渭河阶地不同生境反枝苋种子萌发的热量需求研究
2020-12-14王雅馨魏永胜刘嘉聪
王雅馨,杨 婷,魏永胜,刘嘉聪
(1.西北农林科技大学 生命科学学院,陕西杨凌 712100;2.西北农林科技大学 植物保护学院,陕西杨凌 712100;3.西北农林科技大学 机械与电子工程学院,陕西杨凌 712100)
入侵杂草危害是全球重大生态问题之一[1]。反枝苋(AmaranthusretroflexusL.)为苋科、苋属1a生草本植物,是一种典型的C4植物,具有高光合效率、低CO2补偿点的光合特性[2],以及生长快、籽实多等生物学特性,一株成株的反枝苋种子产量高达1万~3万粒[3]。同时,反枝苋具有较强的表型可塑性[4],可通过增加自身的生态幅和耐受力来适应环境因子频繁变化的生境,如反枝苋在昼夜温度为23℃/13℃和20℃/10℃条件下单株结实量分别为5465粒和1655粒,单株干质量分别为2.1g和0.7g[5]。反枝苋是世界广布性入侵杂草[6],也是中国目前苋属植物(共9种)中入侵数量居首位和分布最为广泛的一种[7],对农田的危害面积在中国暖温带高达36%[8]。反枝苋可在棉花、玉米[9]、小麦[10]、烟草、大豆[11]等多种作物的田地生长,通过化感作用、与作物竞争光照[12]、水分[13]和营养物质[14]以及作为病虫害的传播介体等,造成作物大量减产[15],导致严重的经济损失。例如,棉花田中每平方米增加1株反枝苋可导致棉花单株铃数减少20.81%~84.98%[16]。此外,反枝苋对家畜健康[17]、人类健康[18]以及生态环境都有一定的危害。
全球气候变暖条件下,作为C4植物的反枝苋的生态适应情况鲜有报道。种子萌发阶段是反枝苋成功入侵多类型生境的关键阶段,苗期则是实施防控的关键时期[19]。种子萌发所需的时热累积模型参数具有一定的生理和生态意义,能够有效预测不同温度条件下的种子萌发时间及种苗出土时间[20]。但目前关于反枝苋萌发的热量需求尚不清楚,无法准确预测其在田间的萌发日期。因此,通过探究不同生境下反枝苋种子的萌发热量需求,探讨反枝苋对全球气候变暖的响应,并预测其萌发日期,对进一步掌握反枝苋的生物学特性,优化反枝苋的防控方法,以提高防效有重要意义。为此,本研究拟利用积温模型(thermal time model),定量估计不同生境下反枝苋种子的萌发热量需求,预测反枝苋对全球气候变化的响应及萌发日期,进而为反枝苋的适时防除提供理论 依据。
1 材料与方法
1.1 材 料
反枝苋种子于2019年9月采自陕西省咸阳市杨凌区渭河阶地(图1),该区位于西北农林科技大学北校区(北纬 34°15′,东经 108°03′,海拔:519 m,即头道塬,以N表示)、南校区(北纬 34°17′,东经 108°04′E,海拔:459 m,即二道塬,以S表示)以及渭河河岸旁(北纬34°13′,东经 108°07′,海拔:438 m,即河滩地,以R表示),每个地随机取3个样点(间距>300 m),每样点各3株。种子去杂、自然风干后保存于牛皮纸信封,室温通风干燥处储藏,待用。

R.河滩地;S.二道塬;N.头道塬;下同
1.2 方 法
1.2.1 种子质量评价 种子千粒质量与含水量的测定:随机选取200粒种子进行测量,重复3次(共600粒),称量后计算千粒质量。另取3份各100粒种子,称量后放入称量瓶,置于80 ℃烘箱烘24 h后取出,于干燥器内冷却30 min至室温,称取干质量后计算含水量。
种子活力评估:活力测定用TTC染色法[21],轻压种子使种皮破裂,置于培养皿中,加入w=0.5%TTC溶液10 mL,置35 ℃黑暗条件下4 h,之后检查种子的胚,统计染色结果。胚有光泽的鲜红色且组织状态正常的为有活力的种子。并计算活力,具体为:V=n/N×100%,式中V为种子活力,n为染色种子数,N为供试种子总数。试验重复3次,每次50粒种子。
1.2.2 萌发试验 取3个生境的种子各50粒,用φ=75%的酒精消毒0.5 h,蒸馏水冲洗3次,置于铺有双层滤纸、直径为9 cm的培养皿中,加5 mL蒸馏水后加盖分别置于10、20、25、30、35和40 ℃的恒温培养箱(上海福玛,SPX-250B)内进行萌发试验。试验重复5次。每个培养箱中置1支UT330ABC温度计,采样间隔0.5 h,测得实际温度分别为10.2、19.2、24.7、28.9、36.5和39.6 ℃。自吸水后计时,以种子胚根突破种皮1 mm为萌发,每间隔2 h观察并记录萌发率。计数后的种子取出,至连续1周无种子萌发时结束试验。最终计数未萌发的种子,并评估种子萌发率(G)。G=ni/(N×V) ×100%,ni为i时刻萌发种子数,N为供试种子总数,V为活力。
1.3 数据分析
1.3.1 萌发过程的拟合 利用sigmaplot 14.0以经过活力校正的累积萌发率(y)对种子吸水时间(x)做散点图,利用Chapman 3 Parameter方程y=a(1-e-bx)c拟合,估计参数a、b、c。其中a为估计的最高萌发率,有生物学意义,因此本模型为半理论模型。
利用上述方程分别计算,当累积萌发率y=20%、30%、…70%时,所需萌发时间x的值,以1/x(萌发速率)对温度作图,最适温度之前的各萌发率线性回归方程的斜率为相应的1/θT(g)。θT(g)为调整累积萌发率达到g%(相应的萌发率,即y)所需要的积温,所有θT(g)呈正态分布[22]。
1.3.2 调整累积萌发率达到50%所需积温 θT(50)及积温标准差σθT的评估 利用正态分布函数进行评估:对应的种群相对频率呈正态分布,对其进行标准正态离差转换。以θT(g)为横坐标,达到不同萌发率的概率密度Probg为纵坐标做散点图,并进行线性回归方程拟合Probit(θT(g))=(θT(g)- θT(50))/ σ θT(g)。拟合曲线的截距为 [-θT(50)]/ σθT(g),斜率为1/σθT。故θT(50)为负的截距除以斜率,σθT为斜率的倒数。
1.3.3 萌发时间预测 气象资料由西北农林科技大学旱区农业研究院提供。利用公式y=y0+asin(2πx)/b+c(式y0为年平均温度,x为儒略日,a、b和c为待估参数)对杨凌地区2008-2018年的月平均温度的散点图进行回归拟合,估计参数y0,其中y0为年平均温度。以历年拟合方程中的y0(年均温度)对年份进行线性回归,预测2020的平均温度的y,进而估计萌发时间。
1.3.4 数据处理 数据用R软件进行方差分析及多重比较(Duncan’s法),结果以“平均值±标准误”的形式表示。用Sigmaplot 14.0软件制图。
2 结果与分析
2.1 反枝苋种子质量评估
3个不同来源的反枝苋种子均呈黑色圆形,直径1~2 mm,种皮较硬,表面光滑且有光泽。不同生境种子的千粒质量、含水量及活力均有显著差异(表1)。河滩地、二道塬和头道塬三地的千粒质量分别为0.348 5、0.311 7、0.322 5 g,含水量分别为30.29%、25.32%、24.38%,活力分别为87.61%、80.37%、87.83%。

表1 不同生境反枝苋种子质量Table 1 Quality of Amaranthus retroflexus seeds collected from three habitats
2.2 反枝苋萌发过程
3个不同来源的种子萌发过程存在明显差异(图2),在10.2 ℃时三地的种子萌发均极少。在19.2 ℃时,头道塬反枝苋种子萌发率即可达到40%以上,而其他两地的种子萌发不足5%。头道塬反枝苋种子萌发最适温度为28.9 ℃,低于二道塬和河滩地的最适温度36.5 ℃。头道塬、二道塬、河滩地相应的最高萌发率分别为70.83%、90.66%和34.68%。表明低海拔地区种子更喜高温。海拔越低,累积萌发曲线变化越缓,表明其萌发速率越低,种子萌发不整齐(图3)。
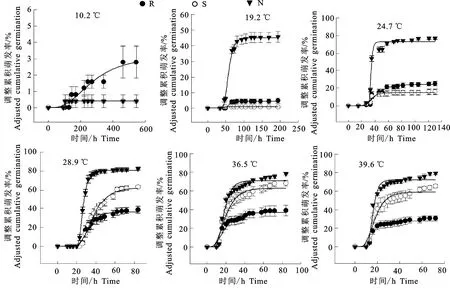
图2 反枝苋种子不同温度条件下种子的调整累积萌发率Fig.2 Adjusted cumulative germination of Amaranthus retroflexus seeds at different temperatures

图3 不同温度条件下头道塬和二道塬反枝苋种子的萌发速率直线Fig.3 Germination rate against constant temperatures for Amaranthus retroflexus seed collected from first and second tiered terraceland
2.3 反枝苋时热模型参数评估
由于本试验河滩地反枝苋种子的最高萌发率为34.68%,无法评估θT(50),故只讨论二道塬、头道塬两地种子萌发的θT(50)及标准误σθT。
头道塬和二道塬种子的Tb为13.98和 23.14 ℃;θT(50)为398.74和326.79 ℃·h,(由于标准差较大,因此二者差异达不到显著水平,表2)反枝苋调整累积萌发率随着积温的增大而升高(图 4)。头道塬种子萌发所需积温变幅窄,为334.04 ~463.44 ℃·h;而二道塬的较宽,为177.76~ 475.82 ℃·h。高海拔种源的种子萌发所需积温明显高于低海拔种源的种子(表2和图4)。头道塬种子的萌发积温标准差为64.70 ℃·h,不足二道塬(149.03 ℃·h)的一半(表2),表明头道塬种子萌发时间更为集中,自然条件下的适应性弱于二道塬的种子。
2.4 反枝苋种子萌发日期预测
对杨凌地区2008—2018年各月平均温度的散点图进行回归拟合,结果表明,杨凌多年平均温度(y0)分别为8.9、11.6、13.1、8.8、11.4、12.1、13.2、13.3、12.9、14.3和12.3 ℃,其对年份的线性回归方程为y=0.348 6x-689.714 4,R2= 0.424 3。预测2020年平均温度为14.5 ℃。可以得到2020年的日平均温度数据方程:y= 14.5+14.04sin(2πx/405.21+4.84),利用以上公式找到积温达到θT(50)及达到其95%的置信限时的日期。预测萌发日期以及最早、最晚萌发日期如表3所示。二道塬的反枝苋种子萌发率达50%的日期是6月6日,晚于头道塬的4月18日,但萌发时期变幅为15 d,而头道塬的则只用 4 d。因此,水热条件好时,反枝苋种子萌发时间区间更宽,防控更加困难。

表2 反枝苋种子萌发时热模型参数估计Table 2 Model parameters estimation of Amaranthus retroflexus seed germination

图4 头道塬和二道塬反枝苋种子萌发的积温模型曲线Fig.4 Thermal time diagram of Amaranthus retroflexus seeds collected from first and second tiered terraceland
3 讨 论
3.1 反枝苋萌发的热量需求
不同生境反枝苋种子萌发的基础温度、热量需求均存在较大差异(表2)。虽然头道塬、二道塬两地海拔相差只有60 m,但多年物候表明,二道塬植物开花早于头道塬1~2周。位于低海拔的二道塬反枝苋种子的萌发基础温度比头道塬的高9.16 ℃。表明在水热条件相对较好的条件下,反枝苋种子萌发启动虽然晚,但萌发周期长;而处于高海拔的反枝苋种子,萌发启动温度低,但萌发所需积温较高,种子不会因短暂适宜的温度而全部萌发,在温度积累到较高水平才萌发,这是一种防止高海拔地区温度条件对其幼苗不利、造成损害的萌发策略。高海拔的反枝苋种子的σθT小,表明高海拔来源的种子萌发积温范围窄,萌发相对整齐,会在短时间内萌发,为后期的营养生长和生殖生长留出更长的时间。

表3 积温模型预测二道塬和头道塬两地反枝苋萌发的日期Table 3 Prediction of germination date of Amaranthus retroflexus seeds based on thermal time model
通过对比几种经济作物萌发的基础温度与积温需求发现,反枝苋种子萌发对基础温度要求较高,而对积温要求较低。如大豆种子萌发的基础温度为3.33 ℃,积温需求为216.7 ℃·d;花生萌发的基础温度为7.5 ℃,积温需求为87.5 ℃·d[23]。可以看出,反枝苋萌发基础温度高于大豆和花生,所需积温远远小于大豆、花生等经济作物。相较于其他杂草,如野燕麦种子萌发的基础温度为-3.27 ℃,积温需求为2 146.70 ℃·h等,同样具有明显差异。对于萌发基础温度低、积温高的种子来说,种子萌发需要较长的时间,以保幼苗躲过不利的温度条件正常生长[24]。而反枝苋萌发基础温度较高,可确保幼苗于适宜的温度条件下生长,故对积温没有较高要求。
3.2 反枝苋对未来气候变暖的可能响应
反枝苋种子的萌发率随温度的升高增大,在10.2 ℃时几乎不萌发;19.2 ℃时高海拔种源的种子最先萌发,24.5 ℃以上时,3种来源的种子仅需40 h都可达到其最高萌发率(图2)。马鹏里等[25]对1961-2014年气候变暖背景下中国干旱变化的研究表明,中国西北地区干旱程度明显减轻,因此西北地区有向暖湿气候发展的趋势。在这一背景下,反枝苋达到种子萌发基础温度的日期将会提前[26],其萌发也将提前。有研究表明,萌发早的反枝苋生活史周期较长,春季物候期提前,秋季物候期延迟[26],生育期更长,有更充分的营养生长,因而种子的质量与数量将会更大,全球变暖将会增加其繁殖优势。此外,反枝苋作为一种典型C4植物,具有高光合效率、低CO2补偿点的光合特性,全球气候变暖将更有利于其生长,因此会对田间生产造成更大影响。本研究中,低海拔暖湿条件下的种子的σθT大,表明低海拔来源的种子萌发积温范围广,自然适应能力强,田间防除更加困难。因此,在全球气候变暖的条件,反枝苋的防控难度将增加。
3.3 反枝苋防控建议
反枝苋在适宜的条件下可达到90%的萌发率(图2),表明若不及时防除,将对田间生产造成极大危害。反枝苋对多种除草剂都产生了抗性[27],但幼苗期抗性较弱,施用除草剂可以达到较好的防治效果。因此,定量估计反枝苋的萌发日期,可以为适时化学除草提供可靠依据,减少用药,提高效率。由于正在萌发的种子被土壤覆盖,喷洒茎叶处理剂对其进行防除没有明显效果。该时期可采取翻土、施用苗前土壤处理除草剂,例如氯溴隆、利谷隆、溴谷隆等对其进行防治[28]。本研究中,反枝苋萌发后3 d,幼苗能长到15 mm左右,5 d长到25 mm左右并进入2叶期,此时使用450.0 g a.i./hm250%溴苯腈、350.0 g a.i./hm270%麦草畏或600.0 g a.i./hm240%甲基磺草酮/特丁津混剂可有效防治反枝苋[29]。若在4~5叶期防控,可考虑施用孢子浓度107个/mL的Alternariaalternate,持续6 h的湿度,导致叶片坏死,植株萎蔫死亡[28]。小根蒜、臭椿、角茴香根等多种植物的提取液都能够抑制反枝苋种子发芽,降低幼苗鲜质量,并且随着提取物浓度的提高,抑制效果更加明显。
4 结 论
反枝苋种子的萌发的基础温度和最适温度较高,萌发热量需求少,是典型喜温杂草,萌发周期短,在全球变暖的大背景下,其萌发日期将会提前,萌发期和生活史周期将会延长,其生长繁殖都会处于优势地位,防控难度增加。杨凌地区正常年份的萌发日期为4月16日至6月12日(因COVID-19疫情影响,原计划的野外调查无法实施),此后1~2周内实施化学除草效果最好。
