甘露糖基糖蛋白N-乙酰氨基葡萄糖转移酶在恶性肿瘤中作用的研究进展
2020-12-24闫文武汪进国
杜 俊,闫文武,汪进国
(1.皖南医学院;2.皖南医学院附属弋矶山医院胃肠外科,安徽 芜湖 241000)
N-乙酰氨基葡萄糖转移酶是催化UDP-GlcNAc连接到N-连接型聚糖或者O-连接型聚糖上的一类酶。其中以UDP-GlcNAc为糖基供体,并将其连接到五糖核心甘露糖残基上的GnTs目前最少有七种,分别为GnT I、GnT II、GnT III、GnT IV、GnT V、GnT VI和GnT IX。N-聚糖的五糖核心是由2个N-乙酰氨基葡萄糖和3个甘露糖(mannose,Man)构成,其中每个Man残基上可以至少形成一条分支糖链(又称天线)。GnTs属于II型跨膜糖蛋白且对糖基接受体比较专一。GnTs对底物的催化有严格的顺序,必须由GnT I和GnT II形成二天线N-聚糖后才能形成三、四、五天线N-聚糖,而GnT III可以抑制N-聚糖分支天线的形成(图1)。
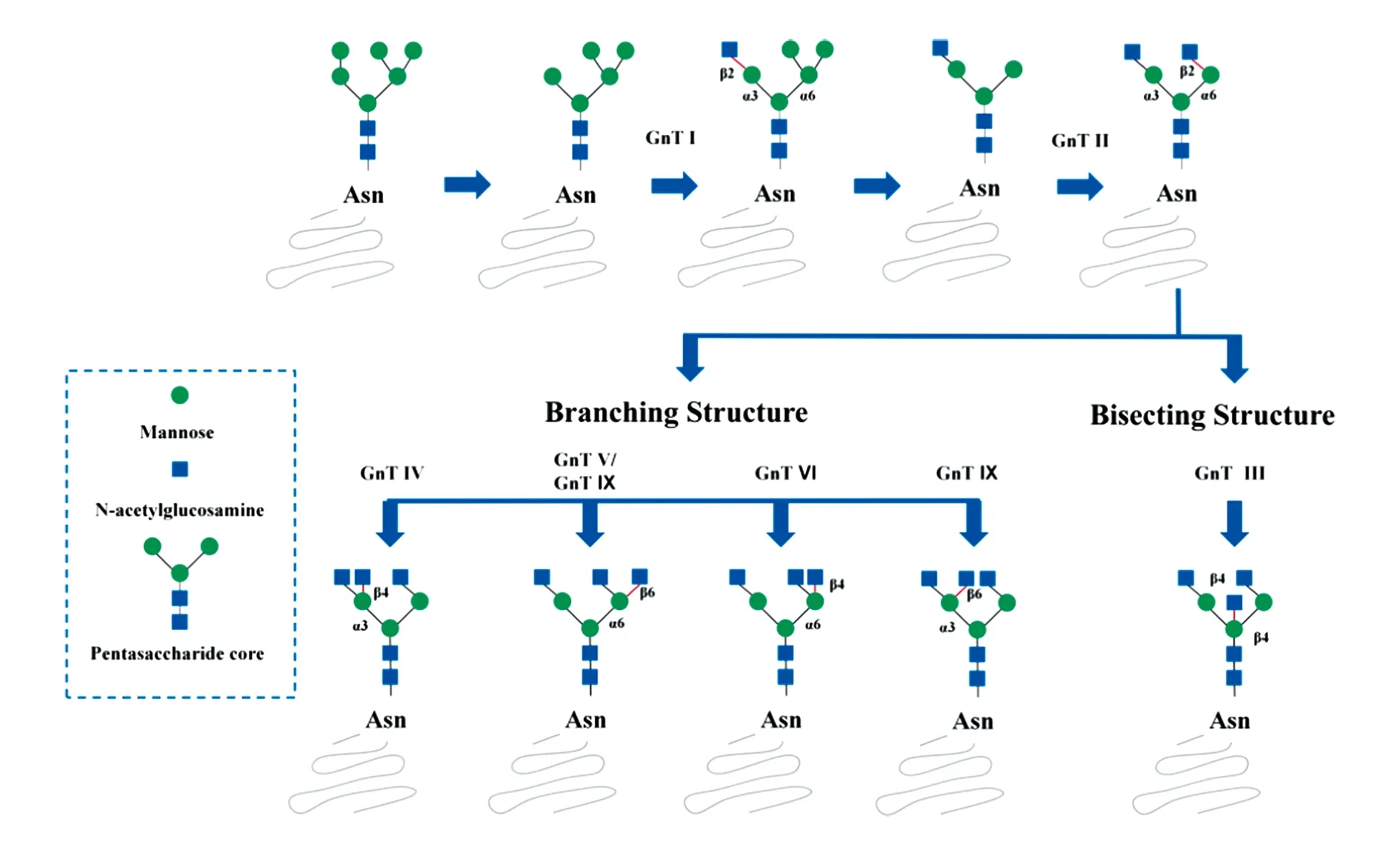
图1 N-聚糖的分支和平分结构及其相关N-乙酰氨基葡萄糖转移酶
1 GnT I与恶性肿瘤
α-1,3-甘露糖基糖蛋白β-1,2-N-乙酰氨基葡萄糖转移酶(GnT I)催化UDP-GlcNAc连接到五糖核心中α-1,3甘露糖残基末端,并且是启动合成复杂型和杂合型N-连接型聚糖的关键酶。通过siRNA转染技术和CRISPR/Cas 9基因编辑技术降低Hela细胞中MGAT1的表达,可以减少细胞N-聚糖的分支结构并且抑制肿瘤细胞的恶性表型[1]。MGAT1在胶质瘤中高表达,EGFR信号通路既可以激活MGAT1和GLUT1蛋白的表达,同时MGAT1也可以调节GLUT1蛋白表达进而部分地促进胶质瘤细胞的增殖和迁移[2]。Wnt/ β-catenin信号通路激活也可以增加MGAT1蛋白表达,并且构建稳定表达MGAT1的Huh7细胞株在体内外实验验证了其促进肝癌细胞的作用[3]。与之相反的是有文献导报MGAT1在肝癌组织样本中低表达且与预后不良相关[4]。这可能与GnT I的表达减少导致高甘露糖型N-聚糖增多有关。
2 GnT II与恶性肿瘤
α-1,6-甘露糖基糖蛋白β-1,2-N-乙酰氨基葡萄糖转移酶(GnT II)是MGAT2基因编码由447个氨基酸组成的II型跨膜蛋白,在高尔基体中发挥作用。GnT II催化UDP-GlcNAc连接到五糖核心中α-1,6甘露糖残基末端。GnT I和GnT II按先后次序催化形成二天线N-聚糖后再由其他酶催化形成多分支N-聚糖。MGAT2基因突变会导致先天性糖基化障碍综合征IIa型[5],但目前为止尚未报道GnT II在恶性肿瘤作用机制。
3 GnT III与恶性肿瘤
β-1,4-甘露糖基糖蛋白β-1,4-N-乙酰氨基葡萄糖转移酶(GnT III)在高尔基体中催化UDP-GlcNAc连接到五糖核心中α-1,4甘露糖残基末端,形成二分型/平分型GlcNAc N-聚糖。含有二分结构的N-聚糖可以抑制GnT IV、GnT V和GnT IX继续催化N-聚糖形成分支结构,所以GnT III可能起到调节平衡N-聚糖糖链分支的作用。上皮-间充质细胞转化(epithelial-mesenchymal transition,EMT)是肿瘤浸润和转移的一个重要的机制。EMT可以被多种细胞信号通路诱导,并且GnT III可能参与其中。通过人表皮生长因子诱导GE11上皮细胞EMT过程中,GnT III被特异性的下调并且凝集素鉴定结果显示二分结构N-聚糖明显下降。将MGAT3稳定转染GE11细胞后,E-cadherin表达增多且纤维连接蛋白表达减少[6]。转化生长因子β1(TGF-β1)诱导肝癌细胞EMT模型中Smad3和Erk信号通路被激活从而抑制GnT III mRNA及平分型GlcNAc结构形成[7]。TGF-β1也可以通过激活JNK/P38/PIK3信号通路抑制非肌肉肌球蛋白II A(NMIIA)的表达。NMIIA表达下调可以使核心2 β-1,6-N-乙酰氨基葡萄糖转移酶(C2GnT-M)的表达增加,并且C2GnT-M抑制GnT III表达从而促进肺癌细胞的EMT和转移[8]。GnT III的表达可以显著抑制β4整合素上的β1,6-GlcANc分支结构从而抑制细胞迁移和肿瘤形成[9]。相反的是,有文献报道GnT III的表达与乳腺导管内原位癌亚型粉刺癌及病理分型高核级有关,并提示不良预后[10]。
GnT III仅在不含或β-2,6唾液酸化含量较低的肿瘤细胞中才起到肿瘤抵抗迁移作用。因为GnT III仅可以抑制β-2,3唾液酸化但不可以抑制β-2,6唾液酸化的能力,并且当ST6GAL1过表达时反而可以抑制GnT III在肿瘤细胞中抗迁移作用。虽然GnT III催化N-聚糖形成的二分结构可以抑制GnT IV、GnT V、GnT IX从而作为抑癌因子,但这种抑制似乎是有限的,可能受到其他糖基转移酶的影响。所以GnT III与其他糖基转移酶在的相互作用影响肿瘤进展机制仍需阐明。
4 GnTIV与恶性肿瘤
α-1,3-甘露糖基糖蛋白β-1,4-N-乙酰氨基葡萄糖转移酶(GnT IV)有三个同工酶分别是:GnT IVa(MGAT4A)、GnT IVb(MGAT4B)和hGnT IV-H(MGAT4C)。其中,GnT IVa和GnT IVb具有相似的催化活性,分布于高尔基体中。GnT IVa和GnT IVb在不同组织分布表达也各不相同。在胰腺癌组织中以GnT IVb的表达为主,而癌旁组织中则以GnT IVa的表达为主。跨膜蛋白细胞粘附分子(CAMs)在细胞之间相互黏附中起重要作用。CAMs大部分都是糖蛋白,它们与胞外配体结合后可以改变尾部结构,经过一系列反应后激活多种信号通路,从而影响细胞增殖、迁移、分化和凋亡。GnT IV可以催化胞外糖蛋白形成糖链分支结构,进而增加肿瘤细胞的恶性行为。CD147是免疫球蛋白超家族成员之一并且可诱导分泌基质金属蛋白酶促进肿瘤的迁移。GnT IVa的表达可以改变细胞膜上CD147糖链分支影响小鼠肝癌细胞的转移能力。整合素存在于细胞膜外层,调节细胞和细胞基质之间的粘附作用。抑制GnT IVa的表达可以减少整合素β1上β-1,4 GlcNAc的分支最终降低绒癌的侵袭性。Nishino等人通过使用特异性识别由GnT IVa催化的N-聚糖糖链分支的凝集素进行免疫组织化学染色,结果显示在侵袭性葡萄胎和子宫内膜癌滋养细胞中存在大量与其相关的N-聚糖分支结构。GnT IVa催化形成高度分支化的溶酶体相关膜蛋白2并在细胞膜表面表达,这影响了细胞与细胞外基质(extracellular matrix,ECM)之间的黏附性从而促进滋养细胞肿瘤迁移和侵袭[11]。GnT IVa可以调节细胞表面蛋白核心α-1,6 岩藻基三天线聚糖的表达从而影响肝癌细胞的转移。
5 GnT V与恶性肿瘤
α-1,6-甘露糖基糖蛋白β-1,6-N-乙酰氨基葡萄糖转移酶(GnT V)在高尔基体中催化N-聚糖合成β-1,6分支。GnT V表达与肝癌和肾透明细胞癌不良预后有关,并且在肿瘤凋亡,肿瘤增殖、肿瘤免疫、肿瘤耐药和肿瘤转移起到至关重要的作用。上皮细胞生存必须依赖ECM的粘附,一旦脱离ECM就会失巢凋亡。MGAT5可以增加结肠癌细胞的锚定依赖性和锚定非依赖性生存能力,使结肠癌细胞在脱离ECM时抵抗失巢凋亡[12]。同时GnT V也可以调节细胞周期,抑制GnT V表达可以使U87细胞停滞在G0/G1期,而提高GnT V表达可以诱导G0/G1期向G2/M期转变。在肿瘤免疫方面,GnT V过表达转基因小鼠巨噬细胞表面受体的糖链发生变化从而抑制巨噬细胞免疫应答,并且白细胞介素-10的表达显著降低可以导致结肠炎的加重,这些表型的改变增加结肠肿瘤的发生几率。放射治疗是肿瘤治疗方法之一,对于某些肿瘤具有良好的治疗效果。但是患者在接受放射治疗过程中可能会出现放疗抵抗的情况,GnT V可能通过EMT参与肿瘤细胞的放疗敏感性。使用RNA干扰技术将鼻咽癌细胞、小细胞肺癌细胞和胶质瘤细胞中GnT V敲除后可以明显增强肿瘤细胞对放射的敏感性,所以GnT V是治疗放射抵抗的重要靶点。化学药物治疗是肿瘤治疗最有效的方法,其中吉西他滨是治疗膀胱癌的一线药物。吉西他滨耐药是导致膀胱癌患者死亡的重要因素,GnT V可以使人平衡核苷转运体1上的糖链分支结构改变,提高其转运活性继而增加膀胱癌细胞对吉西他滨的敏感性[13]。
肿瘤转移是肿瘤患者致死的最主要原因,GnT V在细胞黏附和EMT也起到重要作用。GnT V增加口腔癌细胞癌胚抗原相关细胞黏附分子6上糖基化位点N256的糖链分支激活EGFR信号通路进而促进细胞的侵袭和转移[14]。GnT V降低μ型受体蛋白酪氨酸磷酸酶的酪氨酸磷酸酶活性然后通过PLCγ-PKC信号通路促进胶质瘤细胞转移[15]。PIK3/Akt信号通路可以激活肝癌细胞中GnT V的表达,同时在EMT过程中GnT V表达促进CD147与整合素β1相互作用激活PIK3/Akt信号通路增强细胞侵袭和迁移的潜能[16]。有研究发现miR124-3p可直接靶向GnT V从而抑制乳腺癌细胞的生长[17]。综上所述,GnT V是肿瘤治疗的有效靶点。
6 GnTVI与恶性肿瘤
α-1,3-甘露糖基糖蛋白β-1,4-N-乙酰氨基葡萄糖转移酶同工酶C(hGnT IV-H/ GnT VI)催化UDP-GlcNAc转移到五糖核心中α-1,6甘露糖残基末端形成β-1,4 GlcNAc糖链分支。存在最多分支结构的N-聚糖带是二分结构的五天线N-聚糖,它仅存在于脊椎动物中。其次是五天线N-聚糖,目前在母鸡输卵管中发现,但尚未在哺乳动物中报道。有研究发现GnT VI的一级结构与GnT IV的同工酶hGnT IV-H(MGAT4C)的一级结构非常相似[18]。使用慢病毒干扰技术降低MGAT4C的表达,HEK293细胞表面的CD133表达及其AC133表位糖链分支结构显著降低。RWPE1和VCaP细胞株过表达MGAT4C后可以促进前列腺良恶性肿瘤细胞的增殖和迁移。以上结果表明GnT IV-H可能在肿瘤进展中起到促进作用。虽然如此,MGAT4C并未直接表现出其含有GnT IV的活性,所以MGAT4C是否含有N-乙酰氨基葡萄糖糖基转移酶的功能有待验证。
7 GnTIX与恶性肿瘤
α-1,6-甘露糖基糖蛋白β-1,6-N-乙酰氨基葡萄糖转移酶同工酶B(GnT Vb/ GnT IX)是GnT V相关同系物,它们之间具有42%的同源相似度。与GnT V相比它能催化更多的底物,GnT IX既可以催化UDP-GlcNAc转移到五糖核心中α-1,3或者α-1,6连接型甘露糖残基末端,也可以催化O-甘露聚糖形成β-1,6 GlcNAc糖链分支结构。GnT IX主要在脑组织和睾丸组织中表达,而在其他组织肿瘤中的功能尚不清楚。启动子甲基化异常可以影响基因的表达,通过分析肝癌、乳腺癌、宫颈癌和黑色素瘤中甲基化差异发现,MGAT5B在这几种癌症中均出现启动子异常甲基化[19]。经菜豆白细胞凝集素免疫组化首次发现前列腺癌细胞比正常前列腺上皮细胞的GnT IX表达明显提高。Gao等人使用凝集素亲和法比较正常人和肝癌患者血清糖蛋白表达差异,结果显示GnT IX在肝癌患者血清中的明显上调[20]。虽然这些结果表明MGAT5B可能属于促癌基因,但其在肿瘤中的分子作用机制仍需进一步研究。
8 总结与展望
GnTs家族通过催化N-聚糖形成分支结构或平分结构从而调节各种恶性肿瘤增殖和转移等恶性行为(表1)。目前,GnT III和GnT V已在多种肿瘤中被验证前者属于抑癌因子后者属于促癌因子。然而其他的GnTs家族成员在肿瘤中的详细的作用机制仍不清楚。在现代分子生物学发展过程中,糖组学越来越受到人们的关注。糖链不像是DNA合成需要模板一样,而是通过糖基转移酶催化形成构象不一的糖链,这样就给糖蛋白的研究带来很多困难。但是,随着实验技术的不断成熟,科研方法不断完善,新的且有催化活性的GnTs家族成员及其作用机制可能不断被发现。因此深入研究GnTs在肿瘤中作用靶点、参与肿瘤的信号通路以及与其他糖基转移酶之间的相互作用,为靶向治疗癌症提供新思路。
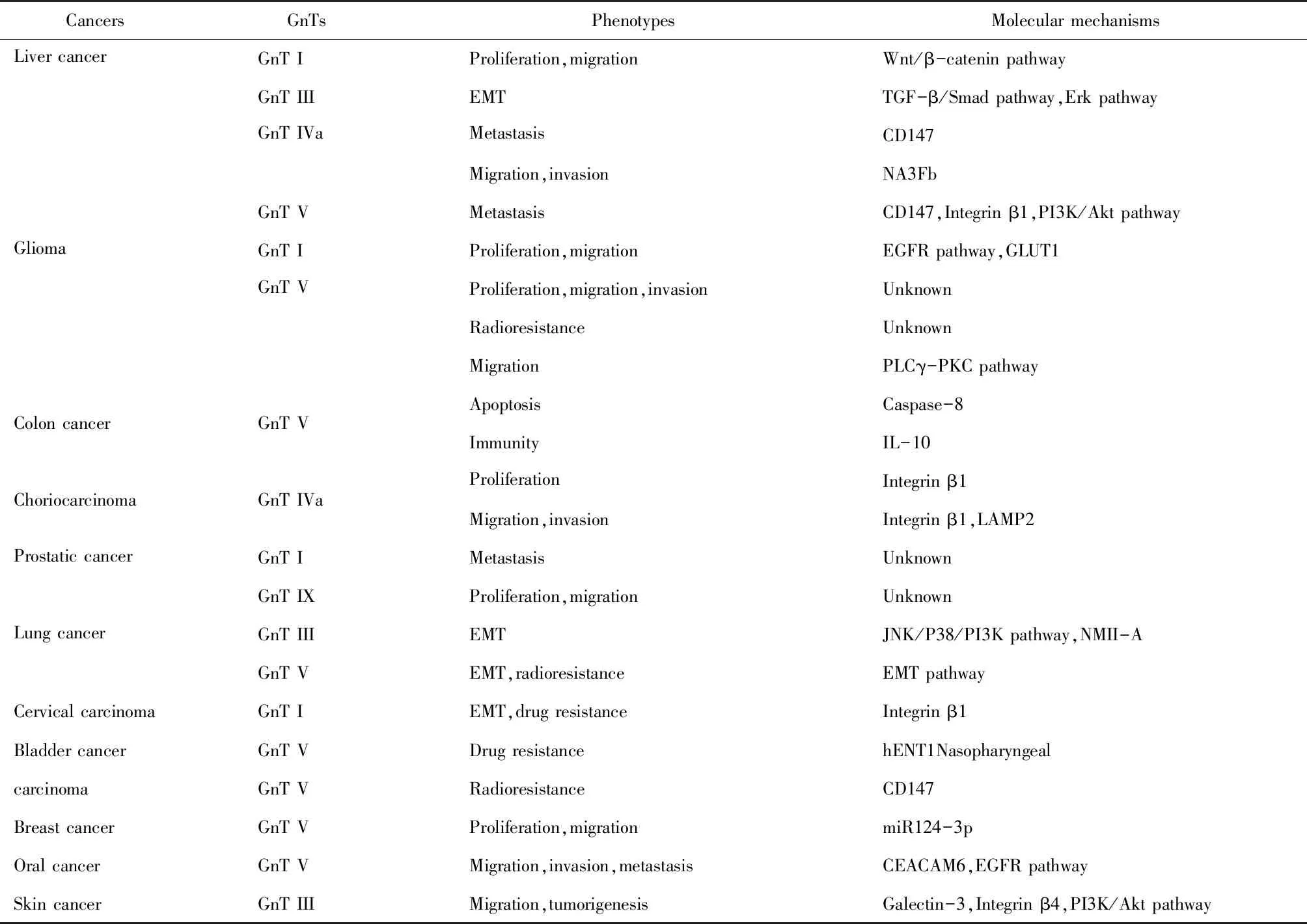
表1 GnTs相关恶性肿瘤的表型和分子机制
