昆虫离子型受体及其功能研究进展
2021-01-12郭金梦董双林
郭金梦, 董双林
(南京农业大学植物保护学院, 南京 210095)
昆虫化学感受(包括味觉和嗅觉)在昆虫觅食、寻找配偶和选择产卵部位以及躲避天敌等行为中起到重要的作用(Hansson and Stensmyr, 2011)。已知的昆虫化学感受相关蛋白主要包括气味结合蛋白(odorant binding protein, OBP)、气味受体(odorant receptor, OR)、化学感受蛋白(chemosensory protein, CSP)、味觉受体(gustatory receptor, GR)、感觉神经元膜蛋白(sensory neuron membrane protein, SNMP)、离子型受体(ionotropic receptor, IR)和气味降解酶(odorant degrading enzyme, ODE)等(Eyunetal., 2017)。
IR由离子型谷氨酸受体(ionotropic glutamate receptor, iGluR)家族演化而来,是一类保守的配体门控离子通道(ligand-gated ion channel),在配体分子与其结合后通道打开,导致膜内外离子的流动从而产生膜电位。Benton等(2009)基于生物信息学手段从黑腹果蝇Drosophilamelanogaster基因组中鉴定了61个IR,此后随着基因组和转录组测序技术的发展,其他很多种昆虫的IR也相继被鉴定。然而,有关IR的功能研究主要集中在果蝇,其他昆虫中很少报道。早期研究证实,昆虫IR是酸、胺等挥发性物质的受体,但近年发现IR还可以感知其他气味物质,并参与包括味觉、温度感受和湿度感受等嗅觉以外的感受。本文从结构、进化、基因鉴定、感器和组织表达谱以及功能等方面,综述了昆虫IR的研究进展,以期为深入研究昆虫IR的功能、作用机制及在害虫防治中的应用提供参考。
1 IR的结构特征及进化
1.1 IR的结构特征
根据序列特征、组织表达谱和系统发育关系,IR分为两个亚家族:触角IR(antennal IR)和趋异IR(divergent IR)(Crosetetal., 2010)。触角IR主要或仅在触角中表达(Crosetetal., 2010);趋异IR主要在触角以外的组织中表达,在种内和种间的保守性均较低。基于Clustal Omega多序列比对分析表明,果蝇中42个趋异IR(Crosetetal., 2010)间的氨基酸序列一致性低于55.48%, 42个果蝇趋异IR与18个棉铃虫Helicoverpaarmigera趋异IR(Liuetal., 2018)间的氨基酸序列一致性低于24.48%。每个IR至少需要一个共受体(co-receptor)才能发挥功能,IR共受体类似于嗅觉共受体(odorant receptor co-receptor, Orco),只能和IR共表达来行使功能;迄今发现4个IR共受体(IR8a, IR25a, IR76b和IR93a),在种间比较保守(Bentonetal., 2009; Silberingetal., 2011; Aietal., 2013)。
IR由iGluR进化而来,两者结构类似。IR的结构包括胞外的N端(N)和由S1与S2组成的配体结合域(ligand-binding domain, LBD), 3个跨膜结构域(TM1-TM3),离子通道孔(P),以及胞内C端(C)。在果蝇的66个IR中,共受体IR8a和IR25a与iGluR相同,其N端还有一个氨基端结构域(amino-terminal domain, ATD);IR21a, IR40a, IR64a和IR93a的N端较长(约400个氨基酸),但与ATD的相似性极低;其他IR的N末端很短(约200个氨基酸),缺少ATD序列(图1)。果蝇66个IR间氨基酸序列的一致性约为10%~70%,因此不同IR可能具有不同的功能特点(Bentonetal., 2009; Rytzetal., 2013)。
与另外两类化感受体OR和GR相比,IR有以下特点:(1)基因数目较少,通常低于相应物种中OR和GR的数目(一些蜚蠊目昆虫除外);(2)跨膜结构域较少,只有3个,而OR和GR通常有7个;(3)开放阅读框较长,多于500个甚至高达900多个氨基酸(Liuetal., 2018),但OR和GR一般少于500个氨基酸。
1.2 IR的进化及起源
基因复制是物种获得新基因的主要途径,不仅导致物种间基因数量的变异,还促进了新种的形成。通过非等位基因同源重组形成的基因复制,是IR等大多数化感受体基因扩增的普遍机制。研究发现,鳞翅目IR家族中发生了大量的基因复制事件,表现为多个基因在同一条染色体上紧密串联,并且几乎所有的触角IR都是单拷贝(Liuetal., 2018)。此外,IR的基因丢失和假基因化现象也被证实(Crosetetal., 2010)。
比较基因组学研究可以揭示IR的起源。分析发现,IR存在于所有被研究的原口动物,包括蜕皮动物(例如线虫和节肢动物)和冠轮动物(例如软体动物和环节动物),但在后口动物、刺胞动物门和扁盘动物门中不存在(Crosetetal., 2010; Rytzetal., 2013)。不同物种中IR的数目差异很大,从秀丽隐杆线虫Caenorhabditiselegans中的3个到甲壳动物淡水枝角水蚤Daphniapulex中的85个(Rytzetal., 2013)以及德国小蠊Blattellagermanica中的800多个(Robertsonetal., 2018a)。iGluR广泛存在于真核生物包括所有被分析的后生动物(Metazoa)中,而IR25a是与iGluR序列最相似的IR,并且在原口动物中高度保守,推测其可能是IR家族中最古老的成员(Crosetetal., 2010)。
早期研究认为,IR40a,IR21a,IR76b和IR8a为昆虫特有(Crosetetal., 2010),但后来的研究发现,这些所谓的“昆虫特异性”触角IR的同源基因也存在于昆虫以外的动物中(Eyunetal., 2017)。如蜈蚣Strigamiamaritima中有IR8a和IR40a(Chipmanetal., 2014),蜘蛛Dysderasilvatica中有IR21a和IR40a(Vizuetaetal., 2017),真宽水蚤Eurytemoraaffinis和智利鱼虱Caligusrogercresseyi中有IR8a和IR21a,鲤锚头蚤Lernaeacyprinacea中有IR76b(Eyunetal., 2017)。利用PhyML软件采用最大似然法,我们对上述智利鱼虱、真宽水蚤、鲤锚头蚤和蜈蚣S.maritima4种非昆虫动物(根据文献未找到蜘蛛的相应序列)及6种昆虫的同源基因的氨基酸序列进行了进化分析,发现所有昆虫的同源基因聚在一起,而其他动物的聚在一起,与其系统亲缘关系相一致(图2)。这类基因的进化及其功能分化值得深入研究。
2 昆虫IR的鉴定
随着测序技术和生物信息学工具的快速发展,越来越多的昆虫IR得以鉴定,这些昆虫涉及鞘翅目、鳞翅目、双翅目以及膜翅目等至少11个目160多种。表1列出了基于基因组数据鉴定的67种昆虫的IR的数量。在这些IR中,只有黑腹果蝇等个别昆虫的部分IR的功能得到研究,涉及嗅觉、味觉、听觉及温湿度感受(详见下文“昆虫IR的功能”)。另外,这些IR大多是基于基因组测序数据通过生物信息学分析所得,需要进一步进行克隆验证。从得到的基因组数据看,不同昆虫IR基因的数量不同,东方蜜蜂Apiscerana、西方蜜蜂Apismellifera只有10个IR基因,德国小蠊中达到惊人的897个,但其中393个是假基因。在目水平上,半翅目和膜翅目的IR数量较少(丽蝇蛹集金小蜂Nasoniavitripennis例外),一般20个左右;鳞翅目30~40个左右;蜚蠊目最多,5种昆虫中最少的也有75个;双翅目不同种昆虫的IR数量变化较大(表1)。
3 昆虫IR基因的感器和组织表达谱
IR基因在不同感器中的表达以果蝇中研究得最为清楚。果蝇成虫触角上着生有毛形感器(trichoid sensillum)、锥形感器(basiconic sensillum)和腔锥形感器(coeloconic sensillum)3种感器;腔锥形感器又分为4个亚型(ac1-ac4),每个腔锥形感器含有2~3个嗅觉感觉神经元(olfactory sensory neuron, OSN)(Yaoetal., 2005)。大量研究表明,果蝇触角表达有16个IR,其中10个仅在腔锥形感器神经元中表达,它们单独或2~3个共表达于一个亚型中(Bentonetal., 2009; Crosetetal., 2010; Rytzetal., 2013);4个IR基因(IR21a,IR40a,IR64a和IR93a)不在腔锥形感器中表达,而在触角芒(arista)和感觉囊 (sacculus) 神经元中表达; 另外2个共受体基因(IR8a和IR25a)在感觉囊和触角的很多腔锥形感器中广泛表达,IR25a还在触角上的锥形感器和毛形感器中有微量表达(Bentonetal., 2009; Abuinetal., 2011)。此外,在果蝇唇瓣中,IR56a与GR66a在某些苦味感受的神经元中共表达,GR5a,IR56b和IR56d在一些糖敏感的神经元中共表达(Kohetal., 2014)。在其他昆虫中,棉铃虫的IR8a和IR25a在成虫触角的腔锥形感器中表达(张夏瑄, 2018);中红侧沟茧蜂Microplitismediator的IR8a则表达于具有嗅觉功能的板形感器内(Wangetal., 2016)。
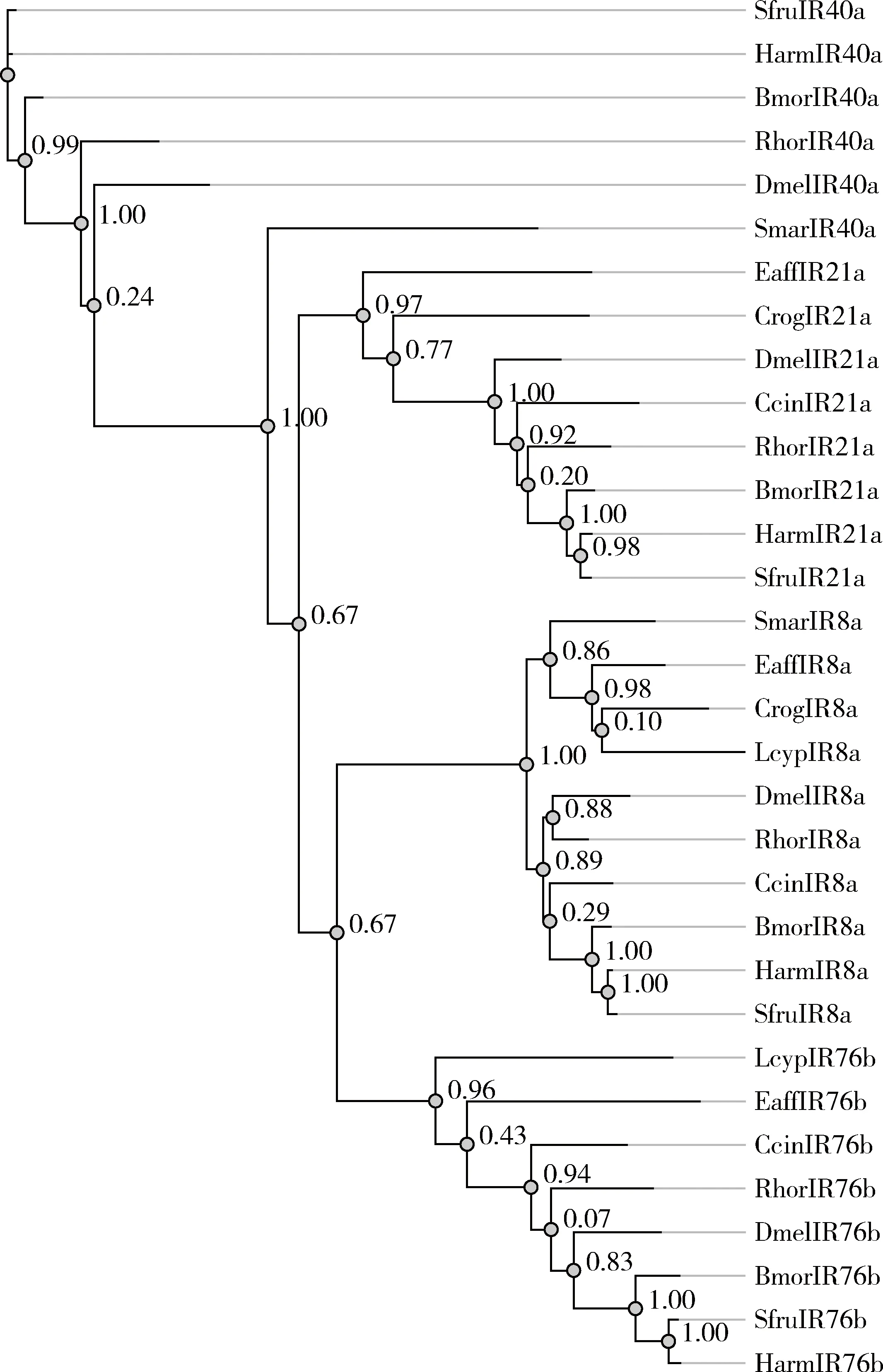
图2 基于氨基酸序列最大似然法构建的不同物种的IR8a, IR21a, IR40a和IR76b的系统发育树
触角IR基因主要在触角中表达,而趋异IR基因主要在触角以外的组织中表达。果蝇趋异IR基因中的IR20分支有35个IR基因,它们在唇瓣、足、咽和翅等不同的组织表达,其中IR47a,IR56a,IR56b,IR56d和IR94e在唇瓣中表达,IR52a在翅和足中表达(Kohetal., 2014; Heetal., 2019)。在亚洲小车蝗Oedaleusasiaticus中,IR25a和IR76b在主要化感组织中高表达,IR8a仅在触角、下唇、下颚须和腹部有少量表达,其他3个IR基因(IR1,IR2和IR3)在大多被测组织中都有较高的表达(Zhouetal., 2019)。在棉铃虫中,80%以上的触角IR基因在触角中高表达(Liuetal., 2018)。在甜菜夜蛾Spodopteraexigua中,9个IR基因主要在触角中表达,其中IR1.1和IR93a在雄虫触角中高表达,IR75p.1和IR41a在雌虫触角中表达较高(Duetal., 2018b)。在疆夜蛾Peridromasaucia中,IR75d和IR75q.2在雄虫触角中表达量较高,IR2,IR60a和IR68a在雌虫触角中表达较高(Sunetal., 2020)。在东方粘虫Mythimnaseparata中,IR93a在嗅觉和非嗅觉组织中都有表达,但大多数IR基因主要在触角中表达(Duetal., 2018a)。在小地老虎Agrotisipsilon中,14个IR基因主要在触角中表达,其中IR12为雄虫特异表达(Guetal., 2014)。IR基因不同的组织表达谱及性别差异,表明这些基因担负不同的生理功能。

表1 基于基因组数据鉴定到的昆虫离子型受体(IR)的数量(截至2020年3月)

续表1 Table 1 continued
4 昆虫IR的功能研究方法
昆虫IR的功能研究方法分为体外和体内两类。体外方法常用的有细胞系(人胚肾上皮细胞HEK293或昆虫Sf9细胞系)和爪蟾卵母细胞(Xenopusoocyte)表达系统;体内常采用果蝇空神经元(empty neuron system)系统、RNA干扰和CRISPR-Cas9系统。
细胞系表达方法是通过将候选IR基因表达载体转染到细胞系,结合钙离子成像技术或膜片钳技术检测化合物刺激引起的细胞兴奋信号,从而获得IR的配体(Zhangetal., 2013)。爪蟾卵母细胞表达系统则是在非洲爪蟾卵母细胞中表达IR基因,通过双电极电压钳系统(two-electrode voltage clamp, TEVC)记录卵母细胞对不同化合物刺激的电生理反应,获得IR的配体(Aietal., 2013; Shanetal., 2019)。体外研究方法相对简单,但偏离昆虫的活体生理环境,所得结果有时不能完全反映其真实情况。
体内研究技术中,果蝇空神经元系统是利用GAL4/UAS系统将外源候选IR基因特异性地表达在果蝇的空神经元中,再利用单感器记录(single-sensillum recording, SSR)技术测定转基因后神经元的电生理反应,确定IR的配体化合物(Abuinetal., 2011; Grosjeanetal., 2011; Prieto-Godinoetal., 2017; Rimaletal., 2019)。该系统应用于果蝇以外特别是双翅目以外昆虫的IR基因的研究时,同样存在偏离本种昆虫生理环境的缺点。RNA干扰是将靶基因同源的双链RNA或者小分子RNA导入到细胞内,干扰目的基因mRNA的翻译使蛋白的表达量下降(Liuetal., 2010)。RNA干扰操作简便,但该方法很难彻底沉默目的基因的表达,并且在鳞翅目等昆虫中的效果较差。CRISPR-Cas9技术则是在DNA水平上对目的基因进行定点突变,从而彻底沉默目的基因的表达,所得结果更为明确和可靠,但步骤较为复杂,需要经过多代筛选来获得纯合的昆虫突变品系(Rajietal., 2019; Zhangetal., 2019)。
5 昆虫IR的功能
5.1 嗅觉功能
IR可以特异性感受某些气味,如氨气、1,4-丁二胺、苯乙醛、丙醛和己醇等(Bentonetal., 2009)。研究发现,氨和挥发性胺对果蝇的吸引作用由表达IR92a的神经元介导,沉默表达IR92a的神经元或IR92a基因降低了胺诱发的生理和行为反应(Minetal., 2013)。多胺(尤其是腐胺、精胺和亚精胺)为细胞生长和增殖等必需,对于生殖过程尤其重要。多胺的含量在果实收获后和发酵过程中显著增加,因此果蝇喜食熟透或发酵的果实,摄入多胺可以提高其产卵量和孵化率。IR41a和IR76b共表达的神经元对于调控多胺类物质对果蝇的吸引作用必不可少(Hussainetal., 2016)。
酸类气味的感受对于昆虫选择取食和产卵场所至关重要。果蝇对酸类化合物的反应,在化合物低浓度时常表现为趋向,而高浓度时为忌避(Aietal., 2010; Chen and Amrein, 2017)。果蝇的4种腔锥形感器(ac1-ac4)均感受各种酸分子。在ac1中,IR31a和IR8a共表达的神经元对2-氧代戊酸有反应;在ac2中, IR75a和IR8a共表达的神经元对乙酸有强烈反应;在ac3中,IR75b和IR75c分别感知丁酸和丙酸,IR75b的LBD结构中单一氨基酸(T523S)的差异导致其对己酸敏感;在ac4中,IR84a和IR8a共表达的神经元对苯乙酸和苯乙醛有反应(Abuinetal., 2011; Silberingetal., 2011; Prieto-Godinoetal., 2017)。此外,果蝇唇瓣苦味感受神经元中IR7a的缺失,显著降低果蝇对乙酸的反应,但对其他有机酸(乙醇酸、柠檬酸、乳酸和丙酸)无影响(Rimaletal., 2019)。
果蝇的求偶行为由fruitless(fru)基因控制,翅中大多数表达IR52a的神经元都表达fru。雄虫IR52a的缺失导致雌雄间交配延迟、交配成功率降低,推测IR52a具有感受雌虫和雄虫信息素的作用(Heetal., 2019)。二氧化碳是昆虫寻找宿主、产卵场所和食物的重要线索,但果蝇只有在觅食状态时才被二氧化碳吸引,其中共受体IR25a为必需(van Breugeletal., 2018)。
除果蝇外,最近对其他昆虫的研究逐渐增多。糖醋液引诱是诱捕和监测粘虫等蛾类昆虫的常用方法,其中乙酸是关键成分。在东方粘虫4种乙酸敏感的感器(as1, as2, as3和as4)中,IR8a总是与另外3个IR(IR64a, IR75q.1和IR75q.2)中的一个共表达,推测IR8a作为共受体参与对乙酸的感受(Tangetal., 2020)。在烟草天蛾中,Zhang等(2019)使用CRISPR-Cas9系统分别敲除Orco,IR8a和IR25a发现,IR8a-/-雌蛾对同种幼虫粪便中的抑卵信息素(3-甲基戊酸和己酸)的反应降低,从而不再避开被幼虫取食过的叶片。雌蚊靠一系列信号(二氧化碳、体味、热量、水分和视觉信号等)来寻找人类宿主,Raji等(2019)通过CRISPR-Cas9将埃及伊蚊的IR8a敲除后,蚊子不再被人体气味中的乳酸等酸性成分所吸引。
在天敌昆虫中红侧沟茧蜂中,Shan等(2019)利用爪蟾卵母细胞表达系统测定发现,IR64a1/IR8a对含6~8个碳原子的酸、醛、酯和醇等化合物有反应;IR64a2/IR8a则对更长碳链的化合物敏感;此外,IR64a2/IR8a还对棉铃虫的性信息素成分Z9-14∶Ald敏感,暗示其在识别寄主昆虫中的作用。
5.2 味觉功能
昆虫依靠味觉来识别多种多样的化合物,从而评估潜在的营养成分、毒素、产卵地点、配偶、病原体、猎物和捕食者。多项研究表明,IR可以作为独立的GR起作用。食盐对动物有益,但过量则有害。Zhang等(2013)发现了黑腹果蝇使用不同类型的味觉受体神经元(gustatory receptor neuron, GRN)对不同浓度的盐进行味觉编码,其中含有IR76b的GRN用于对低盐的检测,引起果蝇的嗜食。
氨基酸是重要的营养物质。Croset等(2016)在果蝇中的研究发现,不同浓度的氨基酸对幼虫的作用不同:吸引、驱避或者不引起反应。IR76b在果蝇幼虫的GRN中广泛表达,作为共受体参与这些氨基酸引起的行为反应。Ganguly等(2017)研究发现,IR20a和IR76b共表达可以引起神经元对氨基酸的反应,但却抑制IR76b对盐的反应。此外,IR56d, IR25a和IR76b共表达参与对碳酸溶液和脂肪酸的反应(Sánchez-Alcaizetal., 2018)。
此外,果蝇IR56a和味觉受体GR66a在一些苦味感觉神经元中共表达,GR5a与IR56b和IR56d在一些糖感觉神经元中共表达。因此,某些神经元的味觉特性可能由GR和IR共同决定(Kohetal., 2014)。
5.3 听觉功能
在果蝇中,多个IR在触角的江氏器(Johnston’s organ)中表达,其中缺失IR64a和IR94b的突变果蝇不影响江氏器的功能,而缺失IR75a的果蝇明显影响了江氏器敏感性,表明IR参与调节江氏器神经元的功能,进而参与听觉过程(Senthilanetal., 2012)。
5.4 温度感受
Chen等(2015)研究发现,果蝇的共受体IR25a在感受温度的微小变化中起重要作用。在果蝇幼虫中,背部低温感觉细胞(dorsal organ cool cell, DOCC)是一组特殊的温度敏感神经元,可以感知每秒几千分之一度的微小温差,在避冷行为中起关键作用。研究发现,IR93a与IR21a和IR25a的共表达介导了DOCC对寒冷的感知(Knechtetal., 2016; Nietal., 2016)。Budelli等(2019)还发现,果蝇成虫触角芒中的“降温感觉细胞”(cooling cell)也由IR93a, IR21a和IR25a所介导,沉默其中的任何一个IR,均降低果蝇对高温(31℃)和低温(16℃)的避开。在冈比亚按蚊Anophelesgambiae中,IR21a是寻找热源的关键受体(Greppietal., 2020)。
5.5 湿度感受
环境湿度影响昆虫的适宜性和地理分布。在果蝇的感觉囊室中,有一组神经元包括干细胞(dry cell)和湿细胞(moist cell),它们分别对干燥和潮湿的空气敏感,从而共同感知外界环境的湿度变化。进一步研究发现,干细胞的功能需要IR40a和2个共受体(IR25a和IR93a)来介导(Enjinetal., 2016; Knechtetal., 2016),湿细胞则需要IR68a和2个共受体的参与(Knechtetal., 2017)。因此,果蝇的湿度感受是由依赖于IR68a的潮湿感受和依赖于IR40a的干燥感受联合完成的。
6 小结与展望
综上所述,IR是一类新发现的昆虫感觉受体,借助于快速发展的组学技术和基因功能分析技术,已经在很多昆虫中鉴定到大量的IR基因并初步揭示了IR基因的多样性功能。今后应在以下几方面加强研究:首先,与气味受体(OR)和味觉受体(GR)相比,获得功能鉴定的离子型受体(IR)的数量较少且主要限于果蝇,其他昆虫尤其是重要农林害虫中IR的功能需要进一步探究。其次,IR功能涉及嗅觉、味觉以及温湿度感受等,虽然已经鉴定了部分IR的功能,但具体的作用机制仍不清楚,需要加强研究。第三,参与多种分子感受的IR及其共受体已被证实,但同一IR介导两种或多种嗅觉(或味觉)分子的模式机制尚不清楚,对IR及其与共受体复合体三级结构的解析方面有待突破。第四,昆虫通过不同类型的受体(GR, IR和OR等)来感觉化学刺激,需要明确单个神经元中不同类型受体间的调控机制。最后,在解析IR结构及功能的同时,应加强基于IR的活性化合物的筛选技术及转基因等新型行为调控技术,为害虫的可持续防治提供支撑。
