根结线虫生防真菌交枝顶孢原生质体的制备与再生体系构建
2020-12-28姚玉荣霍建飞郝永娟
姚玉荣 霍建飞 郝永娟

摘要 本研究以根结线虫生防真菌交枝顶孢Acremonium implicatum为供试菌株,探究了菌龄、降解酶种类、酶解温度和时间、不同渗透压稳定剂、不同pH等因素对其原生质体制备的影响,并建立了交枝顶孢原生质体制备体系;制备的原生质体在SR培养基上再生率达到17.78%。此外,GFP遗传转化菌株的获得充分表明制备的原生质体可作为后续遗传转化材料,为进一步研究交枝顶孢对根结线虫的生防机理奠定基础。
关键词 交枝顶孢; 原生质体制备; 再生; 绿色荧光蛋白
中图分类号: S 435.45; S 476
文献标识码: A
DOI: 10.16688/j.zwbh.2019447
Abstract In order to establish protoplast-mediated genetic transformation of the biocontrol fungus Acremonium implicatum, the conditions for the isolation and regeneration of A.implicatum protoplasts were optimized, including culture time, enzyme system, enzymatic digestion time and temperature, type of osmotic stabilizers, pH value and so on. For the preparation of protoplasts, solid regeneration medium was better with a regeneration rate of protoplasts of 17.78%. The transformant strain with GFP was obtained. The system may build a foundation for further study on the mechanism of biocontrol against the root-knot nematode.
Key words Acremonium implicatum; protoplast preparation; regeneration; GFP
根结线虫Meloidogyne spp.是一类世界性分布的重要植物病原物,给全球农业生产带来了巨大的经济损失[1]。蔬菜生产是农业生产中不可或缺的一部分,随着保护地蔬菜种植面积的逐年增加,根结线虫病害发生日益严重。根结线虫种类繁多,分布广,致病性强且寄主范围广泛、环境适应性强,能侵染几乎所有的蔬菜作物[1-2]。在蔬菜作物中,以茄科、十字花科和葫芦科等受害最重[3]。蔬菜作物被根结线虫侵染后会导致产量降低,品质下降。根结线虫还会与其他病原物形成复合侵染,造成更加严重的经济损失。根结线虫的防治主要为土壤熏蒸剂和化学杀虫剂[4],但是由于使用不规范、危害人类健康、污染环境等问题的频繁出现,越来越多的化学农药被禁止使用[5-6]。寻找高效且环境友好的根结线虫防治措施是当前亟待解决的问题之一。因此应用微生物防治根结线虫已经成为国内外研究的重点内容。木霉Trichoderma spp.[7-8]、拟青霉Paecilomyces spp.[9]以及枝顶孢Acremonium spp.[10-11]等生防菌对根结线虫的防治效果显著。然而,对生防真菌防治线虫机制的研究相对较少,限制了当前生物防治方法的应用。鑒于这种情况,需要加深对生防真菌防控机理的研究,而这项研究的关键技术就是建立生防菌的高效遗传转化体系。丝状真菌高效遗传转化体系的构建一般通过农杆菌介导或者聚乙二醇介导的原生质体转化来实现[12-17]。原生质体转化具有转化效率高、操作简单等特点,因此原生质体的制备是遗传转化的首要问题。
近年来,许多学者对丝状真菌遗传转化进行了大量的研究,但是由于真菌细胞壁组成和结构十分复杂,制备原生质体的体系和条件存在很大的差别。交枝顶孢A.implicatum对根结线虫具有良好的生防作用[18-19],考虑到其在线虫防治中良好的应用前景,对该菌进行原生质体制备与再生的研究,可为A.implicatum遗传体系的建立以及该菌生防机理的研究奠定良好的基础。
1 材料与方法
1.1 菌株
交枝顶孢A.implicatum菌株由中国农业科学院蔬菜花卉研究所蔬菜病害室保存。
1.2 培养基和试剂
马铃薯汤汁培养基(马铃薯200 g,D-葡萄糖20 g,定容至1 L;PDB)用于菌株产孢。马铃薯固体培养基(马铃薯汤汁培养基,1.5%琼脂;PDA)用于制备新鲜菌丝。固体再生培养基(0.1%酵母提取物,01%酶水解干酪素,1 mol/L蔗糖, 0.7%琼脂粉;SR),液体再生培养基(0.1%酵母提取物,0.1%酶水解干酪素,1 mol/L蔗糖; LR)。
原生质体制备所需试剂:PTC溶液(60%聚乙二醇3350,10 mmol/L Tris-Cl pH 7.5,50 mmol/L氯化钙),STC溶液(1.2 mol/L山梨醇,50 mmol/L氯化钙,10 mmol/L Tris-HCl)。
1.3 交枝顶孢原生质体的制备
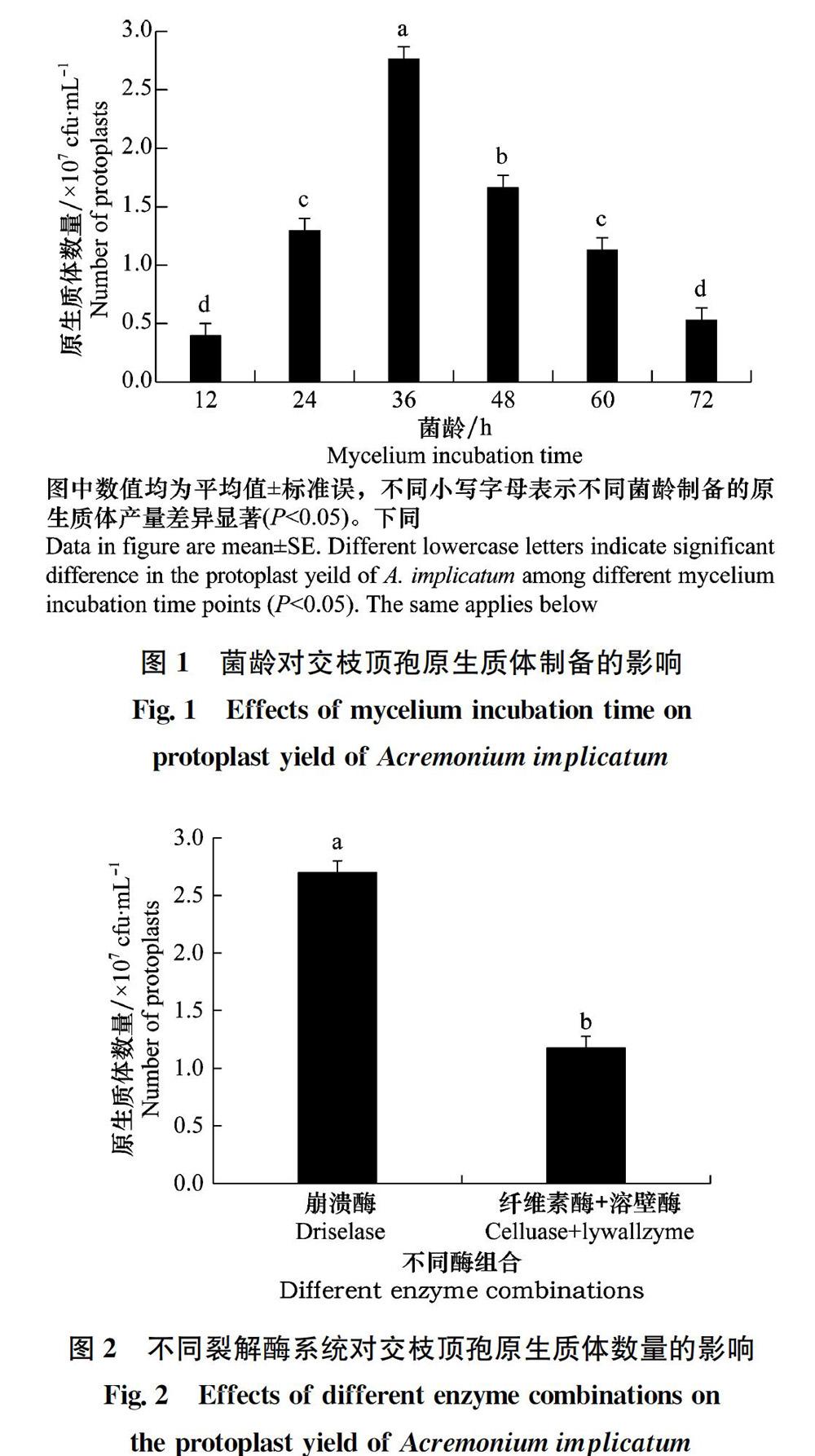
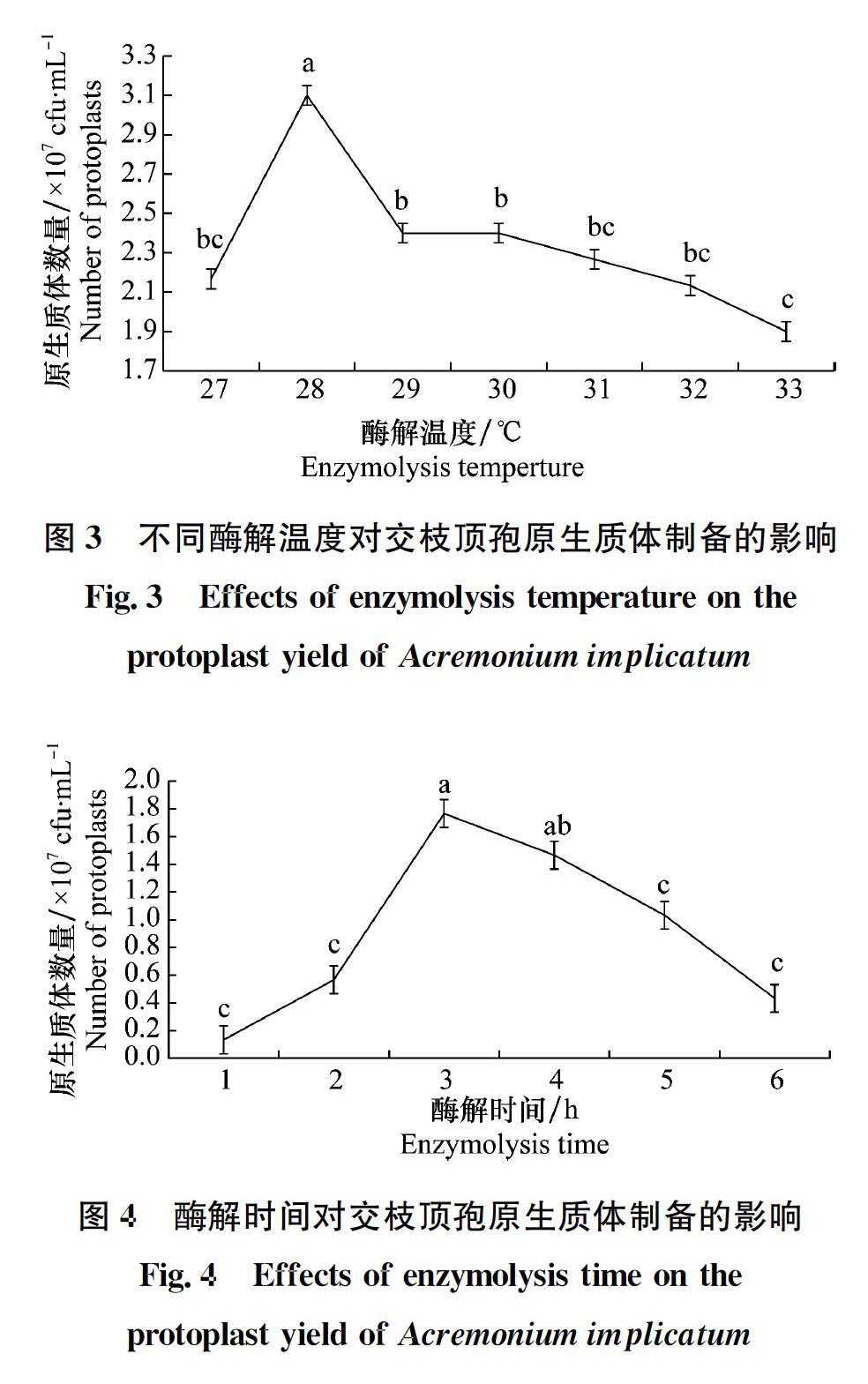
本试验采用酶解细胞壁法制备生防真菌交枝顶孢的原生质体。设置菌丝培养时间、裂解酶系统、酶解时间和温度、不同pH等试验参数,建立该菌株原生质体制备和再生体系。
1.3.1 菌丝培养时间对原生质体制备的影响
参照Parsons等[20]的方法制备原生质体并略有改动。交枝顶孢菌株在PDA平板上28℃黑暗培养5 d,用5 mm打孔器在菌落边缘打孔,取一菌块置于PDB中25℃,150 r/min 培养2 d,收集孢子并制备成均匀的孢子悬浮液(1×108个/mL)。取1 mL孢子悬浮液置于100 mL PDB中25℃, 150 r/min分别培养24、36、48、60、72 h,用灭菌过滤网过滤,然后用渗透压稳定剂反复充分冲洗收集菌丝,待菌丝冲洗至团状后转至灭菌的50 mL离心管中(离心管需提前称重),整个操作过程在超净工作台进行。用离心管称取收集好的菌丝0.5 g,加入2 mL酶解液(20 mg/mL崩溃酶)后置于摇床上振荡孵育,3 h后观察原生质体获得率,选出最适宜制备交枝顶孢原生质体的菌龄。试验设置3次重复。
為了检测制备的原生质体能否用于转化,将构建好的含有GFP基因的表达载体pCH-sGFP(中国农业科学院蔬菜花卉研究所提供)通过聚乙二醇介导的方法进行遗传转化。在蓝色激发光下,观察到转化子发出明亮的绿色荧光(图8),说明GFP基因已经成功转入交枝顶孢A.implicatum中,且能够正常表达。
3 结论与讨论
原生质体的制备和再生是建立真菌遗传转化体系过程中的关键技术,直接影响遗传转化效率。本研究采用单因素试验研究了影响内生真菌交枝顶孢原生质体制备和再生的条件,明确了交枝顶孢原生质体制备及再生的最适宜条件为:孢子液摇培36 h,用灭菌的过滤网过滤后,以pH为6.5的0.7 mol/L NaCl为渗透压稳定剂反复洗涤菌丝,用浓度为20 mg/mL的崩溃酶按照0.5 g菌丝:2 mL酶液在28℃恒温(120 r/min)酶解3 h,以SR培养基为再生培养基。
研究发现制备原生质体时菌丝的生理状态对原生质体产量和质量有着重要的影响。通常在菌丝线性生长早期原生质体容易制备,但也并不是菌丝越幼嫩越容易酶解。菌丝在其生长的某个阶段对降解酶最为敏感,可能是由于菌丝不同生长阶段细胞壁的结构组成不同,使得其对降解酶的敏感性不同[24-25]。
不同种类真菌细胞壁组成和结构不同,所用的裂解酶和酶解时间差异很大。弭宝彬等发现15 mg/mL崩溃酶对尖孢镰刀菌辣椒专化型Fusarium oxysporum f. sp. capsicum酶解效率最高[26]。赵小强等对制备大丽轮枝菌Verticillium dahliae原生质体的条件进行优化时发现,裂解酶浓度为10 mg/mL时原生质体产量最高[27]。因此,细胞壁降解酶是原生质体制备的另一个关键因素。试验采用了两种降解酶系统,试验结果表明对于交枝顶孢而言,20 mg/mL的崩溃酶酶解效果优于纤维素酶和溶壁酶混合酶的酶解效果。
在交枝顶孢原生质体制备的过程中,原生质体的数量随着酶解时间的延长呈现先增加后减少的趋势,在3 h时达到峰值,可能的原因为酶解时间长导致酶液活性降低或原生质体破裂。时涛等在橡胶多主棒孢的原生质体制备以及李伶俐等在甘蓝枯萎病菌原生质体制备研究中也有类似发现[28-29]。
本研究建立了食线虫真菌交枝顶孢原生质体的制备和再生体系,同时探明了体系相关的影响因素,为进一步研究交枝顶孢对根结线虫的生防机理提供了重要的材料基础。
参考文献
[1] ABAD P, GOUZY J, AURY J M, et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita [J]. Nature Biotechnology, 2008, 26(8): 909-915.
[2] TRUDGILL D L, BLOK V C. Apomictic, polyphagous root-knot nematodes: exceptionally successful and damaging biotrophic root pathogens [J]. Annual Review of Phytopathology,2001,39(1):53-77.
[3] 刘维志. 植物病原线虫学[M]. 北京:中国农业出版社,2000.
[4] NOLING J, BECKER J O. The challenge of research and extension to define and implement alternatives to methyl bromide[J].Journal of Nematology, 1994,26(4S): 573.
[5] 董锦艳,张克勤,赵智娴,等.杀线虫菌物毒素的研究进展(Ⅱ)[J].中国生物防治,2001,17(3):138-141.
[6] AMARAL D R, OLIVEIRA F E R, OLIVEIRA D F, et al. Purification of a Fusarium moniliforme metabolite toxic to Meloidogyne exigua [J]. Summa Phytopathologica (Brazil), 2003,29(1):25-29.
[7] SHARON E, BAR-EYAL M, CHET I, et al. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum [J]. Phytopathology, 2001,91(7): 687-693.
[8] GOSWAMI J, PANDEY R K, TEWARI J P, et al. Management of root knot nematode on tomato through application of fungal antagonists, Acremonium strictum and Trichoderma harzianum [J]. Journal of Environmental Science and Health Part B, 2008, 43(3):237-240.
[9] KIEWNICK S, SIKORA R A. Biological control of the root-knot nematode Meloidogyne incognita by Paecilomyces lilacinusstrain 251 [J]. Biological Control, 2006, 38(2): 179-187.
[10]YAN Xiaoning, SIKORA R A, ZHENG Jingwu. Potential use of cucumber (Cucumis sativus L.) endophytic fungi as seed
treatment agents against root-knot nematode Meloidogyne incognita [J]. Journal of Zhejiang University Science B, 2011, 12(3): 219-225.
[11]林森,武侠,曹君正, 等.产生几丁质酶的交枝顶孢(Acremonium implicatum)对南方根结线虫生防潜力[J].植物病理学报,2013,43(5):509-517.
[12]赵培宝,周庆新,郭芳先, 等.限制性内切酶介导的串珠镰刀菌插入突变和致病性突变体的分离[J].植物病理学报,2007,37(5):545-548.
[13]许梦秋,姜杰,孙漫红,等.限制性内切酶介导的粉红粘帚霉67-1转化体系构建[J].中国生物防治学报,2013,29(2):263-269.
[14]沈慧敏,李超,高利,等.原生质体法介导真菌遗传转化的研究进展[J].植物保护,2017,43(2):25-28.
[15]范亮波,李梅,冀颖,等.根癌农杆菌介导的木霉遗传转化及应用进展[J].生物技术通报,2010(3):1-5.
[16]胡彩平,郑金龙,高建明,等.根癌农杆菌介导柱花草炭疽菌遗传转化体系的优化[J].热带作物学报,2013,34(6):1007-1012.
[17]JIANG Dewei, ZHU Wei, WANG Yunchuan,et al. Molecular tools for functional genomics in filamentous fungi: Recent advances and new strategies [J]. Biotechnology Advances, 2013,31(8):1562-1574.
[18]TIAN Xueliang, YAO Yurong, CHEN Guohua, et al. Suppression of Meloidogyne incognita by the endophytic fungus Acremonium implicatum from tomato root galls [J]. International Journal of Pest Management, 2014, 60(4): 239-245.
[19]YAO Yurong, LIN Runmao, TIAN Xueliang, et al. The complete mitochondrial genome of the nematophagous fungus Acremonium implicatum [J]. Mitochondrial DNA Part A, DNA Mapping, Sequencing and Analysis, 2016, 27(5): 3246-3247.
[20]PARSONS K A, CHUMLEY F G, VALENT B. Genetic transformation of the fungal pathogen responsible for rice blast disease [J]. Proceedings of the National Academy of Sciences, 1987, 84(12):4161-4165.
[21]張卉, 刘长江. 姬松茸原生质体形成和再生的研究[J]. 微生物学报, 2003,23(3):18-22.
[22]李晶莹, 孙婷婷, 张国权,等.大型真菌原生质体的制备与再生研究[J]. 中国林副特产, 2015(4):98-101.
[23]REHMAN L, SU Xiaofeng, GUO Huiming, et al. Protoplast transformation as a potential platform for exploring gene function in Verticillium dahliae [J/OL]. BMC Biotechnology, 2016, 16(1): 57. DOI:10.1186/s12896-016-0287-4.
[24]孙传宝,朱春宝,许文思.产黄青霉原生质体制备和再生影响因子分析[J].中国抗生素杂志,2001,26(4):241-243.
[25]宋庆涛,张国珍,董金皋.玉米大斑病菌原生质体的制备与再生[J].微生物学通报,2003,30(2):41-44.
[26]弭宝彬,张吉祥,杨宇,等.尖孢镰刀菌辣椒专化型原生质体制备条件优化[J].生物技术通报,2013,4(4):96-100.
[27]赵小强,陈志荣,何芳,等.大丽轮枝菌原生质体的制备与再生[J].生物技术通报,2018,34(7):166-173.
[28]时涛,裴月令,刘先宝,等.橡胶树多主棒孢病菌原生质体转化体系的建立[J]. 热带作物学报,2009,30(9):1351-1358.
[29]李伶俐,严红,李兴红,等.甘蓝枯萎病菌原生质体的制备与再生条件的优化[J]. 中国农学通报,2011,27(10):203-207.
(责任编辑:杨明丽)
