,使得SPS活性短暂性升高。A组、B组SPS活性分别于第9天、第12天达到峰值,A组、B组SPS活性峰值分别比CK组高57%、53.05%,且各外源乙烯处理组间均有显著差异(P<0.05),A组、B组SPS活性达到峰值后虽有下降,但在储运期间依然显著高于CK组、C组,说明节点后和全程乙烯处理明显增大了甘薯的SPS活性。
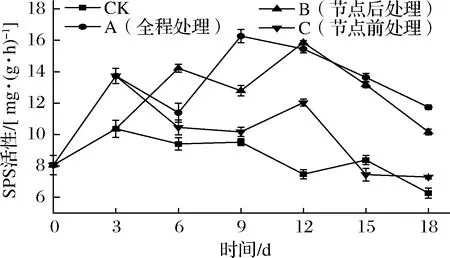
图8 不同外源乙烯处理时机对甘薯SPS活性的影响Fig.8 Effect of different exogenous ethylene treatment on SPS activity of sweet potato
2.9 不同外源乙烯处理时机对甘薯AI和NI活性的影响
转化酶根据其最适pH分为AI和NI,在蔗糖分解反应中起关键作用,分解产物为葡萄糖和果糖,且此反应不可逆[1]。如图9-a所示,各组甘薯AI活性均呈先升后降的趋势。6 d时B组的AI活性骤然下降,直到18 d,A组、B组AI活性均低于C组、CK组,差异显著(P<0.05),由此可见,节点后和全程乙烯处理会降低甘薯AI活性。而C组AI活性在第9天时达到峰值,可能是由于9 d时C组发芽旺盛,甘薯块根的蔗糖加速分解以供芽生长。如图9-b所示,各组甘薯NI活性均呈波动下降趋势。直到储运结束,尽管乙烯处理没有改变甘薯NI活性下降的趋势,但A组、B组NI活性全程高于CK组、C组,差异显著(P<0.05)。综上所述,节点后和全程1 g/L乙烯利处理会降低甘薯AI活性,增强NI活性。
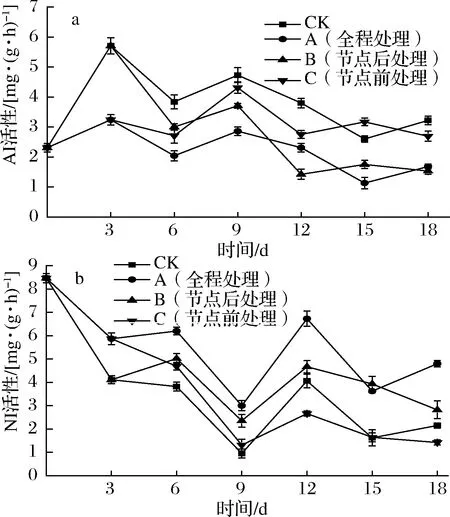
图9 不同外源乙烯处理时机对甘薯 AI(a)和NI(b)活性的影响Fig.9 Effect of different exogenous ethylene treatment on the activity of AI(a) and NI(b) activities in sweet potato
3 结论与讨论
实验结果表明节点后和全程1 g/L乙烯利处理能更好地抑制甘薯发芽。其中,CK组甘薯在第3天出现发芽迹象后,发芽一直处于旺盛状态;节点前乙烯处理虽能将甘薯的发芽时间延迟9 d,但开始发芽后芽苗迅速生长,储运后期发芽状态与CK组无明显差异,这可能是由于外源乙烯处理增大了甘薯中还原糖向蔗糖转化的速度[33],储备了更多蔗糖,并在甘薯开始发芽后为其提供能量;全程乙烯处理组在储运后期才出现发芽现象;节点后乙烯处理组虽然是在CK组出现发芽迹象后才进行乙烯处理,但与全程乙烯处理一样能起到有效的抑芽作用。
外源乙烯处理会促使部分还原糖转化为蔗糖,但节点前处理组在储运后期由于发芽旺盛会加速蔗糖消耗,节点后和全程处理组蔗糖含量在整个储运期间都维持在较高水平;同时节点后和全程乙烯处理还会明显增强甘薯的SPS活性,降低AI活性,而甘薯的SPS活性增强会加速蔗糖合成,AI的活性降低会减少蔗糖的不可逆分解[1],这些酶活性的改变均有利于甘薯中蔗糖的累积,而有研究表明,较高的蔗糖含量有助于抑制芽的生长[7-8]。
外源乙烯处理均会增强甘薯的呼吸强度,同时诱导α-淀粉酶、β-淀粉酶活性增大,从而加快甘薯淀粉向小分子可溶性糖转化的速率,导致甘薯淀粉含量快速下降的同时延缓可溶性糖含量下降的速率。但节点前处理组甘薯在取出乙烯利后呼吸强度迅速降低,之后由于发芽旺盛才再次回升,并且由于发芽旺盛需要消耗大量淀粉和可溶性糖,加速了可溶性糖含量的下降速率。对于食用甘薯来说可溶性糖含量高,甘薯甜度更高[27],因此节点后处理组和全程处理组甘薯的风味相对更佳。
综上所述,如果只是在节点前处理不能起到很好的抑芽作用,而传统的果蔬保鲜处理方式,大多都是在采收后、流通前进行处理,而这个时候甘薯还没有发芽,属于节点前。而在流通过程中节点后放入乙烯利不具备操作性。因此在流通前将乙烯利放在包装内,在整个流通过程中一直释放乙烯(即全程处理),抑芽作用效果最好,且具备操作性。节点后处理也有抑芽作用这一结论的另外一个应用意义在于,如果不是流通,而是在贮藏库贮藏过程中观察到甘薯出现萌芽迹象,在此时进行乙烯处理,可以抑制其他甘薯发芽以及芽生长,将经济损失降到最低。
