岩溶木本植物对干旱的生理生态适应
2021-01-12黄甫昭李健星李冬兴陈婷王斌陆树华李先琨
黄甫昭 李健星 李冬兴 陈婷 王斌 陆树华 李先琨
摘 要:受全球气候变暖和季风气候影响,西南岩溶区年降水量及其在季节间的分配发生明显变化,无雨期频率和持续时间增加,且基岩风化严重,基质储水能力差,致使巖溶木本植物面临的季节性和地质性干旱加剧。该文通过参考相关文献分析结构性状和生理调节探讨岩溶木本植物如何适应地质性和季节性干旱。结果表明岩溶木本植物应对干旱的策略与其他干旱、半干旱区的植物大体一致,主要有抗旱和避旱两种策略:抗旱性植物一般具有比叶面积小、叶肉多汁、储水组织发达、细胞液浓度高等适应干旱的特征,可通过增加木材密度、增强木质部导管的抗栓塞性和提高水分利用效率以适应干旱;避旱植物则可通过小而密的气孔和叶脉、发达的表皮毛、栅栏组织和维管束鞘等结构特征减少水分丧失,并可通过落叶、深根吸收深层水源和脱落酸(ABA)介导提早关闭气孔以适应干旱。虽然关于岩溶植物形态结构和生理调节对干旱适应机制的研究取得了一定进展,但仍然存在一些亟待解决的问题,例如:深入研究岩溶地区基岩水分状况及其对植物的贡献;加强岩溶木本植物根系结构和生物量分配、树木构型及根际微生物与木本植物干旱适应的协同关系研究;同时探索如何将岩溶植物生态适应研究成果应用于生产实践中,科学指导石漠化治理与生态修复。
关键词:岩溶,木本植物,干旱,形态结构,生理调节
中图分类号:Q948,Q945
文献标识码:A
文章编号:1000-3142(2021)10-1644-10
Abstract:Affected by global warming in southwest karst area and monsoon climate,the annual precipitation in southwest karst area and its distribution between seasons have changed obviously,and the frequency and duration of non-rain period are increasing. In addition,karst woody plants face more serious seasonal and geological droughts due to the strong weathering of bedrock and poor water storage capacity. This paper discusses how karst plants adapt to geological and seasonal drought by analyzing their structural characters and physiological regulation by referring to relevant literatures. The results indicate that like plants in arid and semi-arid areas,karst woody plants have two strategies to cope with drought:drought resistance and drought avoidance. Drought-resistant plants generally have some drought-resistant characteristics,such as small specific leaf area,succulent mesophyll,well-developed water storage tissue and high cell fluid concentration and so on,and can adapt to drought by increasing wood density,enhancing vessel anti-embolism and improving water use efficiency. Drought-avoidant plants can reduce water loss by small and dense stomata and leaf veins,developed superficial fur,palisade tissue and vascular sheath,and adapt to drought by defoliation,deep root absorption of deep water and early stomatal closure mediated by Abscisic acid (ABA). Although some progress has been made in the research on karst plants adapting to drought through morphological structure and physiological regulation,there are still some problems to be solved urgently,such as the study of bedrock water condition in karst area and its contribution to plants,the root structure and biomass allocation of karst woody plants,the relationship between tree configuration and drought adaptation and synergistic relationship between rhizosphere microorganisms and woody plants. At the same time explore how to apply the results of physiological and ecological adaptation of karst woody plants to drought in production practice,and scientifically guide the control and ecological restoration of rocky desertification.
Key words:karst,woody plants,drought,morphological structure,physiological regulation
水是植物赖以生存的必要条件,也是植物生命活动中最活跃的成分之一,对植物生长、发育和分布具有显著影响,尤其在干旱、半干旱地区和土壤浅薄的喀斯特地区,水是限制植物生长和分布的最主要因素(Gu et al.,2015; Sillmann et al.,2017; Fu et al.,2019; 谭凤森等,2019)。随着全球气温上升,许多地区的年降水量及其在季节间的分配发生明显改变,极端干旱发生的频率显著增加、持续时间更长,岩溶区季节性和地质性缺水有加剧趋势,引发大规模树木死亡的可能性更大,对现有岩溶植被的科学管护、可持续经营及石漠化区植被修复提出了挑战(Reichstein et al.,2013; Anderegg et al.,2019,2020; Brodribb et al.,2020)。
岩溶区是同沙漠边缘一样的脆弱生境,是世界上最主要的生态脆弱区之一。全球岩溶分布面积达2 200万km2,中国岩溶分布面积为344万km2,約占国土面积的1/3。我国是世界上岩溶分布面积最大的国家,以面积大、发育充分、形态多样、保存完好,成为“世界岩溶的立典之地”,其中,以西南岩溶区分布面积最大,主要分布在以云贵高原为中心的贵州、云南、广西和重庆等地(陈洪松等,2004;袁道先,2014)。
西南岩溶区虽然降水充沛,但因季节分配极为不均,时常出现季节性干旱,且其地表地下二元结构,地表水大量渗漏,地下水深埋,土层浅薄且分布不连续,土壤总量少,储水能力低,岩溶干旱严重,所以,水分亏缺是该地区植被生长分布及恢复重建的主要限制因子。经过长期进化和自然选择,岩溶木本植物形成复杂的结构和生理调节系统以适应其所处生境,理解岩溶木本植物干旱适应机制对植被恢复重建和现有植被科学管护具有重要意义。本文通过参考相关文献分析岩溶植物根系、木质部、叶片等功能性状和气孔开合、渗透势调节、抗氧化防御等生理调节探讨岩溶植物适应地质性和季节性干旱的机理,并总结分析亟待解决的问题及研究前景,为进一步研究岩溶植物的抗旱机理提供参考。
1 西南岩溶区季节性与地质性干旱特征
西南岩溶区受热带、亚热带季风气候影响,年降雨量可达1 100~2 000mm,但季节分配极为不均,春夏季多雨潮湿,秋冬季少雨干旱,加上全球气候变暖,年降水量及其在季节间的分配发生明显变化,极端干旱发生的频率显著增加,时常出现季节性干旱。
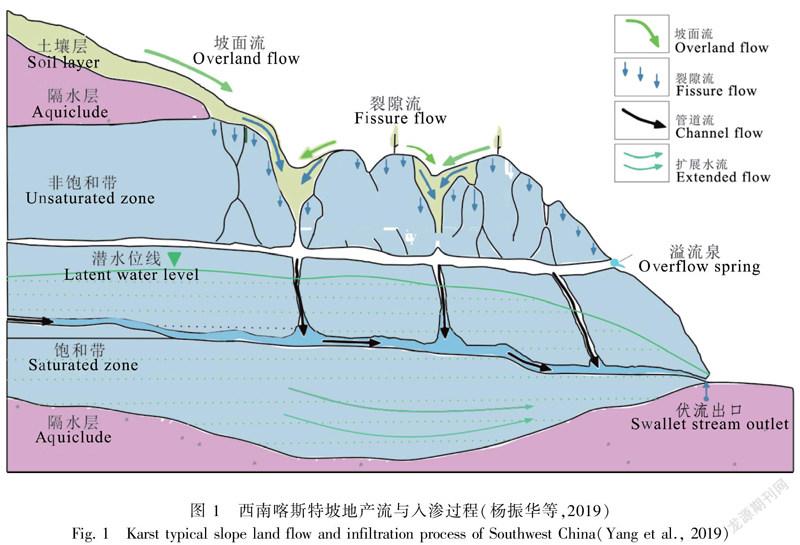
地质性干旱是岩溶区另一显著特征,是指在湿润气候条件下,水资源总量充沛的岩溶地区出现的一种特殊干旱现象,与碳酸盐岩岩性密切相关。碳酸盐岩风化形成的石灰土,其理化性质有别于地带性土壤,表现为富钙、偏碱、质地偏粘,营养元素供给速率慢且不平衡,土壤养分含量高但易流失。岩溶山区土壤与母岩之间界面明显,缺乏过渡层,结合力差,水土流失,且碳酸盐岩风化成土作用缓慢,致使这些地区土层非常瘠薄(Legrand et al.,1973;袁道先,2014)。西南岩溶地区年降水量在1 100 mm以上,叠加高温和碳酸盐岩的可溶性,地下河发育强烈,地表有较多溶洞、溶洼、溶沟、溶隙、漏斗和落水洞天窗等,水文地质结构表现为地表地下双重空间结构(图1)(杨振华等,2019)。另外,其土层浅薄,岩石裸露率高,致使雨水很快渗漏到地下,浅薄土层贮存的土壤水蒸发迅速,最终导致其在多雨的湿润季节,也频繁遭受地质性干旱。
2 岩溶植物结构对干旱的适应
作为一个有机体,植物会根据所处生境条件权衡各方面需求构建各器官结构以适应不断变化的环境,从而增加其存活率(Delzon,2015)。在季节性和地质性缺水易发的岩溶生态系统中,植物常通过调整根系、叶片和木质部等结构应对干旱,且各部位结构和功能往往存在着协同或权衡的关系。
2.1 植物根系结构对干旱的适应
岩溶区时常出现季节性和地质性干旱,但该区域依然能发育形成物种丰富、结构完整、层次复杂的植物群落的一个重要原因是不同树种能够占据不同的水文生态位(Schwinning,2013; Ding et al.,2018;黄甫昭等,2019)。根系是植物吸收水分和养分的重要器官,由于植物根系的趋水性,深根是植物在缺水生态系统或季节性干旱环境中的一个重要特征。在干旱地区,木本植物大多拥有垂直根系,其长可达地上部分的10倍。植物根系越发达,从深层土壤吸收的水分和养分越多,就越能适应干旱和贫瘠环境(任伟等,2020)。在喀斯特山地,可观察到很多植物的根系沿岩隙或湿润的岩壁下延达数十米深。对亚热带81个岩溶洞穴植物根系的DNA测序表明,根系样品分属12种植物,根系下扎深度大于10 m的树种有7个,有两种植物根系下扎深度达29 m(黄玉清等,2018)。当上层土壤水分有效性降低时,深根植物会增加对深层土壤/基岩水或地下水的利用,维持蒸腾和光合作用(West et al.,2012; Nie et al.,2019)。
植物水分来源与根系分布密切相关,有根系分布的基质层不一定是植物水分来源区,但无根系分布的基质层绝不能成为水分来源地。浅根系植物主要利用来自最近降水的浅层土壤水,而深根系植物则可同时利用不同深度的水源,且能依靠储存在岩石裂缝或裂隙中的水分抵御干旱(陈洪松等,2013)。研究南亚热带大块孤立出露基岩上同一群落内不同树种水分来源时发现,胸径较大的乔木菜豆树(Radermachera sinica)和圆叶乌桕(Triadicar otundifolia)等利用较深层的水源(基岩水或地下水),而胸径较小的灌木黄荆(Vitex negundo)、红背山麻杆(Alchornea trewioides)和石岩枫(Mallotus repandus)等则利用较浅的土壤/表层岩溶水源(Ding et al.,2021)。根据地上和地下生物量之间的异速生长关系,根系大小和深度通常随植株的生长而增加。岩溶生态系统中,根系扎入岩溶包气带获取基岩水或地下水是一个缓慢的过程,这个过程随植株大小和年龄的增加而增加(Schenk,2002; Phillips,2018),最终表现为大树根系多且深,可利用深层水源;小树根系浅而少,只能利用表层水源,通过不同的根系结构占据不同的水文生态位。
多項研究表明,深根植物能利用深层水分为其生长于干旱易发生态系统提供了潜在的竞争优势(Yang et al.,2017; McDowell et al.,2019)。Ding et al.(2021)研究亚热带岩溶次生群落发现,黄荆、红背山麻杆、火棘(Pyracantha fortuneana)和石岩枫等浅根植物在持续几个月的干旱中植株会落叶甚至死亡,而与其共存的深根植物菜豆树、圆叶乌桕和粗柄槭(Acer tonkinense)等却不会落叶和死亡。在意大利和美国得克萨斯州喀斯特地区进行的研究也表明浅根树种因干旱导致的树冠落叶和死亡率更高(Johnson et al.,2018;Crouchet et al.,2019)。可见,在时常面临季节性干旱的生态系统中,根系深扎是植物应对干旱的一个关键的功能特征。与深根系植物通过利用岩石裂隙水或深层水分抗旱不同,浅根系植物一般可通过提高水分利用效率和发展高密度浅层根系提高对浅层水分的吸收效率,还可通过降低叶片气孔导度和提前落叶以适应干旱(Schwinning,2010;陈洪松等,2013;Liu et al.,2021)。
此外,还有研究表明,植物蒸腾和地面蒸发减弱或停止的旱季,深根植物可从深层湿润土层中吸收水分,再由侧根释放到表层较干燥的土层中;当上层土壤湿润而底层土壤干燥时,水分会沿着根系由上层土壤向深层流动(Dawson,1993; Fan et al.,2013; Sun et al.,2018)。可见同一群落中,植物根系结构和分布差异对水分吸收和再分配具有重要作用,且存在一种非常契合的互利共存机制,从而维持自然生态系统中物种多样性和群落稳定性。
2.2 木质部结构对干旱的适应
植物木质部主要由导管、薄壁组织和纤维组成,导管可纵向输送水分和营养物质,薄壁组织主要有储存水分、非结构性碳水化合物和其他营养素以及防御病原体的作用,而纤维具有机械支撑作用(Plavcová et al.,2016; Schenk et al.,2017)。木质部性状是植物水分运输、物质存储、机械支持的关键功能性状。岩溶常绿植物通常采取保守的“高安全性”水分利用策略,具有较大的木材密度和较小的导管,水分传导速率和光合作用速率相对落叶树种较低,但有更强的维持膨压、抵抗气穴化栓塞能力和更高的水分利用效率,能够在缺水环境下维持水分运输,从而维持叶片和枝条的生理功能(Maherali et al.,2004;谭凤森等,2019;Tan et al.,2020;Ding et al.,2021)。岩溶落叶植物则倾向于采用冒险的“高效率”水分利用策略,具有小的木材密度和较大的木质部导管,因而具有较高的导水率和光合作用能力,通过雨季快速生长补偿旱季碳物质的损失。落叶植物的叶片膨压丧失点较高,对干旱更为敏感,当面临一定的水分胁迫时,木质部更易发生气穴化栓塞,气孔关闭也较早;当干旱进一步加重时,则通过落叶降低蒸腾以减少水分的丧失,从而维持水分输送安全(Choat et al.,2005; Johnson et al.,2012; Zhu et al.,2017)。半落叶植物的水分利用策略介于常绿和落叶植物之间。与落叶物种相比,半落叶植物木质部导管较小,对栓塞的抵抗力相对更强,但由于干旱期间,部分叶片继续进行光合碳合成,蒸腾耗水使植物体内水势更负,更易导致不可逆的木质部栓塞,最终导致植株枯死。与常绿植物相比,半落叶植物的木质部导管更大,极端干旱条件下更容易栓塞。因此,半落叶植物的干旱耐受策略在中度干旱时会显示兼顾水力安全和碳合成的优势,但在极端干旱时,其较长的生长期增加了水分输送风险(Kukowski et al.,2013)。Chen et al.(2021)在研究云南元江干热河谷稀树草原植被时观察到同一干旱事件中半落叶树种的枝条死亡率和枯梢率更高,也证实了干旱胁迫时半落叶植物水力安全风险更高。
同时,有些岩溶植物如薄叶羊蹄甲(Bauhinia tenuiflora)和囊托羊蹄甲(B. touranensis)的木质部导管具有独特的“二元性”,即同时具有粗大的导管(>400 μm)和众多细小导管(<30 μm),可以兼顾“高安全性”和“高效率”(Zhu et al.,2017)。另外,相比同区域的非岩溶森林,很多岩溶优势植物的木质部具有丰富的薄壁细胞,具有储存水分和碳水化合物的功能,在防止栓塞扩散以及栓塞修复方面具有重要作用(Morris et al.,2016)。在干旱条件下,薄壁组织较多的树种更多地依赖木质部薄壁组织促进栓塞体修复,而不是完全依靠其较强的抗栓塞性,植物木质部存在储藏能力和抗栓塞性的权衡(Chen et al.,2014; Pratt et al.,2017; Kiorapostolou et al.,2019)。
由此可见,岩溶植物木质部结构分化在适应干旱中具有重要作用。岩溶常绿植物通常拥有较大的木材密度和较小的导管,增强木质部抗栓塞能力,采用“高安全性”水分利用策略,落叶物种则倾向采用“高效率”水分利用策略,木质藤本植物则采用“高安全性”和“高效率”兼顾的水分利用策略。
2.3 叶片结构对干旱的适应
叶片是植物较敏感和脆弱的部分,为了适应干旱的环境,旱生植物的叶子通常朝着降低蒸腾和增加贮藏水分两个方面发展。岩溶植物为适应所处生境,叶片常常厚而小,同时具有较厚的蜡质层和角质层,栅栏组织发达,排列紧密,比叶面积较小(Cao,2014; Geekiyanage et al.,2018);同时叶肉多汁,储水组织发达,细胞液浓度高,保水力强(Wright et al.,2004);此外,岩溶植物叶片、枝条上常覆盖有较厚的柔毛,以减少蒸腾失水,而且叶片被毛还可以增加对小降雨量和冷凝水的利用(López et al.,2016;谭凤森等,2019)。Fu et al.(2019)在比较中国西南岩溶和非岩溶植物叶片功能性状时发现,岩溶植物的叶片氮磷含量、近轴表皮厚度、远轴表皮厚度、气孔密度及叶片水分利用效率显著高于邻近区域的非岩溶植物,在水分利用结构和生理上更为保守。同时岩溶植物的叶脉密度大、导管长、抗栓塞性强,能快速地将水分输送至叶片的各个部位,保证水分供应安全性,提高光合速率(Chen et al.,2021)。由此可见,叶片结构在岩溶植物适应季节性缺水中具有重要作用,在长期的季节性缺水生境下,岩溶植物叶片往往利用小而密的气孔和叶脉、发达的表皮毛、栅栏组织和维管束鞘等结构特征来减少水分丧失,从而适应季节性和地质性干旱。
3 巖溶植物适应干旱的生理调节
植物为适应干旱,除了形成独特的形态结构以外,还可通过一系列的生理调控来适应,如气孔调节、渗透势调节、抗氧化防御和内源激素调节等。
3.1 气孔调节与干旱适应
气孔调节是植物适应所处生境水分条件的重要途径之一。叶片膨压丧失点水势与气孔关闭时的水势密切相关,是植物水力学中非常关键的性状,与叶片的渗透调节能力和耐旱性有紧密联系(Bartlett et al.,2014; Maréchaux et al.,2015)。研究发现植物在失水过程中,叶片细胞膨压的丧失会引起气孔关闭,而气孔在木质部水势降低至发生气穴化阈值之前便会关闭,从而减缓植物木质部水势的下降速率,避免木质部气穴化栓塞(Nicolas et al.,2017; Wang et al.,2017; 杭红涛等,2019)。北热带岩溶木本植物的叶片膨压丧失点较低,大部分物种并不会通过提前关闭气孔以防止水势的降低,同时还保持较高光合速率,说明岩溶木本植物还具有其他有效的水力安全策略(Fu et al.,2012;谭凤森等,2019)。Zhang et al.(2021)研究发现,中国西南岩溶森林绝大多数树种枝条的P50(丧失50%导水率时的木质部水势)要小于树叶P50,且树叶的木质部导管先于树枝栓塞,可减少水分继续损耗,避免枝条木质部栓塞受损。在干旱生境中保护树枝比保护树叶更加重要(Zhu et al.,2016),通过叶片栓塞保护枝条水分运输安全,具有“舍车保帅”的作用,这可能是岩溶植物没有过早关闭气孔又能应对干旱的调节机制之一。
3.2 渗透势调节与干旱适应
渗透势调节是植物应对干旱的另一有效途径。在干旱生境下,为防止过度失水,植物会积累大量的无机和有机渗透调节剂来降低细胞的渗透势,增强细胞持水能力,改善植株水分状况,从而提高植物的适应性(Lum et al.,2014;代英超等,2015;Nohong et al.,2015)。岩溶植物叶片可溶性糖等渗透调节剂的积累在叶片膨压维持中具有重要作用。对亚热带岩溶常见植物青冈(Cyclobalanopsis glauca)、圆叶乌桕、火棘和小果蔷薇(Rosa cymosa)的研究发现,随着干旱加剧,这些植物通过积累脯氨酸、可溶性糖等有机渗透调节剂调控质膜渗透势,维护酶和细胞膜稳定,减少水分胁迫对细胞的伤害;同时累积钠、钾等无机离子调节液泡渗透势,维持细胞膨压(Liu et al.,2011;张中峰等,2012;刘珊等,2016)。Geekiyanage et al.(2018)在研究喀斯特季节性雨林不同生境优势树种叶片功能性状时发现,干旱山顶优势树种叶片积累的可溶性糖含量是低海拔湿润生境优势树种的四倍。欧芷阳等(2018)对北热带喀斯特季节性雨林建群种蚬木(Excentrodendron tonkinense)的研究发现,蚬木在重度干旱胁迫下通过累积丙二醛和脯氨酸以适应干旱胁迫。
由此可见,地质性或季节性缺水时,岩溶植物可通过在体内积累各种渗透调节物质以调控质膜渗透势,维护酶和细胞膜的稳定,减少干旱对细胞的伤害,从而维持植物个体的存活和生长。
3.3 抗氧化防御与干旱适应
抗氧化防御也是植物适应干旱的另一重要途径。植物有氧代谢过程会产生活性氧,正常条件下,活性氧可以维持动态平衡状态,但干旱胁迫会扰乱细胞的自我调节,导致活性氧在细胞中过量累积,破坏膜质、叶绿体结构,损伤蛋白质、核酸等生物大分子,引起植物代谢紊乱,最终导致植株死亡(Lü et al.,2019)。经过长期进化,植物细胞形成了一套复杂的抗氧化防御系统以维持细胞内活性氧的稳态平衡。抗氧化物质主要包括抗氧化酶和抗氧化剂两类,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)等是植物细胞的主要抗氧化酶(Gill et al.,2010;任伟等,2020)。植物体内抗氧化酶及其活性越高,其抗旱能力就越强(Frazeli et al.,2007;刘锦春等,2011)。岩溶区不同演替阶段代表性植物蚬木、构树(Broussonetia papyrifera)和火棘等在适度干旱条件下依靠增加体内SOD、POD、CAT的含量或活性应对干旱(井瑾等,2017;欧芷阳等,2018)。干旱条件下,植物膜脂过氧化水平和丙二醛含量升高,会影响到细胞的正常代谢,岩溶植物可通过增强保护酶活性等抗氧化防御以清除活性氧自由基,降低膜脂过氧化程度,避免干旱对其造成伤害,从而增强其抗旱性。
3.4 碳酸酐酶活性与干旱适应
碳酸酐酶(carbonic anhydrase,CA)广泛存在于植物及单细胞绿藻中,可催化CO2+H2OHCO3-+H+双向反应,加速无机碳向羧化酶活性部位扩散,提高CO2固定效率(李强等,2011)。植物叶片中碳酸酐酶(CA)除了能增强CO2固定能力外,其活性对植物适应干旱亦有积极意义(罗绪强等,2012)。为应对岩溶生境的季节性和地质性干旱,岩溶植物通常通过关闭气孔或者减小气孔导度来应对,这一生理行为在减少蒸腾防止叶片水分进一步丧失的同时也会减少叶片对大气CO2的吸收,影响植物的CO2同化能力。为应对干旱引起的碳源减少,岩溶植物可通过碳酸酐酶高效利用碳酸氢根离子作为碳源。在遭受干旱时,岩溶植物的叶片碳酸酐酶活力升高,通过将细胞内的碳酸氢根离子转化成水和CO2,应对干旱引起的CO2不足(吴沿友,2011)。岩溶植物通过碳酸酐酶利用岩溶环境中丰富的碳酸氢根离子合成水和CO2以补充干旱引起的叶片水分不足和胞间CO2减少,在提高植物水力安全的同时维持光合产物的合成,提高岩溶植物的干旱适应性,这是岩溶植物最为特别的干旱适应策略。
3.5 脱落酸(ABA)调节与干旱适应
在水分缺乏条件下,气孔开合、渗透势调节、抗氧化防御和碳酸酐酶活性是植物应对干旱的主要生理途径,它们通常作为一个有机系统协同应对干旱,而这种协同主要由脱落酸(ABA)介导。ABA是植物体内一种重要的植物激素,可以调节逆境反应、代谢变化、基因表达、种子休眠、幼苗生长和植物蒸腾等(Fujii et al.,2009;Liu et al.,2021)。干旱会诱导大量ABA产生,一方面,产生的ABA可使保卫细胞胞质Ca2+浓度升高,抑制质膜上的H+泵,使质膜上的H+-ATP酶(质膜上的H+-ATP酶是H+和K+的转运体)活性降低,从而抑制H+外流和K+内流并关闭叶片气孔,进而降低植物的光合作用和蒸腾能力,最终保证植物的水力安全(Sukhova et al.,2017)。另一方面,ABA产生后,保卫细胞胞浆pH升高,K+外流的通道被激活,保卫细胞中的K+流出,导致细胞脱水和气孔关闭(Kim et al.,2010;Liu et al.,2021)。可见,植物激素,尤其是ABA通过复杂的调节机制在诱发植物一系列干旱适应机制中具有不可替代的作用。
4 总结与展望
植物作为生态系统的初级生产者,其生产力关乎整个生态系统的物质和能量供给。在当前全球气候变暖、极端天气增加、高温干旱频发的形势下,水资源短缺仍是未来全球面临的主要问题,因此,有关植物适应干旱的研究将会持续成为植物科学的前沿研究领域之一。
近年来,许多学者从形态结构和生理调节等方面对岩溶木本植物干旱适应性进行了研究。结果表明,岩溶木本植物应对干旱主要有抗旱和避旱两种策略:抗旱性植物一般具有比叶面积小、叶肉多汁、储水组织发达、细胞液浓度高等适应干旱的特征,同时通过增加木材密度,增强木质部导管抗栓塞性和提高水分利用效率以适应干旱;避旱植物则通过发育小而密的气孔和叶脉、发达的表皮毛、栅栏组织和维管束鞘等结构特征以减少水分丧失,并通过落叶、深根吸收深层水源和ABA介导提早关闭气孔以适应干旱。虽然对岩溶木本植物适应干旱的认识不断深入,但由于地质背景的特殊性、地形地貌的复杂性和生境的高度异质性,岩溶山区水分运移和赋存过程十分复杂,加上技术手段的限制,当前研究还存在一些问题,例如:难以明确岩溶地区基岩水分状况及其对植物的贡献、岩溶异质性生境不同植物的干旱适应策略、岩溶木本植物根系结构及其对干旱适应的意义以及根际微生物在岩溶植物水分获取中的作用等。因此,提出以下建议:
(1)深入研究岩溶地区基岩水分状况及其对植物的贡献。一直以来,对岩溶地区水分状况的认识是岩溶基岩缝隙管道丰富,降水迅速从地下管道流失然后补给地下水,所以,岩溶区的主要水源是土壤水和地下水。虽然有研究表明植物可利用岩石缝隙和基岩水分(Rose et al.,2003;陈洪松等,2013;Rempe et al.,2018; Ding et al.,2021),但其并不被认为是一个重要的水库。最近,McCormick et al.(2021)对植物水分来源研究表明,木本植物通常会获取大量储存在基岩中的水分用于蒸腾作用,美国加州地区植物对基岩水的利用超过年均降水量的25%以上,利用基岩水补充蒸腾的植物的生物量占整个区域地上生物量的50%以上。但截止目前,岩溶区基岩水分状况及其对植物的贡献所知甚少,岩溶地区植物是否受到地质性干旱限制也存在争议。因此,需要加强对岩溶地区基岩水的评估,可进一步理解岩溶植物的水分来源及其适应策略,同时对岩溶地区水资源管理和开发具有重要意义。
(2)加强岩溶木本植物根系结构和生物量分配研究。光合产物分配是植物适应各种环境的主要策略之一,根冠比是植物适应干旱的重要评估指标之一(Joslin et al.,2000; Foxx et al.,2019)。水分不足和养分缺乏会促使光合产物向根生长的相对分配,最终导致植物具有更高的根冠比和更强的水分和矿物质吸收能力(Kozlowski,2002; Seleiman et al.,2021)。岩溶地区水分、养分相对缺乏,植物是否将更多的光合产物用于根系生长,是理解岩溶植物干旱适应的关键科学问题之一。但岩溶植物根系大部分穿插于岩石缝隙,传统的开挖土壤剖面和微根管观测法很难在岩溶地区使用,限制了对岩溶地区植物根系结构和地下生物量的研究。今后可通过岩溶生境模拟装置开展相关控制实验,加强岩溶木本植物的光合产物分配策略研究,并探索通过穿透射线成像(肖爽等,2020;Liang et al.,2020)等原位根系观测识别技术对岩溶植物根系结构及其生物量进行研究,不但能深入理解岩溶植物如何通过调整根系结构来适应干旱,而且对准确评估岩溶植被碳汇功能具有重要意义。
(3)开展岩溶木本植物树木构型与干旱适应关系研究。现有研究从根系、树干和叶片结构等方面探讨了岩溶木本植物对干旱的适应机制,而树木构型作为植物最直观的结构却少有人研究其在植物水分获取和干旱适应方面的功能。半干旱区研究表明植物为应对干旱,通常形成“大冠幅,小树高”的树木構型(Dai et al.,2020),半干旱区灌木通过冠层汇集形成的树干茎流聚集到根部土壤的水分是当地降雨量的11~221倍,植物在干旱季可使用这些聚集的水分,表明冠层汇集聚水是灌木对干旱环境的一种主动适应形式和高效用水策略(李小雁,2011)。在岩溶峰丛区,时常能观察到从洼地到山脊(海拔高差100~300 m),植被迅速从高大乔木变成矮小灌木,是为由岩溶木本植物应对干旱的策略调控值得探究。
(4)开展岩溶环境下根际微生物与木本植物协同关系研究。大量研究表明,在土壤-植物根系-微生物互作体系中,根系中的细菌能通过产生植物生长调节因子或调节植物的激素水平促进植物根系发育及伸长,促进植物产生抗氧化物和活性氧清除剂等,从而提升植物的耐旱水平(Lakshmanan et al.,2017; 庞志强等,2020);菌根真菌则通过其菌丝网络帮助宿主植物吸收水分、养分,还能调控宿主植物的水通道蛋白、干旱相关基因,促进植物在干旱胁迫下的代谢和生长(Miransari,2014; 张中峰等,2018; 袁志林等2019)。岩溶生境干旱频发,养分相对缺乏,植物与微生物如何协同应对生境变化,可能是理解植物适应干旱的有效途径之一。
(5)科学研究的最终目是指导生产实践,如何把岩溶植物对干旱的适应研究成果应用于生产,也是值得重视的问题。例如:基于岩溶木本植物干旱适应性状构建抗旱植物筛选模型,为岩溶抗旱植物筛选提供支撑;筛选能提高岩溶植物水分吸收能力的微生物,研发菌根化育苗技术,增强种苗适应岩溶干旱的能力、提高造林成活率;科学评估石漠化山地植被修复的生态需水量;基于岩溶木本植物水文生态位分化研究,为植被恢复和复合农林经营的物种配置提供理论依据。
中国西南岩溶区已知分布有维管束植物7 000余种,植物资源丰富且特有性强,岩溶生境多样化孕育了植物多样化的表型,为研究木本植物适应特殊生境的结构、生理及其遗传学基础提供了丰富的资源。因此,随着技术手段的不断发展、岩溶生态系统研究方法论的逐渐完善,植物对岩溶地质性和季节性干旱的适应性研究将取得大量创新性的成果,也将有利于岩溶区的生态修复与生态系统管理。
致谢 感谢多位匿名审稿人对本文撰写提出的宝贵意见和建议。
参考文献:
ANDEREGG WRL,TRUGMAN AT,BADGLEY G,et al.,2020. Divergent forest sensitivity to repeated extreme droughts\[J]. Nat Clim Change,10(12):1-5.
BARTLETT,MK,ZHANG Y,KREIDLER N,et al.,2014. Global analysis of plasticity in turgor loss point,a key drought tolerance trait [J]. Ecol Lett,17(12):1580-1590.
BRODRIBB TJ,POWERS J,COCHARD H,et al.,2020. Hanging by a thread? Forests and drought\[J]. Science,268(6488):261-266.
CAO KF,FU PL,CHEN YJ,et al.,2014. Implications of the ecophysiological adaptation of plants on tropical karst habitats for the ecological restoration of desertified rocky lands in southern China\[J]. Sci Sin Vit,2014,44(3):238-247.[曹坤芳,付培立,陈亚军,等,2014. 热带岩溶植物生理生态适应性对于南方石漠化土地生态重建的启示\[J]. 中国科学:生命科学,44(3):238-247]
CHEN HS,NIE YP,WNAG KL,2013. Spatio-temporal heterogeneity of water and plant adaptation mechanisms in karst regions:a review [J]. Acta Ecol Sin,33(2):317-326. [陈洪松,聂云鹏,王克林,2013. 岩溶山区水分时空异质性及植物适应机理研究进展 [J]. 生态学报,33(2):317-326.]
CHEN HS,WANG KL,2004. Characteristics of karst drought and its countermeasures [J]. Res Agric Mod,25:70-73. [陈洪松,王克林,2004. 岩溶干旱特征及其治理对策 [J]. 农业现代化研究,25:70-73.]
CHEN YJ,CAO KF,SCHNITZER SA,et al.,2014. Water-use advantage for lianas over trees in tropical seasonal forests [J]. New Phytol,205:128-136.
CHEN YJ,CHOAT B,STERCK F,et al.,2021. Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form [J]. Ecol Lett. DOI:10.1111/ele.13856.
CHEN ZC,ZHANG YT,YUAN WJ,et al.,2021. Coordinated variation in stem and leaf functional traits of temperate broadleaf tree species in the isohydric-anisohydric spectrum [J]. Tree Physiol,DOI:10.1093/treephys/tpab028.
CHOAT B,BALL MC,LULY JG,et al.,2005. Hydraulic architecture of deciduous and evergreen dry rainforest tree species from north-eastern Australia [J]. Trees,19(3):305-311.
CROUCHET SE,JENSEN J,SCHWARTZ BF,et al.,2019. Tree mortality after a hot drought:distinguishing density-dependent and-independent drivers and why it matters [J]. Front For Glob Chang,2:21.
DAI JY,LIU HY,WANG YC,et al.,2020. Drought-modulated allometric patterns of trees in semi-arid forests [J]. CommBiol,405(2020):1234567890.
DAI YC,XU KY,MA K,et al.,2015. Physiological responses of the rare and endangered Ardisia violacea (Myrsinaceae) seedlings to progressive drought stress [J]. Acta Ecol Sin,35(9):2954-2959. [代英超,徐奎源,馬凯,等,2015珍稀濒危植物堇叶紫金牛对持续干旱的生理响应 [J]. 生态学报,35(9):2954-2959.]
DAWSON TE,1993. Hydraulic lift and water use by plants:implications for water balance,performance and plant-plant interactions [J]. Oecologia,95 (4):565-574.
DELZON S,2015. New insight into leaf drought tolerance [J]. Funct Ecol,29(10):1247-1249.
DING YL,NIE YP,CHEN HS,et al.,2021. Water uptake depth is coordinated with leaf water potential,water-use efficiency and drought vulnerability in karst vegetation [J]. New Phytol,229(3):1339-1353.
DING YL,NIE YP,SCHWINNING S,et al.,2018. A novel approach for estimating groundwater use by plants in rock-dominated habitats [J]. J Hydrol,565:760-769.
FAN T,LI J,LI Y,et al. 2013. Stable isotopic study on water utilization sources of Pinus yunnanensis plantations in the central Yunnan karst plateau [J]. Adv Mat Res,726-731:3888-3893.
FRAZELI F,GHORBANLI M,NIKNAM V,2007. Effect of drought on biomass,protein content,lipid zeroxidation and antioxidant enzymes in two sesame cultivars [J]. Biol Plant,51(1):98-103.
FUJII H,ZHU JK,2009. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth,reproduction,and stress [J]. Proc Natl Acad Sci,106(20):8380-8385.
FU PL,JIANG YJ,WANG AY,et al.,2012. Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest [J]. Ann Bot,110(1):189-199.
FU PL,ZHU SD,ZHANG JL,et al.,2019. The contrasting leaf functional traits between a karst forest and a nearby non-karst forest in south-west China [J]. Funct Plant Biol,46(10):907-915.
FOXX AJ,FORT F,2019. Root and shoot competition lead to contrasting competitive outcomes under water stress:A systematic review and meta-analysis [J]. PLoS ONE,14(12):e0220674.
GEEKIYANAGE,N,GOODALE,UM,CAO,KF,et al.,2018. Leaf trait variations associated with habitat affinity of tropical karst tree species [J]. EcolEvol,8(1):286-295.
GILL S,TUTEJA N,2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants [J]. Plant Physiol Biochem,(48):909-930.
GU DX,ZHANG ZF,MALLIK A,et al.,2015. Seasonal water use strategy of Cyclobalanopsis glauca in a karst area of southern China [J]. Environ Earth Sci,74(2):1007-1014.
HANG HT,WU YY,ZHANG KY,et al.,2019. Effects of simulated karst soil habitat stresses on photosynthetic characteristics and dry matter allocation of Robinia pseudoacacia seedlings [J]. Chin J Ecol,38(9):2648-2654. [杭紅涛,吴沿友,张开艳,等,2019. 模拟喀斯特不同土壤生境胁迫对刺槐幼苗光合特性及干物质分配的影响 [J]. 生态学杂志,38(9):2648-2654.]
HUANG FZ,LI DX,WANG B,et al.,2019. Foliar stable carbon isotope composition and water use efficiency of plant in the Karst seasonal rain forest [J]. Chin J Appl Ecol,30 (6):1833-1839. [黄甫昭,李冬兴,王斌,等,2019. 喀斯特季节性雨林植物叶片碳同位素组成及水分利用效率 [J]. 应用生态学报,30(6):1833-1839.]
HUANG YQ,ZHANG ZF,MO L,et al.,2018. Study on the distribution of plant deep root system and its eco-geological function in karst area of south China [R] [黄玉清,张中峰,莫凌,等,2018. 我国南方岩溶区植物深根系分布及其生态地质作用研究 [R]. 科技成果.]
JING J,WANG FZ,ZHANG Y,et al.,2017. Seed germination of Pyracantha fortuneana and the change of antioxidant enzyme activity under drought stress [J]. J Arid Land Resourc Environ,(9):135-139. [井瑾,王方贞,张瑜,等,2017. 干旱胁迫下火棘种子萌发过程及其抗氧化酶活性变化 [J]. 干旱区资源与环境,(9):135-139.]
JOHNSON DM,DOMEC JC,BERRY ZC,et al.,2018. Cooccurring woody species have diverse hydraulic strategies and mortality rates during an extreme drought [J]. Plant Cell Environ,41:576-588.
JOHNSON DM,MCCULLOH KA,WOODRUFF DR,et al.,2012. Hydraulic safety margins and embolism reversal in stems and leaves:Why are conifers and angiosperms so different? [J]. Plant Sci,195(none):48-53.
JOSLIN JD,WOLFE MH,HANSON PJ,2000. Effects of altered water regimes on forest root systems [J]. New Phytol,147(1):117-129
KIM T,BHMER M,HU H,et al.,2010. Guard cell signal transduction network:advances in understanding abscisic acid,CO2,and Ca2+signaling [J]. Ann Rev Plant Biol,61:561-591.
KIORAPOSTOLOU N,DA SOIS L,PETRUZZELLIS F,et al.,2019. Vulnerability to xylem embolism correlates to wood parenchyma fraction in angiosperms but not in gymnosperms [J]. Tree Physiol,39(10):1675-1684.
KOZLOWSKI TT,PALLARDY SG,2002. Acclimation and adaptive responses of woody plants to environmental stresses [J]. Bot Rev,68(2):270-334.
KUKOWSKI KR,SCHWINNING S,SCHWARTZ BF,2013. Hydraulic responses to extreme drought conditions in three co-dominant tree species in shallow soil over bedrock [J]. Oecologia,171(4):819-830.
LAKSHMANAN V,RAY P,CRAVEN KD,2017. Toward a resilient,functional microbiome:drought tolerance-alleviating microbes for sustainable agriculture [J]. Methods Mol Biol,1631:69-84.
LEGRAND HE,STRINGFIELD VT,1973. Karst hydrology—A review [J]. J Hydrol,20(2):97-120.
LI Q,HE YY,CAO JH,et al.,2011. The plant carbonic anhydrase at karst area and its ecological effects [J].Ecol Environ Sci,20(12):1867-1871. [李強,何媛媛,曹建华,等,2011. 植物碳酸酐酶对岩溶作用的影响及其生态效应 [J]. 生态环境学报,20(12):1867-1871.]
LIANG H,XING LY,LIN JH,2020. Application and algorithm of ground-penetrating radar for plant root detection:a review [J]. Sensors (Basel,Switzerland),20(10):2836.
LIU CC,LIU YG,GUO K,et al. 2011. Effect of drought on pigments,osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China [J]. Environ Exp Bot,71(2):174-183.
LIU CN,HUANG Y,WU F,et al.,2021. Plant adaptability in karst regions [J]. J Plant Res,134(5):1-18.
LIU JC,ZHONG ZC,HE YJ,2011. Effects of drought stress and re-watering on the active oxygen scavenging system of Cupressus funebris seedlings in Karst area [J]. Chin J Appl Ecol,22 (11):2836-2840. [刘锦春,钟章成,何跃军,2011. 干旱胁迫及复水对喀斯特地区柏木幼苗活性氧清除系统的影响 [J]. 应用生态学报,22 (11):2836-2840.]
LIU WN,CHEN HS,ZOU QY,et al. 2021. Divergent root water uptake depth and coordinated hydraulic traits among typical karst plantations of subtropical China:Implication for plant water adaptation under precipitation changes [J].Agric Water Manag,249:1-10.
LIU S,HE Q,LI JY,et al.,2016. Physiological responses of the limestone endemic plant Triadica rotundifolia seedlings to drought stress [J]. J S Chin Agric Univ,37(2):96-100. [劉珊,何茜,李吉跃,等,2016. 石漠化树种圆叶乌桕对干旱胁迫的生理响应 [J]. 华南农业大学学报,37(2):96-100.]
LI XY,2011. Mechanism of coupling,response and adaptation between soil,vegetation and hydrology in arid and semiarid regions [J]. Sci Sin Terrae,41(12):1721-1730. [李小雁,2011. 干旱地区土壤-植被-水文耦合、响应与适应机制 [J]. 中国科学:地球科学,41(12):1721-1730.]
LPEZ R,CANO FJ,CHOAT B,et al.,2016. Plasticity in vulnerability to cavitation of Pinus canariensis occurs only at the driest end of an aridity gradient [J]. Front Plant Sci,7:769.
L XP,GAO HJ,ZHANG L,et al.,2019. Dynamic responses of Haloxylon ammodendron to various degrees of simulated drought stress [J]. Plant Physiol Biochem,(139):121- 131.
LUM S,HANAFI MM,RAFII YM,et al.,2014. Effect of drought stress on growth,proline and antioxidant enzyme activities of upland rice [J]. J Anim Plant Sci,24(5) :1487-1493.
LUO XQ,WANG CY,YANG HY,et al.,2012. Studies on adaptive mechanisms of karst dominant plant species to drought and high calcium stress\[J] Chin Agric Sci Bull,28(16):1-5.[罗绪强,王程媛,杨鸿雁,等,2012. 喀斯特优势植物种干旱和高钙适应性机制研究进展\[J]. 中国农学通报,28(16):1-5.]
MAHERALI H,POCKMAN WT,JACKSON RB,2004. Adaptive variation in the vulnerability of woody plants to xylem cavitation [J]. Ecology,85:2184-2199.
MARCHAUX I,BARTLETT K,SACK L,et al.,2015. Drought tolerance as predicted by leaf water potential at turgor loss point varies strongly across species within an Amazonian forest [J]. Funct Ecol,29(10):1268-1277.
MCCORMICK EL,DRALLE DN,HAHM WJ,et al.,2021. Widespread woody plant use of water stored in bedrock [J]. Nature,597(7875):225-229.
MCDOWELL NG,GROSSIORD C,ADAMS HD,et al.,2019. Mechanisms of a coniferous woodland persistence under drought and heat [J]. Environ Res Lett,14:045014.
MIRANSARI M,2014. Mycorrhizal fungi to alleviate compaction stress on plant growth [M]//MIRANSARI M. Use of microbes for the alleviation of soil stresses. New York:Springer,1:165-174.
MORRIS H,PLAVCOV L,CVECKO P,et al.,2016. A global analysis of parenchyma tissue fractions in secondary xylem of seed plants [J]. New Phytol,209(4):1553-1565.
NICOLAS MS,DELZON S,COCHARD H,2017. Plant resistance to drought depends on timely stomatal closure [J]. Ecol Lett,20(11),Doi:10.1111/ele.12851.
NIE YP,DING YL,ZHANG H,et al.,2019. Comparison of woody species composition between rocky outcrops and nearby matrix vegetation on degraded karst hillslopes of Southwest China [J]. J For Res,30:911-920.
NOHONG B,2015. Effect of water stress on growth,yield,proline and soluble sugars contents of signal grass and napier grass species [J]. Am-Eur J Sustain Agric,9:14-21.
OU YZ,CAO YY,TAN CQ,et al.,2018. Effects of drought on photosynthesis and resistance physiology of Excentrodendron hsienmu seedlings in karst habitat [J]. Chin J Ecol,37(11):3270-3276. [歐芷阳,曹艳云,谭长强,等,2018. 干旱胁迫对喀斯特生境蚬木幼苗光合特性及抗性生理的影响 [J]. 生态学杂志,37(11):3270-3276.]
PANG ZQ,YU DQ,2020. Plant root system-microbial interaction system under drought stress and its application [J]. Plant Physiol J,56 (2):109-126. [庞志强,余迪求,2020. 干旱胁迫下的植物根系-微生物互作体系及其应用 [J]. 植物生理学报,56(2):109-126.]
PHILLIPS JD,2018. Self-limited biogeomorphic ecosystem engineering in epikarstsoils [J]. Phys Geogr,39:304-328.
PLAVCOV L,HOCH G,MORRIS H,et al.,2016. The amount of parenchyma and living fibers affects storage of nonstructural carbohydrates in young stems and roots of temperate trees [J]. Amer J Bot,103(4):603-612.
PRATT RB,JACOBSEN AL,2017. Conflicting demands on angiosperm xylem:tradeoffs among storage,transport and biomechanics [J]. Plant Cell Environ,40(6):897-913.
REN W,GAO HJ,WANG RJ,et al.,2020. Research advances in adaptation of higher plants to arid habitats [J]. J Grassl Forage Sci,254(3):8-19. [任伟,高慧娟,王润娟,等,2020. 高等植物适应干旱生境研究进展 [J]. 草学,254(3):8-19.]
REMPE DM,DIETRICH WE. 2018. Direct observations of rock moisture,a hidden component of the hydrologic cycle [J]. Proc Natl Acad Sci,115:2664-2669.
ROSE K,GRAHAM R,PARKER D,2003. Water source utilization by Pinus jeffreyi and Arctostaphylos patula on thin soils over bedrock [J]. Oecologia,134:46-54.
SCHENK HJ,ESPINO S,ROMO DM,et al.,2017. Xylem surfactants introduce a new element to the Cohesion-Tension Theory [J]. Plant Physiol,173(2):1177. Doi:10.1104/pp.16.01039.
SCHENK HJ,JACKSON RB,2002. Rooting depths,lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems [J]. J Ecol,90(3):480-494.
SCHWINNING S,2010. The ecohydrology of roots in rocks [J]. Ecohydrology,3(2):238-245.
SCHWINNING S,KELLY CK,2013. Plant competition,temporal niches and implications for productivity and adaptability to climate change in water-limited environments [J]. Funct Ecol,27(4):886-897.
SELEIMAN MF,AL-SUHAIBANI N,ALI N,et al.,2021. Drought stress impacts on plants and different approaches to alleviate its adverse effects [J]. Plants,10(2):259.
SILLMANN J,THORARINSDOTTIR T,KEENLYSIDE N,et al.,2017. Understanding,modeling and predicting weather and climate extremes:Challenges and opportunities [J]. Weath Clim Extr,18:65-74.
SUKHOVA E,AKINCHITS E,SUKHOV V,2017. Mathematical models of electrical activity in plants [J]. J Membr Biol,250:407-423.
SUN L,YANG L,CHEN LD,et al. 2018. Hydraulic redistribution and its contribution to water retention during short-term drought in the summer rainy season in a humid area [J]. J Hydrol,566:377-385.
TAN FS,SONG HQ,LI ZG,et al.,2019. Hydraulic safety margin of 17 co-occurring woody plants in a seasonal rain forest in Guangxi’s Southwest karst landscape,China [J]. Chin J Plant Ecol,43(3):227-237. [譚凤森,宋慧清,李忠国,等,2019. 桂西南喀斯特季雨林木本植物的水力安全 [J]. 植物生态学报,43(3):227-237.]
TAN FS,SONG HQ,FU PL,et al. 2020. Hydraulic safety margins of co-occurring woody plants in a tropical karst forest experiencing frequent extreme droughts [J]. Agric For Meteorol,292-293:108107.
WANG R,WU YY,XING DK,et al.,2017. Biomass production of three biofuel energy plants’ use of a new carbon resource by carbonic anhydrase in simulated karst soils:Mechanism and capacity [J]. Energies,10(9):1370-1383.
WEST AG,DAWSON TE,FEBRUARY EC,et al.,2012. Diverse functional responses to drought in a Mediterranean-type shrubland in South Africa [J]. New Phytol,195(2):396-407.
WU YY,2011. Trategies to increase carbon fixation and sequestration by karst-adaptable plant [J]. Carsol Sin,30(4):461-465. [吴沿友,2011. 喀斯特适生植物固碳增汇策略 [J]. 中国岩溶,30(4):461-465.]
XIAO S,LIU LT,ZHANG YJ,et al.,2020. Review on new methods of in situ observation of plant micro-roots and interpretation of root images [J]. J Plant Nutr Fert [J]. 26(2):370-385. [肖爽,刘连涛,张永江,等,2020. 植物微根系原位观测方法研究进展 [J]. 植物营养与肥料学报,26(2):370-385.]
YANG FT,FENG ZM,WANG HM,et al.,2017. Deep soil water extraction helps to drought avoidance but shallow soil water uptake during dry season controls the inter-annual variation in tree growth in four subtropical plantations [J]. Agric For Meteorol,234:106-114.
YANG ZH,SONG XQ,SU WC,2019. Slope runoff process and its utilization technology in Southwest karst area [J]. Earth Sci,44(9):2931-2943. [杨振华,宋小庆,苏维词,2019. 西南喀斯特地区坡地产流过程及其利用技术 [J]. 地球科学,44(9):2931-2943.]
YUAN DX,2014. Research on major environmental geological problems and countermeasures in karst rock mountain area in Southwestern China [M]. Beijing:Science Press. [袁道先,2014. 西南岩溶石山地区重大环境地质问题及对策研究 [M]. 北京:科学出版社.]
YUAN ZL,PAN XY,JIN W,2019. Tree-associated symbiotic microbes and underlying mechanisms of ecological interactions:a case study of poplar [J]. Acta Ecol Sin,39(1):381-397. [袁志林,潘雪玉,靳微,2019. 林木共生菌系统及其作用机制——以杨树为例 [J]. 生态学报,39(1):381-397.]
YUN SX,QIN HN,2014. Status quo and protection countermeasures of plant resources in Dian-Qian-Gui Karst area [C]. Shenyang:11th National Symposium on Biodiversity Science and Conservation:293-303. [于胜祥,覃海宁,2014. 滇黔桂喀斯特地区植物资源现状与保护对策 [C]. 沈阳:第十一届全国生物多样性科学与保护研讨会:293-303.]
ZHANG QW,ZHU SD,JANSEN S,et al.,2021. Topography strongly affects drought stress and xylem embolism resistance in woody plants from a karst forest in Southwest China [J]. Funct Ecol,35(3):566-577.
ZHANG ZF,YOU YM,HUANG YQ,et al.,2012. Effects of drought stress on Cyclobalanopsis glauca seedlings under simulating karst environment condition [J]. Acta Ecol Sin,32(20):6318-6325. [张中峰,尤业明,黄玉清,等,2012. 模拟喀斯特生境条件下干旱胁迫对青冈栎苗木的影响 [J]. 生态学报,32(20):6318-6325.]
ZHANG ZF,ZHANG JC,XU GP,et al.,2018. Contribution of arbuscular mycorrhizal fungi to water absorption by Cyclobalanopsis glauca [J].Chin J Eccol,37(8):2285-2292. [張中峰,张金池,徐广平,等,2018. 接种菌根真菌对青冈栎水分吸收的贡献 [J]. 生态学杂志,37(8):2285-2292.]
ZHU SD,CHEN YJ,FU PL,et al.,2017. Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability [J]. Tree Physiol,37(11):1469-1477.
ZHU SD,LIU H,XU QY,et al.,2016. Are leaves more vulnerable to cavitation than branches? [J]. Funct Ecol,30(11):1740-1744.
(责任编辑 李 莉)
