藏药麻花秦艽防治细胞因子风暴活性成分与作用机制的网络药理学研究
2021-01-05高小敏刘杜霞张晓英
高小敏,徐 雅,刘杜霞,张晓英,李 捷,仝 敏,赵 勤
藏药麻花秦艽防治细胞因子风暴活性成分与作用机制的网络药理学研究
高小敏,徐 雅,刘杜霞,张晓英,李 捷,仝 敏,赵 勤*
西藏民族大学 藏药检测技术教育部工程研究中心,西藏民族大学医学院藏药筛选实验室,陕西 咸阳 712082
应用网络药理学方法对藏药麻花秦艽防治细胞因子风暴的主要活性成分及可能作用机制进行预测分析。通过中药系统药理学数据库分析平台(TCMSP)及人类基因(GeneCards)数据库收集麻花秦艽的活性化合物和与细胞因子风暴相关靶点;应用Cytoscape 3.6.1软件构建麻花秦艽活性成分-细胞因子风暴靶点调控网络图;构建蛋白-蛋白相互作用网络图筛选核心靶点,利用DAVID在线数据库对核心蛋白进行基因本体论(gene ontology,GO)功能富集分析和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析。筛选得到麻花秦艽与防治细胞因子风暴相关的14个活性化合物和31个有效靶点,核心化合物和靶点分别为熊果酸、木犀草素、山柰酚、豆甾醇、β-谷甾醇和白细胞介素6(interleukin- 6,IL-6)、信号转导子与转录激活子3(signal transducers and transcriptional activators 3,STAT3)、c-Jun氨基末端激酶(C-jun amino-terminal kinase,JNK)、蛋白激酶B1(protein kinase B1,Akt1)、表皮生长因子受体(epidermal growth factor receptor,EGFR)、基质金属蛋白酶(matrix metallopeptidase 9,MMP9)、血管内皮生长因子(vascular endothelial growth factor A,VEGF)、IL-1β、IL-10等。GO分析发现麻花秦艽在抗细胞因子风暴时主要涉及调控细胞凋亡、基因表达、细胞因子活性、一氧化氮生物合成过程的正调控、生长因子活性、免疫应答等过程。KEGG通路分析发现其主要涉及细胞因子风暴相关通路T细胞受体信号通路、酪氨酸激酶JAK和转录因子STAT(Janus kinase-signal transducer and activator of transcription,Jak-STAT)信号通路、低氧诱导因子1(hypoxia-inducible factor 1,HIF-1)信号通路、肿瘤坏死因子信号通路、Toll样受体信号通路、磷酸化磷酯酰肌醇3激酶(phosphatidylinositide 3-kinases,PI3K)-Akt信号通路等核心靶点通路。通过网络药理学数据挖掘初步预测了藏药麻花秦艽防治细胞因子风暴的潜在活性成分及其可能机制,揭示其可通过多成分、多靶点、多通路防治细胞因子风暴对机体造成的损伤,以期为藏药麻花秦艽在防治细胞因子风暴方面的应用提供参考,为开发藏药麻花秦艽的新药用价值提供思路。
麻花秦艽;细胞因子风暴;高细胞因子血症;新型冠状病毒肺炎;网络药理学;熊果酸;木犀草素;山柰酚;豆甾醇;β-谷甾醇
2019年12月世界范围出现了不明原因的新型冠状病毒肺炎,并迅速蔓延。WHO将此病毒命名为严重急性呼吸综合征冠状病毒2(severe acute respiratory syndrome coronavirus 2,SARS-COV-2),所致的疾病命名为新型冠状病毒肺炎(coronavirus disease 2019,COVID-19)[1-2]。临床治疗过程中发现感染了SARS-COV-2的重症患者或轻症患者病情突然恶化时,患者体内出现大量的白细胞介素6(interleukin-6,IL-6)、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、γ干扰素(interferon-γ,IFN-γ)等促炎细胞因子[3-4],呈现细胞因子风暴的特征。细胞因子风暴(cytokine storm)又称高细胞因子血症(hypercytokinemia),是由感染、药物或某些疾病刺激引起的免疫系统过度激活,炎症细胞因子与白细胞在体内不受调控,过度释放,呈现高炎症反应状态,一旦发生可迅速引起急性呼吸窘迫综合征(acute respiratory distress syndrome,ARDS),最终可导致多器官功能衰竭,甚至死亡。多项研究揭示大量且难以控制的细胞因子风暴是导致重症肺炎、全身脏器衰竭的重要原因[5-6]。细胞因子风暴发病机制复杂、疾病进展迅速、死亡率高,寻找有效药物,预防细胞因子风暴发生对防治ARDS具有重大意义。
此次疫情中我国采用中西医联合治疗COVID-19,取得了很好的疗效[7-8],尤其在治疗轻症患者和防治患者病情恶化方面疗效突出,在抑制细胞因子风暴方面有一定的优势。藏医药学是我国传统医学的重要组成部分,此次疫情中也将藏药纳入到一线防疫治疗中,并取得了较好的效果。本课题组正在研究的藏药麻花秦艽Maxim. 是我国西藏高原地产藏药,独特的高原环境孕育了其优良的生物活性,是藏医药中重要的常用药材。藏药麻花秦艽为龙胆科(Gentianaceae)龙胆属(Tourn.) L.多年生草本植物,藏医将其称为“解吉嘎保”,以根和花入药,具有祛风湿、清湿热、止痹痛的功效,常用于胃肠炎、肝炎、胆囊炎等疾病的治疗[9-10]。研究发现麻花秦艽具有良好的抗炎作用,可以改善佐剂和胶原诱导型关节炎模型(collagen-induced arthritis,CIA)大鼠的炎症,降低CIA大鼠血清中炎症因子IL-1β、IL-6和TNF-α水平,抑制滑膜组织中核因子-κB(nuclear factor- κB,NF-κB)p65蛋白表达,减轻滑膜炎症[11-17]。前期的研究结果也显示麻花秦艽具有抗氧化和抗缺氧作用,能够保护低压缺氧大鼠心、脑、肺组织损伤,降低低压缺氧大鼠肺组织上清液中IL-6、IL-1β、TNF-α的含量,减少低压缺氧大鼠心肌组织mRNA的表达[18]。另有研究报道,麻花秦艽其主要成分龙胆苦苷对于小鼠脓毒症急性肺损伤也具有抗感染及保护肺功能等作用,可显著降低肺部中性粒细胞的浸润及炎性渗出,减少脂质过氧化,降低支气管肺泡灌洗液中TNF-α、IL-6含量,减轻肺部炎症反应[19];明显降低脂多糖(lipopolysac- charide,LPS)致小鼠BALF中炎症细胞及中性粒细胞比例,减轻肺部炎症反应,有一定的抗急性肺损伤作用[13];还可显著降低LPS刺激的小鼠巨噬细胞RAW264.7中一氧化氮的产生[20]。其主要成分獐牙菜苦苷对LPS刺激的小鼠巨噬细胞RAW264.7中TNF-α、IL-6的生成具有抑制作用,机制可能与抑制NF-κB通路相关因子p65、核因子-κB抑制物激酶α(nuclear factor κB inhibitor kinase,IKK-α)的水平有关[21]。综上研究结果提示,麻花秦艽可能是新型抗炎药物的有力潜在候选者,其可能具有防治细胞因子风暴发生的作用。因此,本研究通过网络药理学方法分析麻花秦艽防治细胞因子风暴的活性成分、作用靶点及信号通路等,为后续开发麻花秦艽防治细胞因子风暴临床应用提供思路和方法。
1 方法
1.1 麻花秦艽活性成分、靶点收集
通过中国知网、万方数据库相关文献收集麻花秦艽化学成分,将收集的化学成分通过中药系统药理学数据库与分析数据库(TCMSP http//ibts.hkbu. edu.hk/LSP/tcmsp.php)[22]查找成分及对应靶点,排除无靶点、冗长(结构式复杂、数据库无对应成分及靶点)的成分,对最终纳入研究的麻花秦艽活性成分的对应靶点进行Uniprot数据库标准化。
1.2 药物-疾病靶标预测
以关键词“cytokine storm”“hypercytokinemia”在Genecards数据库[23]检索疾病相关靶点。将麻花秦艽活性成分的相关靶点与细胞因子风暴的靶点取交集,得到麻花秦艽活性成分-疾病的靶标及其韦恩图。
1.3 成分-靶点网络的构建
将筛选的麻花秦艽活性成分和疾病靶点通过 Cytoscape软件(V3.6.1)[24]得到活性成分-靶点的调控网络图,对调控网络图进一步分析得出麻花秦艽防治细胞因子风暴的主要活性成分。
1.4 蛋白-蛋白相互作用(protein-protein interaction,PPI)网络的构建和筛选核心靶标
将“1.2”项中的药物-疾病靶标利用String数据库进行分析,设定种属为“homo sapiens”,最小互作分数值≥0.7,不显示在网络中未连接的节点,得出网络图及输出药物-疾病靶标蛋白相互作用结果。通过R语言(3.6.1)筛选核心靶标并绘制核心靶标的条形图。
1.5 GO功能富集分析和KEGG通路富集分析
将筛选的核心靶点利用DAVID在线数据库[25]进行基因本体论(gene ontology,GO)功能富集分析和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析,结果按值由小到大排序及结果分析,<0.05代表统计学有差异。
2 结果
2.1 麻花秦艽活性成分和靶点的筛选
通过TCMSP数据库,将收集的化学成分根据口服生物利用度(oral bioavailability,OB)≥20%,类药性(drug likeness,DL)≥0.09进行筛选,汇总符合筛选条件的化学成分及对应靶点。因熊果酸[26]、当药苷[27]为麻花秦艽主要成分,且具有较好的抗炎作用[28-29]。因此,将熊果酸和当药苷也纳入此次研究,最终筛选出的化学成分共14个(表1),药物靶点共126个。

表1 麻花秦艽活性成分基本信息
2.2 疾病靶点和药物-疾病靶点的交集
Genecards数据库检索与细胞因子风暴相关的靶点,排除重复后共得到315个靶点。通过R语言软件(V3.6.2)映射麻花秦艽活性成分相关靶点到细胞因子风暴靶点,得出麻花秦艽活性成分与细胞因子风暴的有效靶标共31个(图1和表2)。
2.3 成分-靶点调控网络图
将麻花秦艽防治细胞因子风暴的活性成分及作用靶点导入Cytoscape 3.6.1软件,构建可视化分析网络图,图中的菱形代表基因,圆形代表化学成分。图形的大小及颜色深度与度(degree)呈正相关,度值的大小与该成分在疾病中发挥作用的大小相关,值越大表明该成分治疗疾病的可能性越大。结果可得出熊果酸(DY3)、木犀草素(DY11)、山柰酚(DY12)、豆甾醇(DY10)、β-谷甾醇(DY1)在麻花秦艽防治细胞因子风暴时为主要活性成分(图2)。

图1 麻花秦艽防治细胞因子风暴靶点韦恩图

表2 麻花秦艽防治细胞因子风暴的关键靶点
2.4 PPI网络图和核心靶点
将麻花秦艽与细胞因子风暴映射得到的31个靶点导入String数据库,按“1.4”项中的条件进行设置,得到麻花秦艽-细胞因子风暴相互作用网络图,位于中心且边数越多的节点成为核心靶点的可能性越大(图3)。用R语言(V3.6.2)进一步筛选麻花秦艽-细胞因子风暴靶点相互作用网络,按邻接节点数目由多到少形成条形图(图4)。邻接节点的数量与成为核心基因的概率相关,其数量越多成为核心靶点的概率越大。结果发现IL-6、信号转导子与转录激活子3(signal transducers and transcriptional activators 3,STAT3)、c-Jun氨基末端激酶(C-jun amino-terminal kinase,JNK)、蛋白激酶B1(protein kinase B1,Akt1)、表皮生长因子受体(epidermal growth factor receptor,EGFR)、基质金属蛋白酶(matrix metallopeptidase 9,MMP9)、血管内皮生长因子A(vascular endothelial growth factor,VEGF)、IL-1β、IL-10等与细胞因子风暴关系密切。

菱形代表靶点,圆形代表化学成分

图3 PPI网络图
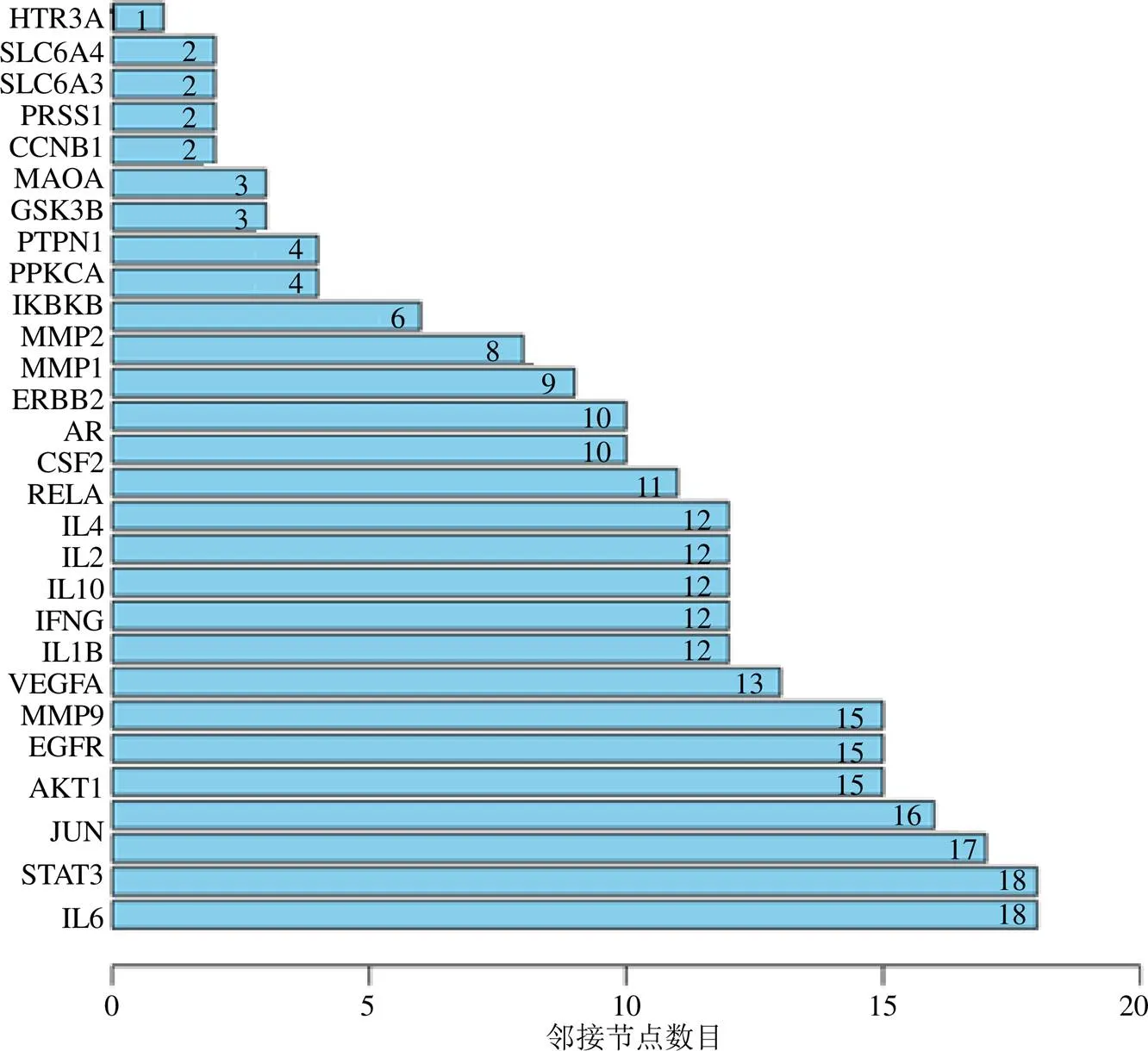
图4 核心靶点条形图
2.5 GO功能富集分析和KEGG通路富集分析
将“2.4”项中的核心靶点利用DAVID数据库进行GO功能富集分析和KEGG通路富集分析。GO功能富集分析共得出231条结果,以生物过程为主,结果显示麻花秦艽防治细胞因子风暴生物过程主要与凋亡、转录、基因表达、一氧化氮生物合成等有关。细胞过程主要与细胞外空隙、细胞外间质有关,分子功能通过影响细胞因子活性、生长因子活性发挥作用(图5、6)。KEGG通路富集分析得出83条相关通路,与细胞因子风暴直接相关的有27条,按照值由小至大排序,分析麻花秦艽防治细胞因子风暴直接相关的前10条信号通路并绘制气泡图,图中圆圈的大小与该通路有关的靶点数呈正相关,颜色越深表示值越小。结果显示,T细胞受体信号通路、酪氨酸激酶JAK和转录因子STAT(Janus kinase-signal transducer and activator of transcription,Jak-STAT)信号通路、低氧诱导因子1(hypoxia- inducible factor 1,HIF-1)信号通路、肿瘤坏死因子信号通路及磷酸化磷酯酰肌醇3激酶(phosphatidylinositide 3-kinases,PI3K)-Akt信号通路可能与麻花秦艽防治细胞因子风暴密切相关(图7)。

图5 GO富集分析饼形图
3 讨论
COVID-19已经在全球迅速蔓延,值得警惕的是,在此次疫情的重症患者体内,发现有大量的炎症因子IL-6、TNF-α、IFN-γ等,目前认为病情加重的原因可能是患者体内启动了“炎症因子风暴”[3]。此外,ICU的患者粒细胞集落刺激因子(granulocytecolony-stimulatingfactor,G-CSF)、趋化因子-10(interferon-inducible protein-10,IP-10)、单核细胞趋化蛋白1(monocyte chemoattractant protein,MCP-1)、巨噬细胞炎性蛋白-1α(macrophage inflammatory protein-1α,MIP-1α)、TNF-α浓度高于非ICU的患者,也说明疾病严重程度与细胞因子风暴相关[3]。因此,早期干预细胞因子风暴可缩短治疗COVID-19疗程,减轻对机体的损伤,减少重症患者、死亡患者数量。现代研究[11-18]揭示藏药麻花秦艽具有较强抗炎作用,可以降低炎症因子IL-1β、IL-6和TNF-α水平,减轻肺部炎症反应,保护急性肺损伤。通过网络药理学方法,本研究预测了麻花秦艽防治细胞因子风暴的核心靶点和活性成分,主要为IL-6、STAT3、JUN、AKT1、EGFR等和木犀草素、熊果酸、β-谷甾醇、山柰酚等。T细胞受体信号通路、PI3k-AKT信号通路、HIF-1信号通路、肿瘤坏死因子信号通路及Jak-STAT信号通路可能与麻花秦艽防治细胞因子风暴密切相关。
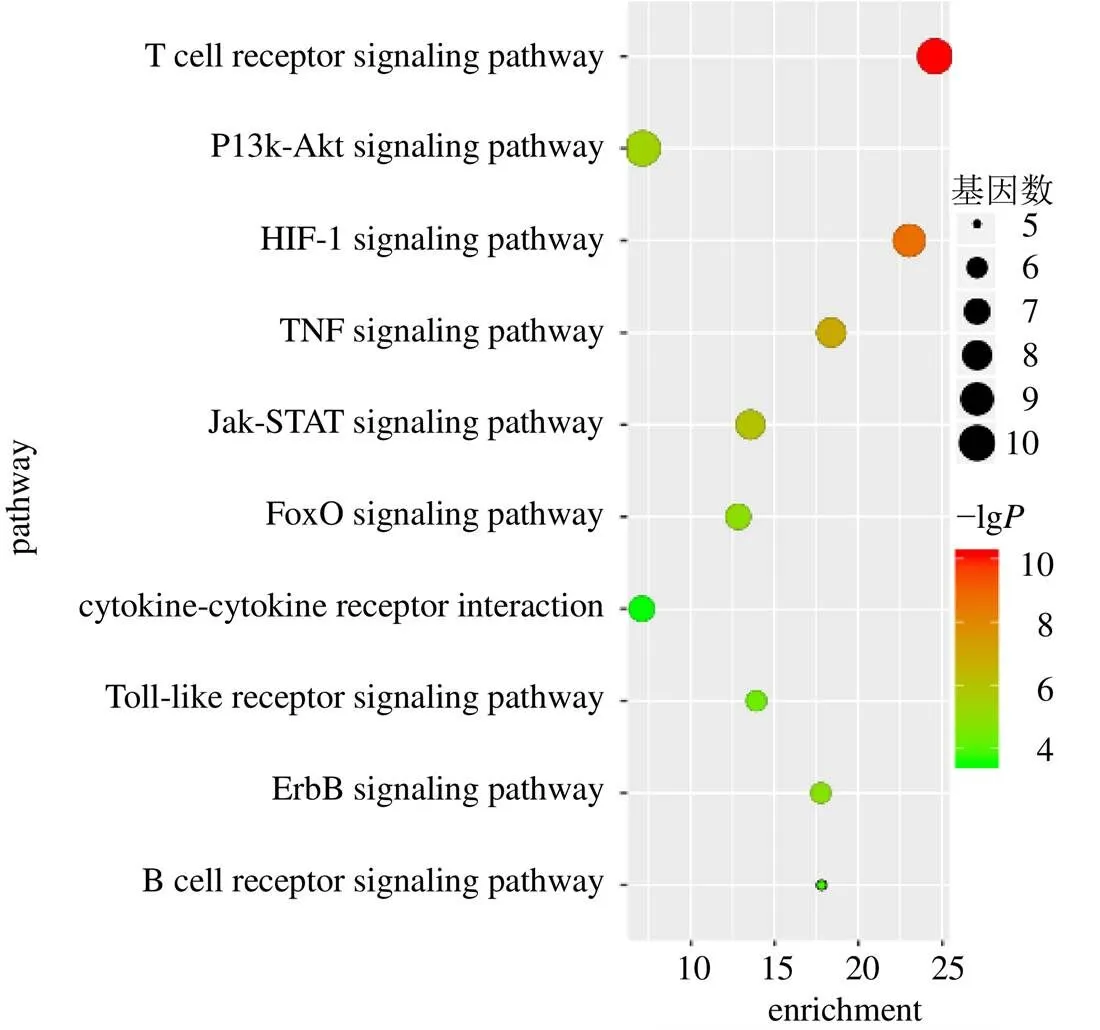
图7 KEGG富集分析气泡图
有研究报道木犀草素可显著降低急性痛风性关节炎大鼠体内Toll样受体2(Toll-like receptor 2,TLR2)、TLR4、髓样分化因子(myeloid differentiation factor 88,MyD88)和NF-κB蛋白的表达,明显减少IL-1β、TNF-α、IL-6细胞因子的释放[30]。其还可通过PI3K-Akt信号通路调节MMP9、MMP1的释放从而抑制小细胞肺癌H1688、H466细胞的增殖与分化,下调VEGF的水平,抑制细胞外基质重塑,减轻炎症因子引起血管内皮细胞过度活化造成的机体损伤[31]。山柰酚可下调乙酰氨基酚诱导的小鼠急性肝损伤p细胞核激酶(Jun nuclear kinase,p-JNK)和内切糖苷酶(endo-glycosidase,Endo G)表达,上调Bcl-2蛋白表达,降低炎症因子IL-1β、IL-6、TNF-α水平[32]。木犀草素、山柰酚均是黄酮类成分,常作为抗细胞因子风暴的主要化学成分,其具有抗炎、降低血管的脆性、改善血管的通透性等作用,还可以通过调节一氧化氮(NO)进一步减轻血管内皮细胞的损伤[33]。熊果酸是一种三萜类天然产物,具有抗氧化、抗炎、调节免疫、抗肿瘤等多种生物学活性[34-36]。β-谷甾醇属于甾醇类,具有调血脂、抗炎、抗肿瘤作用。Valerio等[37]通过LPS诱导的J774A1巨噬细胞,发现β-谷甾醇可升高抗炎因子IL-10的活性,减小趋化因子和促炎因子活性,下调核转录因子的表达。综上所述,麻花秦艽含有的活性成分木犀草素、山柰酚、熊果酸素等具有防治细胞因子风暴的作用,与本研究的推测是一致的。
预测了麻花秦艽防治细胞因子风暴的核心靶点为IL-6、STAT3、JUN、AKT1、EGFR、MMP9、VEGFA、IL-1β、IL-10等。细胞因子风暴是多种促炎因子如白介素家族、肿瘤坏死因子、选择素、黏附素的过高表达,尤其是 IL-6、TNF-α、IL-1β的迅速大量产生打破了免疫平衡,在细胞因子风暴中发挥着重要作用。国家卫生健康委员会、国家中医药管理局最新印发的《新型冠状病毒感染的肺炎诊疗方案(试行第七版)》[2]已经明确将IL-6水平上升作为病情恶化的临床警示指标。TNF-ɑ是细胞因子风暴的核心炎症因子,能够促进多种免疫细胞分化、分泌、杀伤功能,是引起其他细胞因子水平上升和组织损伤的核心。在机体感染病毒时,外周血中TNF-ɑ表达升高,激活T细胞促进IL-1、IL-6、IL-8和IL-12等细胞因子的产生及分泌,从而激活一系列炎性反应,还可激活TNF-ɑ/NF-κB途径参与急性肺损伤的发生[38]。
GO富集分析结果得出,生物过程主要与细胞凋亡、转录,细胞对LPS的反应等有关。细胞组成主要与细胞外空隙功能有关。分子功能通过影响细胞因子活性、生长因子活性发挥作用。机体在肺部感染后,细胞凋亡增加,Bax水平显著升高、Bcl-2水平下降,提示细胞凋亡在感染过程中发挥重要作用[39]。KEGG通路富集分析得出与上述靶点相关的核心通路为T细胞受体信号通路、Jak-STAT信号通路、HIF-1信号通路、肿瘤坏死因子信号通路、Toll样受体信号通路、PI3K-Akt信号通路,这些通路是与细胞因子风暴密切相关的,可能是麻花秦艽防治细胞因子风暴的途径。当抗原侵入人体后与T细胞受体结合,同时白细胞共同抗原抗体45(leukocyte common antigen antibody,CD45)可与CD4或CD8聚集,促使淋巴细胞特异性蛋白酪氨酸激酶抗体P56(lymphocyte-specific protein tyrosine kinase antibody P56,P56 lck)等蛋白酪氨酸激酶(protein yyrosine kinase,PTK)分子去磷酸化,而引发一系列激酶的活化和级联反应,使得活化信号可以逐级下传[40]。JAK-STAT信号通路是参与多种细胞因子信号转导的重要路径,对机体免疫应答、免疫细胞分化发育及炎症反应有重要影响[41]。HIF-1普遍存在于机体各组织和细胞中,当机体发生炎症反应时组织对氧的需求增加,血管血栓的形成或压力增大进一步引起血液的营养来源减少,氧气的供需比率变化使炎症组织低氧情况加重[42]。Toll样受体中的TLR2、TLR4影响炎症因子表达及PI3K-Akt激活在麻花秦艽成分研究中也多次提到。TLR4可识别LPS,并通过TRIF介导TLR4-TRIF或TLR4- MyD88-IKK-NF-κB下游信号通路,产生大量促炎细胞因子和趋化因子介导急性肺炎,由病毒感染坏死细胞释放的高迁移率族蛋白B1(high mobility group B1,HMGB1)可通过TLR4诱导细胞因子分泌,也会导致过度的炎症反应及重症肺炎[43]。而免疫T细胞亚群CD4+/CD8+比例失衡,是激活TLR4-MyD88-IKK-NF-κB信号通路的主要因素,可促使肺泡巨噬细胞大量分泌I型IFN、TNF-α、IL-6、IL-8、IL-1β等炎性细胞因子和MCP-1、MIP-1α、调节活化的正常T细胞表达及分泌的趋化因子(activation regulates expression and secretion of normal T cells,RANTES)、IP-10等趋化因子[44],诱发和加重细胞因子风暴发生急性肺损伤(acute lung injury,ALI)/急性呼吸窘迫(acute respiratory distress,ARDS)。这些研究表明,T细胞受体信号通路、Toll样受体信号通路、PI3K/Akt、JAK/STAT、TNF-α/NF-κB信号途径的激活在细胞因子风暴引起脏器损伤中具有重要的作用,并可以成为潜在的治疗靶点。结合前期对麻花秦艽的免疫作用研究发现,麻花秦艽醇提物可显著降低白细胞、淋巴细胞水平,抑制刀豆蛋白A(concanavalin A,ConA)和LPS诱导的小鼠T、B淋巴细胞增殖,降低CD4+、升高CD8+,使CD4+/CD8+值降低,表明麻花秦艽醇提物可能是通过调整T细胞亚群CD4+和CD8+比例平衡关系来调控小鼠T细胞免疫。这些结果充分显示,麻花秦艽可通过多成分、多靶点及多信号通路发挥抗细胞因子风暴作用。
综上所述,经网络数据挖掘预测得出的麻花秦艽抗细胞因子风暴主要成分、靶点以及信号通路、生物学功能与已经报道文献基本吻合,说明运用网络数据挖掘具有一定的可行性。但是,网络数据挖掘也有一定的缺陷,麻花秦艽中部分成分未在数据库找到而无法进行分析,部分通路仍未证实。总体而言,网络药理学作为一门通过计算机大量数据计算等方法为研究供药物成分、靶点、信号通路的预测具有可信性,在药物研究中提供研究方向指导,为证实药物的作用及挖掘药物新用途提供依据。
利益冲突 所有作者均声明不存在利益冲突
[1] Zhu N, Zhang D Y, Wang W L,. A novel coronavirus from patients with pneumonia in China, 2019 [J]., 2020, 382(8): 727-733.
[2] 国家卫生健康委办公厅. 新型冠状病毒肺炎诊疗方案(试行第七版) [J]. 心肺血管病杂志, 2020, 39(2): 103-107.
[3] Huang C L, Wang Y M, Li X W,. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China [J]., 2020, doi: 10.1016/S0140- 6736(20)30183-5.
[4] 宋霞, 陈涛, 孙晶晶, 等. PCT与IL-6联合检测对新冠病毒感染肺炎的临床价值探讨 [J]. 国际检验医学杂志, 2020, 41(18): 2281-2283.
[5] 尹明星, 曹艳, 施春阳, 等. 中药防治细胞因子风暴的研究进展 [J]. 中草药, 2020, 51(5): 1089-1095.
[6] 郭新坤. HIF-1α核转位调控炎症因子表达在甲型H1N1流感病毒感染致重症肺炎中的作用研究 [D]. 上海: 上海交通大学, 2017.
[7] 何黎黎, 龚普阳, 封玥, 等. 中药在抗新型冠状病毒肺炎 (COVID-19) 引起的细胞因子风暴中的应用分析 [J]. 中草药, 2020, 51(6): 1375-1385.
[8] 王玉亮, 王峰, 耿洁. 细胞因子与细胞因子风暴 [J]. 天津医药, 2020, 48(6): 494-499.
[9] 中国科学院西北高原生物研究所. 藏药志 [M]. 西宁:青海人民出版社, 1991.
[10] 张雅惠. 藏药秦艽花的化学成分研究及中国药典中麻花秦艽品种单列的研究 [D]. 西安: 西北大学, 2009.
[11] 张新新, 贾娜, 孙琛, 等. 大叶秦艽花与麻花秦艽花抗炎镇痛作用的研究 [J]. 西北药学杂志, 2012, 27(4): 341-343.
[12] 包婷雯, 左明丽, 王敏, 等. 藏药麻花秦艽不同部位醇提物的抗炎作用研究 [J]. 中国药房, 2018, 29(22): 3114-3118.
[13] Pan Z, Xiong F, Chen Y L,. Traceability of geographical origin inby UPLC-Q exactive mass and multivariate analyses [J]., 2019, 24(24): E4478.
[14] 王笠, 许亮. 麻花艽的研究进展 [J]. 中国民族民间医药, 2015, 24(6): 17-19.
[15] 赵勤, 王乐乐, 魏立鹏, 等. 麻花秦艽醇提物对佐剂性关节炎大鼠的影响 [J]. 中药药理与临床, 2015, 31(1): 145-147.
[16] 张新新, 贾娜, 孙琛, 等. 大叶秦艽花与麻花秦艽花抗炎镇痛作用的研究 [J]. 西北药学杂志, 2012, 27(4): 341-343.
[17] 贾娜, 崔佳, 文爱东. 藏药麻花秦艽醇提物对胶原诱导型关节炎模型小鼠滑膜组织中NF-κBp65表达的影响 [J]. 中国药房, 2018, 29(15): 2082-2085.
[18] 吴晓军, 李桂新, 徐雅, 等. 麻花秦艽醇提物对高原低氧大鼠肺组织和脑组织的保护作用 [J]. 中药药理与临床, 2019, 35(3): 77-82.
[19] 陈露露, 徐关丽, 袁红梅, 等. 龙胆苦苷对脓毒症所致急性肺损伤小鼠的保护作用 [J]. 第三军医大学学报, 2012, 34(23): 2392-2394.
[20] 张丽, 王永艳, 王应霞, 等. 龙胆苦苷抗急性肺损伤活性研究 [J]. 云南农业大学学报:自然科学, 2018, 33(3): 450-455.
[21] 王君燕, 童晔玲, 赵文慧. 獐牙菜苦苷的抗炎活性及其对NF-κB通路相关因子p65和IKK-α表达的影响 [J]. 中国现代应用药学, 2018, 35(12): 1817-1820.
[22] Ru J L, Li P, Wang J N,. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines [J]., 2014, 6: 13.
[23] Fishilevich S, Nudel R, Rappaport N,. GeneHancer: Genome-wide integration of enhancers and target genes in GeneCards [J]., 2017, doi: 10.1093/database/ bax028.
[24] Otasek, D. Morris, J H, Boucas, J,. Cytoscape Automation: Empowering workflow-based network analysis [J]., 2019. 20(1): D185.
[25] Dennis G J, Sherman B T, Hosack D A,. DAVID: Database for annotation, visualization, and integrated discovery [J]., 2003, 4(9): R60.
[26] 张军文, 李成思, 卢永昌, 等. MEKC测定麻花艽中齐墩果酸和熊果酸含量 [J]. 广西师范大学学报: 自然科学版, 2018, 36(1): 99-104.
[27] 张景瑜, 徐雅, 吴晓军, 等. HPLC法对西藏麻花秦艽正丁醇部位4种环烯醚萜苷类成分的分析 [J]. 中华中医药学刊, 2019, 37(11): 2654-2656.
[28] 陈维维, 张小莉. 熊果酸调控细胞因子法治疗炎症疾病的临床观察 [J]. 中国中医药现代远程教育, 2019, 17(1): 141-143.
[29] 张慧娟, 李菊, 马晓慧, 等. 裂环环烯醚萜苷类化合物的药理作用研究进展 [J]. 药学研究, 2018, 37(11): 659-663.
[30] 沈瑞明, 马丽辉, 郑颜萍. 木犀草素通过TLR/MyD88/NF-κB通路参与急性痛风性关节炎大鼠的抗炎作用 [J]. 中南大学学报: 医学版, 2020, 45(2): 115-122.
[31] 魏付桥, 阳乐彬, 王春云, 等. 木犀草素抑制PI3K/Akt的激活下调结肠癌细胞MMP-9活性 [J]. 中国处方药, 2019, 17(9): 26-27.
[32] 张伟贤, 兰天, 董嘉乐, 等. 山柰酚抵抗对乙酰氨基酚引起的肝细胞损伤研究 [J]. 世界中医药, 2019, 14(1): 39-43.
[33] 白鹭, 李鸿, 覃琴, 等. 黄酮类化合物对血管内皮细胞损伤的保护作用及机制研究进展 [J]. 中国实验方剂学杂志, 2020, 26(12): 203-211.
[34] 沈晓静, 赵红梅, 赵蕾, 等. 齐墩果酸研究进展 [J]. 广州化工, 2019, 47(24): 16-19.
[35] 付亚玲, 高琳, 张东旭, 等. 齐墩果酸和熊果酸提取、分离与测定方法研究进展 [J]. 食品研究与开发, 2020, 41(2): 196-199.
[36] 谭娟, 黄微, 陈善龙, 等. 熊果酸衍生物与查耳酮缀合物的合成及抗炎活性 [J]. 药学学报, 2016, 51(6): 938-946.
[37] Valerio M, Awad A B. β-Sitosterol down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in J774A.1 murine macrophages [J]., 2011, 11(8): 1012-1017.
[38] Cruceriu D, Baldasici O, Balacescu O,. The dual role of tumor necrosis factor-alpha (TNF-α) in breast cancer: Molecular insights and therapeutic approaches [J].(Dordr), 2020, 43(1): 1-18.
[39] 谢丹, 文丹宁, 罗丹. miR-127-5p靶向IRAK4对肺炎链球菌诱导的肺泡上皮细胞凋亡及炎症因子表达的影响 [J]. 临床肺科杂志, 2020, 25(2): 261-266.
[40] 袁璟, 廖玉华, 汪朝晖, 等. 腺苷酸转位酶诱导自身免疫性心肌病小鼠的异常T细胞受体信号通路 [J]. 中国病理生理杂志, 2006, 22(2): 214-218.
[41] 于子涵, 刘英. JAK-STAT1信号通路及细胞因子信号转导抑制蛋白-1在类风湿关节炎中的研究进展 [J]. 风湿病与关节炎, 2014, 3(4): 66-69.
[42] Colgan S P, Eltzschig H K. Adenosine and hypoxia-inducible factor signaling in intestinal injury and recovery [J]., 2012, 74: 153-175.
[43] Creagh E M, O'Neill L A. TLRs, NLRs and RLRs: A trinity of pathogen sensors that co-operate in innate immunity [J]., 2006, 27(8): 352-357.
[44] Le Goffic R, Balloy V, Lagranderie M,. Detrimental contribution of the Toll-like receptor (TLR)3to influenza A virus-induced acute pneumonia [J]., 2006, 2(6): e53.
Active components and mechanism of Tibetan medicineagainst cytokine storm
GAO Xiao-min, XU Ya, LIU Du-xia, ZHANG Xiao-ying, LI Jie, TONG Min, ZHAO Qin
Engineering Research Center of Tibetan Medicine Testing Technology of Ministry of education, Tibetan Medicine Screening Laboratory, School of Medicine, Tibet University for Nationalities, Xianyang 712082, China
To analyze the main active components and possible mechanism of Mahuaqinjiao () in the preventive treatment of cytokine storm by network pharmacology.The therapeutic targets for cytokine storms and its active ingredients ofwere searched and collected from the database of TCMSP and GeneCards; The regulatory network map of active component cytokine storm targets ofwas constructed by using Cytoscape 3.6.1; The network map of protein-protein interaction was constructed to screen the core target, and the gene ontology (GO) function enrichment analysis and KEGG pathway enrichment analysis of the core protein were carried out by using the online database of DAVID.After screening, 14 active components and 31 effective targets for prevention and treatment of cytokine storm were obtained, mainly including IL6, STAT3, Jun, AKT1, EGFR, MMP9, VEGFA, IL-1β, IL-10, and other core targets. Go analysis showed thatmainly involved in the regulation of apoptosis, gene expression, cytokine activity, positive regulation of nitric oxide biosynthesis, growth factor activity, immune response and other processes. KEGG pathway analysis showed that the main pathways related to cytokine storm were T-cell receptor signaling pathway, JAK STAT signaling pathway, HIF-1 signaling pathway, tumor necrosis factor signaling pathway, Toll like receptor signaling pathway, PI3K Akt signaling pathway and other core target pathways.Based on the data mining of network pharmacology, the potential active components and possible mechanism targets ofagainst cytokine storm were predicted preliminarily, in order to provide reference for the application ofin the prevention and control of cytokine storm, and provide ideas for the development of new uses of.
Mahuaqinjiao (Maxim.); cytokine storm; hypercytokinemia; coronavirus disease 2019 (COVID-19); network pharmacology; ursolic acid; luteolin; kaempferol; stigmasterol; β-sitosterol
R285
A
0253 - 2670(2021)01 - 0186 - 10
10.7501/j.issn.0253-2670.2021.01.022
2020-05-11
国家自然科学基金资助项目(81660722);西藏民族大学新冠肺炎疫情应急项目(XZMDYJ02);陕西省中医管理局(2019-ZZ-JC046);西藏民族大学重点项目(13myZP07)
高小敏(1992—),女,硕士在读,研究方向为高原病及藏药药效学。Tel: 18220741795 E-mail: 2456711545@qq.com
赵 勤,教授,硕士生导师,主要从事高原病及藏药药效学研究。E-mail: xyzhaoqin@126.com
[责任编辑 潘明佳]
