芥甘杂种后代F1和F2形态学和细胞遗传学研究
2020-12-10余青兰才吉巴毛赵志刚
赵 彤,谢 晋,殷 婷,余青兰,才吉巴毛,赵志刚
(青海大学农林科学院,青海 西宁 810016)
【研究意义】甘蓝型油菜(Brassicanapus.,AACC,2n=38)是白菜型油菜(B.rapa,AA,2n=20)和甘蓝(B.oleraces,CC,2n=18)进行天然种间杂交,最后通过染色体天然加倍形成的异源四倍体[1-4],其产量高,适应性广,抗逆性强,是新中国成立之后的主栽品种[5-6]。但由于甘蓝型油菜无天然的野生种和后期人工定向选择,造成遗传多样性日益匮乏,遗传基础越发狭窄[7-8]。利用不同类型的油菜进行种间杂交,选育优良后代,是发展油菜产业,培育优质甘蓝型油菜行之有效的方式。【前人研究进展】目前国内外关于油菜与不同种或属之间的材料进行远缘杂交已有许多成功的范例。Wen等[9]以白菜和黄籽甘蓝为亲本进行杂交,成功得到新材料黄籽甘蓝型油菜。王学芳等[10]成功通过白菜类蔬菜(B.campestris, 2n=2x=AA=18)的紫菜苔、小白菜和大白菜与甘蓝型油菜 (B.napus, 2n=4x=AACC=38)进行杂交,得到了新型早熟甘蓝型油菜新种质。Li等[11]用埃塞俄比亚芥与白菜型油菜杂交获得三倍体(ABC),该三倍体可作为芸薹属植物优良性状转移的桥梁。McNaughton[12]首次通过四倍体饲料甘蓝与四倍体饲料萝卜远缘杂交,得到了成功导入萝卜抗根肿病基因的异源四倍体。但是远缘杂交大多存在受精前杂交不亲和,受精后杂种夭亡或不育以及后代疯狂分离等障碍[13],为了提高芸薹属远缘杂交成功率,Zhang等[14]利用胚抢救培养3种白菜型油菜分别和甘蓝杂交后的杂种后代,培养基(MS+1.0 mg·L-1BA + 0.1 mg·L-1NAA)在3种组合的胚抢救中均表现很好的促进作用,之后通过秋水仙素加倍,从而获得甘蓝型油菜。Pelletier[15],Yamanaka[16]等通过原生质体融合的方法获得了萝卜与芸薹属杂种。Leino等[17]利用体细胞融合技术对细胞质诱导的雄性不育拟南芥与甘蓝型油菜杂交,最终筛选出恢复系植株。富贵等[19]通过甘蓝品种与“大黄油菜”为双亲远缘杂交,利用胚抢救技术,获得了甘蓝型油菜的新种质。【本研究切入点】远缘杂交已成为获得新品种,改良现有品种的重要方法。通过种间杂交将芥菜型油菜(B.juncea,AABB,2n=36)的抗病、抗虫、分枝性强、黄种皮等优良遗传特性导入甘蓝型油菜,可改良现有甘蓝型油菜的品质,丰富油菜种质资源。Roy[18]等为将芥菜型油菜B基因组上抗黑胫病的基因导入甘蓝型油菜,利用均抗根肿病的两亲本甘蓝型油菜与芥菜型油菜杂交,培育出的杂种一代成株期抗黑胫病。Wan等[19]也通过远缘杂交得到了广泛变异的甘芥杂种,丰富了甘蓝型油菜的种质资源。为提供芥菜型油菜和甘蓝型油菜杂交后代细胞学证据以及为育种提供更加丰富的变异材料,本研究以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本,以正反杂交试验得到的F1和正交F1自交得到的F2代为供试材料,对其进行了形态学和细胞学分析,并且进行了杂种后代花粉育性以及杂交亲和性分析,为拓宽甘蓝型油菜的遗传基础提供了理论依据。
1 材料与方法
1.1 材料
芥菜型油菜品种为严小村芥,甘蓝型油菜品种为青油14 号。材料均由青海省农林科学院春油菜研究所提供。
1.2 试验方法
1.2.1 性状调查 分别于苗期、蕾期、开花期、成熟期,调查植株的叶型、叶色、叶的大小,叶片表面有无蜡粉、刺毛性状,花蕾形态,雌雄蕊发育情况,并进行拍照记录。
1.2.2 花粉的可染性 晴天采集杂交后代新开的2朵花,取其长、短雄蕊各1枚,将花粉均匀涂在载玻片上,用1 %醋酸洋红染液染色,轻轻盖上盖玻片,在显微镜的10倍镜头下观察计数。大、圆且被染为红色的为可育花粉,小而瘪且未能染色的为不育花粉。
1.2.3 杂交亲和性试验 正交:严小村芥×青油14号;反交:青油14号×严小村芥。
每个杂交组合选取4~8株,记录授粉花蕾数与杂交时间,成熟时单独收取,调查结角数,结籽率,计算结角率与亲和指数,参照刘忠松、徐爱遐等[21-22],以衡量杂交结实力高低。
结角率=收获的角果数/授粉花朵数×100 %
亲和指数=饱满的籽粒数/授粉花朵数
1.2.4 细胞学观察 有丝分裂观察。于晴天上午8:30-10:30在田间取幼嫩花蕾(2~3 mm),置于冰盒带回挑取子房,用 8-羟基喹啉避光暗处理3.5 h后,置于卡诺固定液(乙醇与冰醋酸体积比为3∶1)中固定24 h以上。观察时挑取固定好的子房于1 mol/L 的盐酸中60 ℃水浴6 min左右,单蒸水中漂洗后,用改良的卡宝品红染色压片,于显微镜下观察并记录体细胞染色体数目。
减数分裂观察。于盛花期取幼嫩花蕾置于卡诺固定液中,固定并换洗花蕾直至褪色为白色,4 ℃冰箱保存。挑取幼嫩花蕾的花药于1 mol/L 的盐酸中 60 ℃水浴3 min,取出后放置于单蒸水中漂洗后,用卡宝品红染色压片,在显微镜下观察花粉母细胞(Pollen Mother Cells)减数分裂各时期的染色体行为。
1.2.5 数据处理 利用Excel和SAS软件进行统计分析。
2 结果与分析
2.1 芥甘杂交亲和性分析
以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本分别做正反交,统计杂交后的结角数与所得种子数(表1),芥×甘杂交共授粉175朵花蕾获得1320粒种子,亲和指数7.54,甘×芥杂交共授粉139朵花蕾获得13粒种子,亲和指数为0.09,较前人研究报道的0.1~0.5低,说明本试验中两种材料反交时可交配性较差。以芥菜型油菜作母本与甘蓝型油菜杂交,杂交亲和性显著高于以甘蓝型油菜作母本的反交,与前人报道结论相一致[23-24]。
表1 芥菜型油菜与甘蓝型油菜杂交亲和性分析
2.2 杂种F1和F2代形态学分析
2.2.1 杂种F1形态学分析 以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本,分别做正反杂交试验获得F1种子,其正反交得到的真杂种植株形态如图1,植株形态为介于双亲的中间型,基部叶面有刺毛,茎秆粗壮,叶片大,羽状半深裂,分枝性强,且株高均高于双亲。以青油14号为母本得到的杂种苗期叶片羽裂较深,而母本为严小村芥的杂种植株的分枝数比反交杂种多,且花的形态更偏向母本,花瓣不重叠,呈四瓣分离,反交杂种的花更偏向父本,花瓣有部分重叠,正反交杂种无较大的差异。
2.2.2 杂种F2形态学分析 如图2所示,F2群体植株表型分离明显,每株材料形态都不同,大部分植株偏甘蓝型,占总群体的64.8 %,中间型占总群体的28.2 %,偏芥菜型占总群体的7.7 %。偏甘蓝型的植株叶片表面较光滑平展,有缺刻或全缘,偏芥菜型与中间型的植株基部叶片具有刺毛,叶面稍有皱缩。同时,F2花色也出现了淡黄色及乳白色,且花为乳白色的植株雄蕊没有花粉,表现为雄性不育。
2.3 芥甘杂交后代花粉育性分析
对芥甘杂交后代F1和F2代材料进行花粉可染性分析(表2),F1代中共调查了10株,其花粉可染率范围为17.09 %~45.81 %;F2中共调查了22株,花粉可染率范围为0~91.88 %,其中有一株没有花粉,表现雄性不育。
2.4 杂种F1和F2代细胞遗传学分析
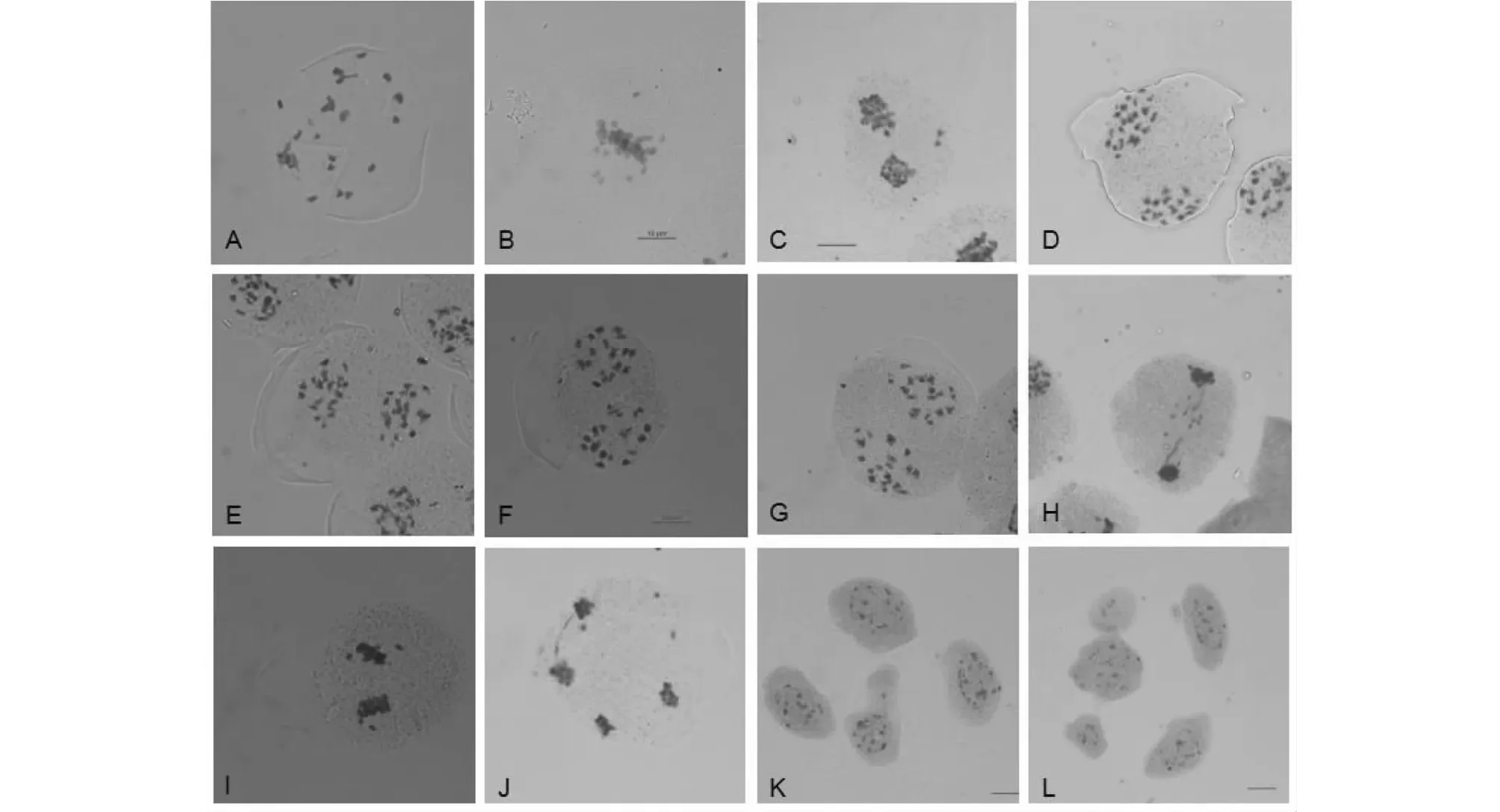
2.4.1 F1细胞遗传学分析 正反交F1染色体组成均为AABC,2n=37(图3),F1减数分裂终变期(图4-A)染色体配对以二价体为主,正交F1二价体数目变化为8~16个,出现12Ⅱ频率最高,除二价体外,绝大部分PMCs均还出现1~3个数目不定的单价体、多价体。反交F1二价体数目变化为8~16个,出现14Ⅱ频率最高,除二价体外,绝大部分PMCs均出现1~4个数目不定的单价体,多价体。正反交PMCs均在中期Ⅰ(图4-B)、后期Ⅰ(图4-C,图5-C)出现异常,染色体落后的频率高,还出现染色体桥。后期Ⅰ(表2)染色体分离方式主要以18:19,其余以17∶20、21∶16、17∶1∶19等。正交中观察到处于四分体时期的PMCs 368个,多分孢子占14.13 %,其中五分孢子较多,还出现较少六分和七分孢子,反交中五分孢子占PMCs总数的5.26 %,六分孢子出现2个。中期Ⅱ、后期Ⅱ也出现许多滞后(图4-I,图4-J),微核(图4-K,图5-L,图5-F,)等现象。

A~D,E~H和I~L分别为亲本以及杂种F1植株的幼苗,花和花蕾。从左至右依次为严小村芥、严小村芥×青油14号F1、青油14号×严小村芥F1以及青油14号。M~N分别为严小村芥×青油14号F1、青油14号×严小村芥F1盛花期植株 A-D, D-H and I-L are young plant, flower, and flower bud of parents and their reciprocal hybrids. The first one is YanXiaoCunJie, and followed by B. juncea × B. napus F1, B. napus× B. juncea F1and Qingyou 14. M-N are mature plants of B. juncea × B. napus F1 and B. napus× B. juncea F1图1 芥甘杂交亲本及正反交杂种F1植株形态Fig.1 Morphological characters of B. juncea, B. napus and their orthgonal and reciprocal hybrids

A~G为芥甘杂交F2,H~L为BC1;表型观察,M~P为F2花观察 A-G are F2 plants, H-L are BC1, M-P are flowers of F2图2 F2表型及花观察Fig.2 Phenotype and flower investigation of F2 and BC1

表2 F1和F2代花粉可育性分析
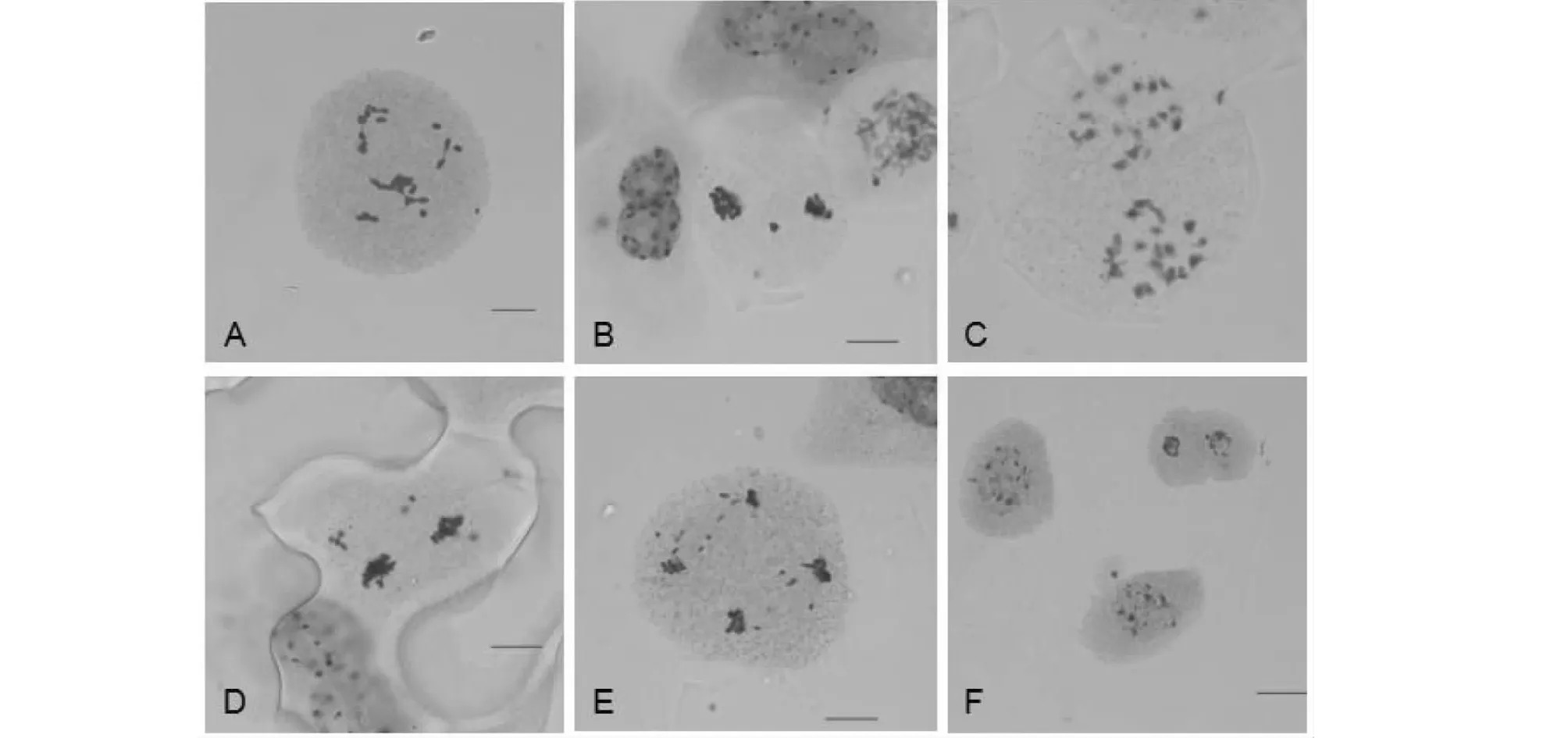
2.4.2 F2细胞遗传学分析 对F2群体进行子房观察,24株植株的体细胞染色体数目均在38条附近(表4),介于33~45条之间(图6)。F2减数分裂终变期染色体配对构型以二价体为主,并且细胞同时有出现不同程度的单价体、三价体、四价体和五价体,部分细胞还出现了六价体。F2PMCs均在减数第一次分裂和第二次分裂中期、后期出现异常,染色体落后的频率高。体细胞染色体数目不同的细胞后期Ⅰ分离方式不同(表5),终变期平均配对规律不同(表6)。
染色体数与甘蓝型油菜染色体数目38 条相同的后代,减数分裂后期Ⅰ染色体分离方式主要以 18∶20,其终变期染色体行为包括(1-3)Ⅰ+(14-19)Ⅱ+1Ⅲ+1Ⅳ+1Ⅴ+1Ⅵ,出现频率最高的是1Ⅰ+17Ⅱ+1Ⅲ(26 %),染色体平均配对规律为1.07Ⅰ+16.72Ⅱ+0.59Ⅲ+0.26Ⅳ+0.1Ⅴ+0.03Ⅵ。
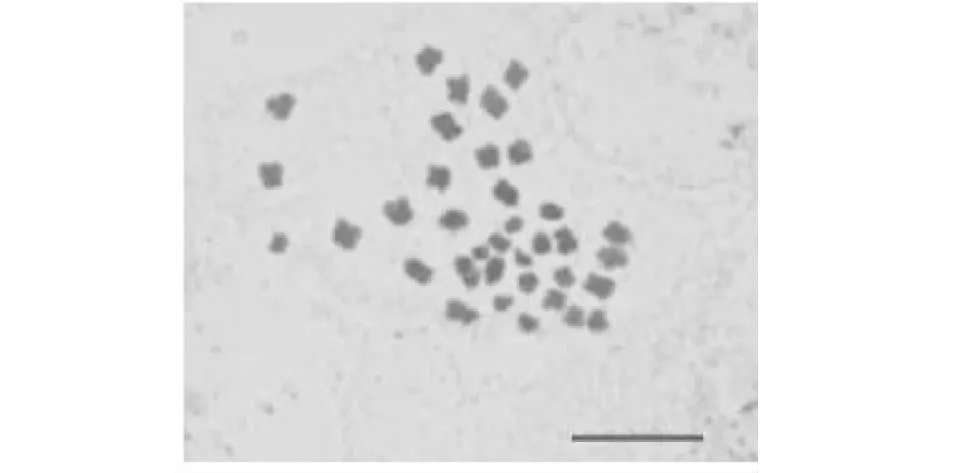
图3 F1体细胞染色体Fig.3 Chromosome of F1
A 终变期;B 中期Ⅰ;C 后期Ⅰ;D~H 后期Ⅰ;I 中期Ⅱ;J 后期Ⅱ; K~L 小孢子时期 A Diakinesis;B MetaphaseⅠ;C AnaphaseⅠ;D-H AnaphaseⅠ;I MetapaseⅡ;J AnaphaseⅡ;K-L Microspore stage图4 正交杂种F1花粉母细胞减数分裂观察Fig.4 Observation on meiosis in the pollen mother cells of orthgonal F1 hybrid
A 终变期;B 后期Ⅰ;C 后期Ⅰ;D 中期Ⅱ;E 后期Ⅱ;F 小孢子时期 A Diakinesis;B AnaphaseⅠ;C AnaphaseⅠ;D MetapaseⅡ;E AnaphaseⅡ;F Microspore stage图5 反交杂种F1花粉母细胞减数分裂观察Fig.5 Observation on meiosis in the pollen mother cells of reciprocal F1 hybrid
3 讨 论
以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本,以正反杂交试验得到的F1和正交杂种所得F2代为供试材料,对其进行了形态学和细胞学分析,并且进行了花粉可染性观察以及杂交亲和性分析。
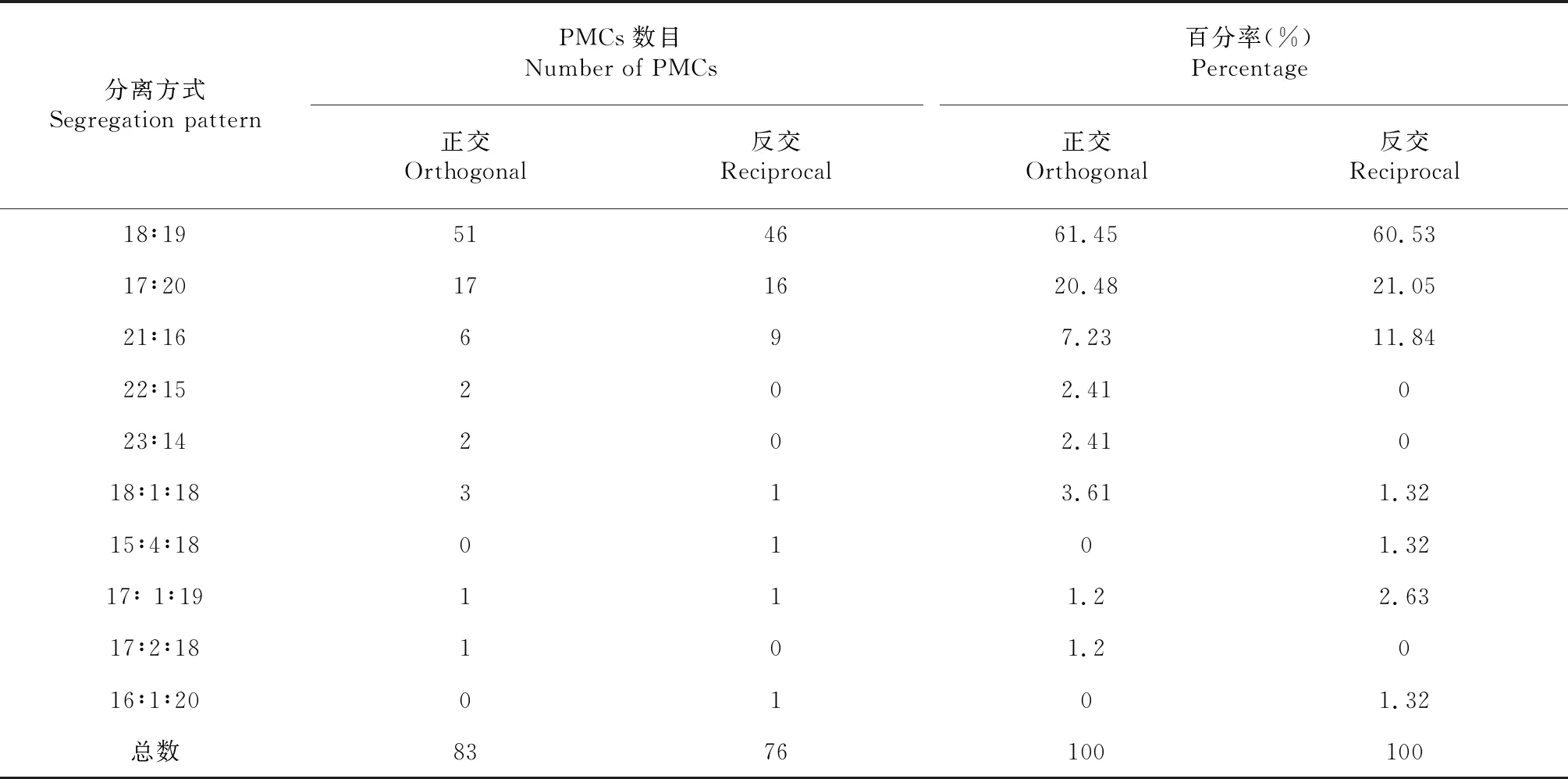
表3 正反交杂种F1 PMCs后期Ⅰ分离方式
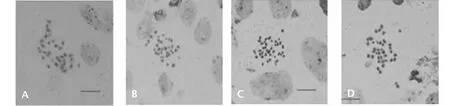
A.36 条染色体;B.38 条染色体;C.42条染色体;D.43条染色体 A.36 chromosomes; B.38 chromosomes; C.42 chromosomes; D.43 chromosomes图6 F2体细胞染色体数目Fig.6 Chromosome of F2

A. 终变期;B. 终变期;C. 中期Ⅰ;D. 中期Ⅱ;E.后期Ⅰ;F.后期Ⅱ A. Diakinesis; B. Diakinesis; C. MetaphaseⅠ;D. MetapaseⅡ;E. AnaphaseⅠ;F. AnaphaseⅡ 图7 杂种F2减数分裂Meiosis in the F2 hybridFig.7 Pollen stainability rate and chromosome number
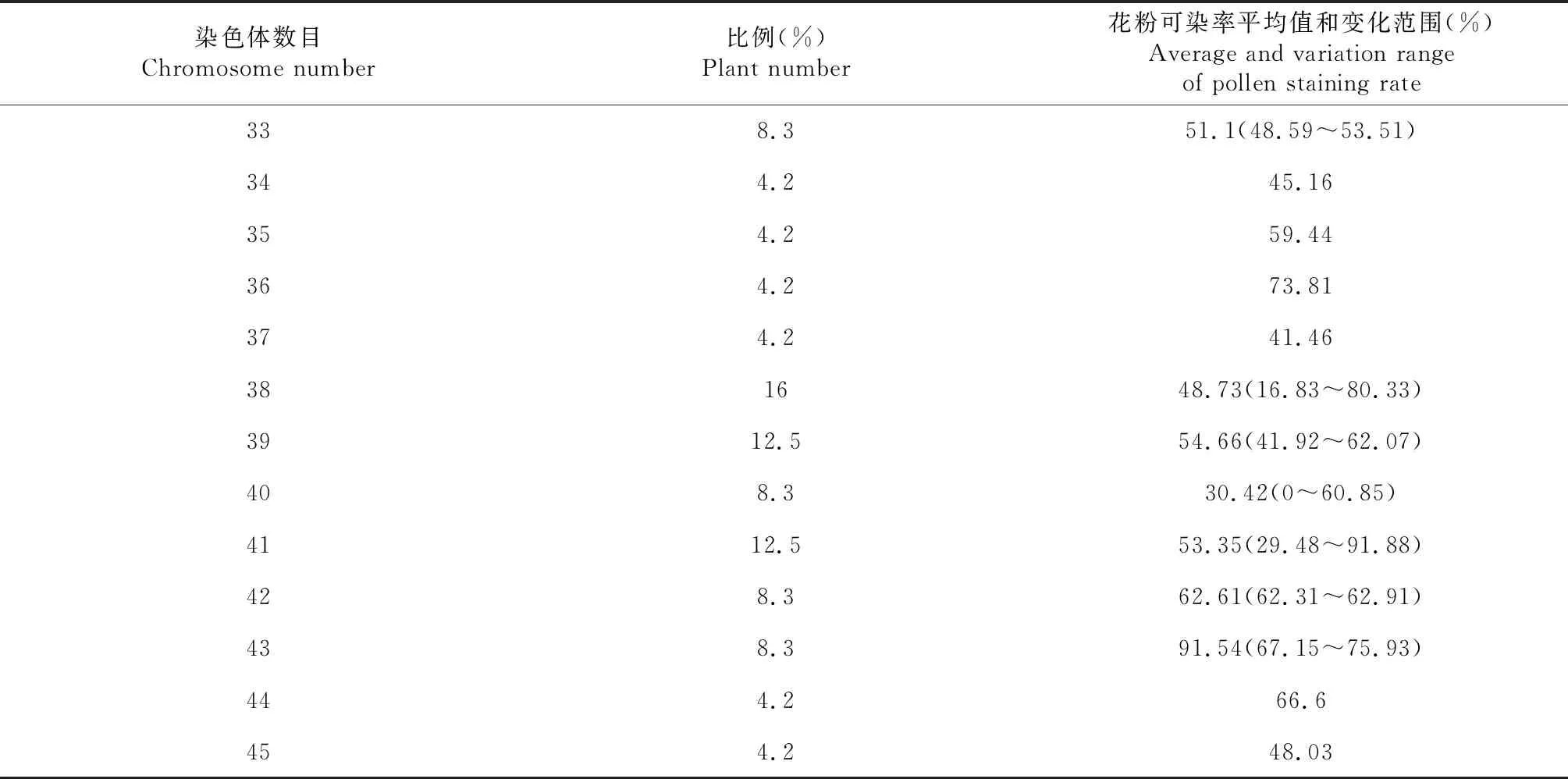
表4 植株花粉可染率和染色体数目

表5 芥甘杂种F2减数分裂染色体行为
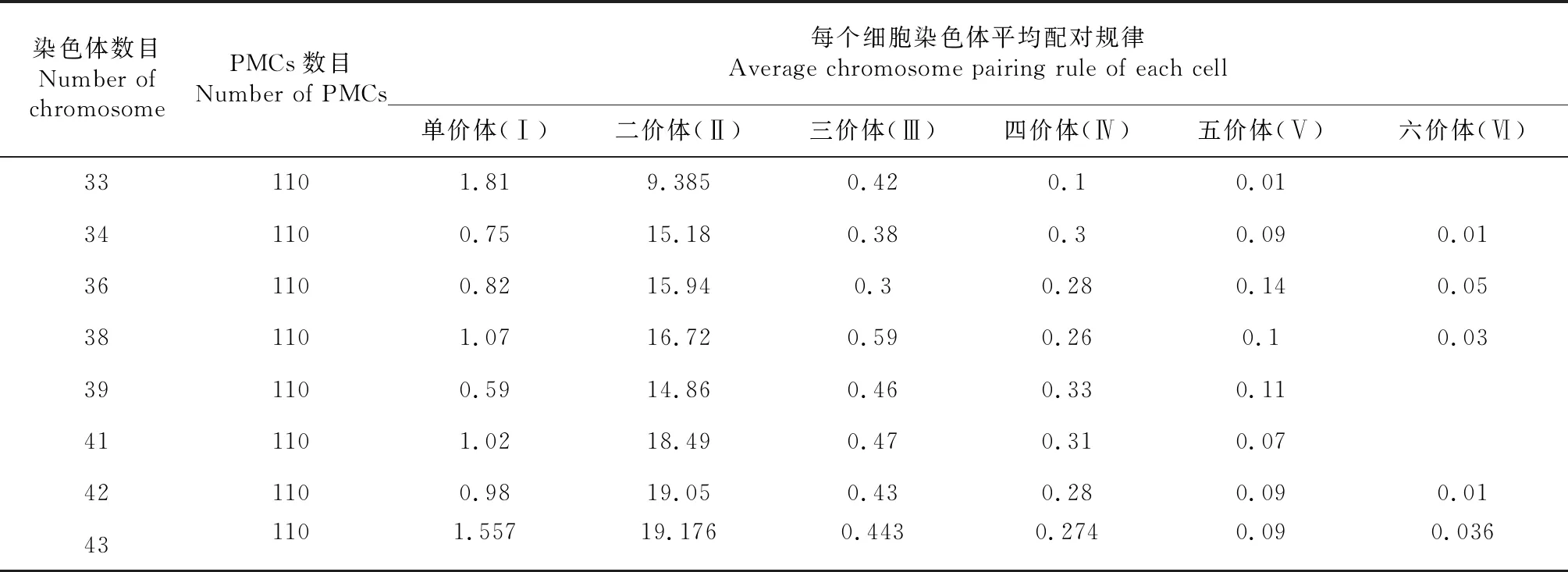
表6 芥甘杂种F2终变期染色体平均配对规律
芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本得到的杂种F1代和F2代表型差异较大,以芥菜型油菜为母本的杂种和与甘蓝型油菜为母本的杂种F1在形态学上基本一致,大部分形态性状超出或介于双亲之间,但在花瓣和叶片等方面表现有些不同,它们分别更偏向于母本。在苗期,F2植株叶片多数偏中间型,但在开花期,表型又偏向于甘蓝型,叶柄抱茎,顶部叶片与甘蓝型相似。F2代染色体数目均在38条左右,表型也接近甘蓝型油菜,连续自交或以甘蓝型油菜回交,可以较快得到新型甘蓝型油菜。
以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本分别做正反交,芥×甘杂交亲和指数7.54,甘×芥杂交亲和指数为0.09,说明以甘蓝型油菜为母本不易得到种子,这一结论与陈纪鹏[25]人工合成甘蓝型油菜与黑芥种间杂种的结论一致。赵洪朝等[26]以1份芥菜型多室油菜与6份甘蓝型油菜杂交,得出为获得较多饱满种子,应采用芥×甘的结论。同时,不同基因型的甘蓝型油菜品种与不同品种的芥菜型油菜杂交的亲和性不同,如甘蓝型油菜×埃塞俄比亚芥[27],但是在甘蓝型油菜远缘杂交中,熊秋芳等[28]证明母本甘蓝型油菜的基因型影响较大。
以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本分别做正反交,F1体细胞染色体数目为37条,符合理论值,但是也有2n=38或34、36的报道[29],其花粉败育程度高。杂种后代F2表型显著分离,染色体数目不定,但大多在38条左右,同时花粉育性有所提高,推测随着世代的增加花粉育性随之提高直至恢复正常,提高了选育新型甘蓝型油菜的成功率。F2出现了不同于亲本的花色,推测可能是因为染色体分布不平衡,同源或部分同源染色体间重组[30],McClintock[31]在烟草种间杂种的研究中对这一推测进行了证实。该研究中F1、F2世代减数分裂终变期均有单价体,多价体等异常配对情况,不排除A、B、C部分同源染色体之间在杂种早期世代存在部分同源配对,从而导致遗传物质在染色体间交换,为创制新型甘蓝型油菜、拓宽其遗传基础提供了可能。
4 结 论
以芥菜型油菜严小村芥与甘蓝型油菜青油14号为亲本进行正反交,正交相比反交更容易获得杂交种子,正反交F1体细胞均具有37条染色体,正反杂种植株形态介于双亲之间,而正交F1植株相比反交F1具有更多的分枝数。以芥菜型油菜为母本杂种F1自交获得的F2表型显著分离,偏甘蓝型较多,染色体数目介于33~45条之间。F2花粉育性提高,自交更易获得种子,F2植株连续自交或以甘蓝型油菜回交、配合细胞学检测,可较快获得新型甘蓝型油菜资源。
