辽宁省稻瘟病菌生理小种及无毒基因鉴定
2020-12-10魏松红朱丽珺王海宁张照茹李昕洋
刘 伟,魏松红,朱丽珺,王海宁,张照茹,李昕洋
(沈阳农业大学植物保护学院,辽宁 沈阳 110866)
【研究意义】由稻梨孢(Pyriculariaoryzae)引起的稻瘟病是目前世界分布最广、危害最重的水稻病害之一[1],近年来,我国辽宁地区稻瘟病的发生也呈现逐年加重的趋势,已成为限制该地区水稻高产和稳产的主要障碍之一[2]。选育和种植抗病品种是防治稻瘟病最有效的措施,水稻对稻瘟病的抗性类型大多数属于小种专化抗病性,由主效抗病基因控制,表现为质量性状,该类抗病类型抗病效能高,遗传方式简单,已成为广泛应用的主要抗病性类型[3-4]。小种专化抗病性符合Flor 的“基因对基因”学说,即抗病基因(R基因)识别病原物的无毒基因(Avr基因),引发不亲和反应[5]。由于稻瘟病菌群体结构复杂,变异速率快,致病力分化严重,导致能克服寄主抗病性的群体快速形成优势,大多数抗病品种在大田推广3~5年后即丧失抗性[6-8]。因此,鉴定稻瘟病菌的无毒基因和生理小种的动态变化,有助于人们根据不同地区流行菌株的无毒基因型选择具有相应抗病基因的水稻品种进行合理布局。【前人研究进展】目前完成初步定位的稻瘟病菌无毒基因至少有40个,成功克隆的稻瘟病菌无毒基因有12个,分别为PWL1、PWL2、AVR1-CO39、Avr-Pita、ACE1、Avr-Piz-t、Avr-Pia、Avr-Pii、Avr-Pik/km/kp、PWL2D、Avr-Pi9和Avr-Pib。刘翔等[9]对2014年分离自湖南桃江病圃丽江新团黑谷的稻瘟病菌进行无毒基因鉴定,结果表明:无毒基因Avr-Pita、Avr-Pish、Avr-Piz、Avr-Piz5、Avr-Pi3和Avr-Pi9出现频率较高。王世维等[10]对2011年分离自辽宁地区的稻瘟病菌进行无毒基因鉴定,结果表明:辽宁省稻瘟病菌主要无毒基因类型为Avr-pita、Avr-pik和AvrPiz-t,AVR1-CO39、Avr-pii和Avr-pia在辽宁省的稻瘟病菌中均未检测到。朱名海等[11]对2014-2015年分离自南繁核心区和非核心区的稻瘟病菌无毒基因携带情况进行PCR检测。结果表明:无毒基因ACE1、Avr-Pik、Avr-Pita、Avr-Piz-t和PWL2在2个区域的稻瘟病菌中分布率较高,无毒基因Avr-Pia在南繁非核心区的稻瘟病菌中未检测到。【本研究切入点】本试验于2017-2018年采集辽宁省7个主产稻区穗颈瘟病标样740余份,通过单孢分离共获得151株供试菌株,采用全国统一的1套7个鉴别品种,对供试菌株进行生理小种鉴定,并对供试菌株进行了10个稻瘟病菌无毒基因携带情况的PCR检测。【拟解决的关键问题】研究结果可为指导辽宁地区水稻品种的选育、布局和轮用以及稻瘟病的综合防控提供理论依据。
1 材料与方法
1.1 供试材料
供试菌株:2017-2018年从辽宁省7个主产稻区(沈阳市、大连市、丹东市、营口市、盘锦市、铁岭市、抚顺市)采集穗颈瘟标样740余份,通过病原菌常规分离和单孢纯化共获得151株供试菌株,其中沈阳25株、大连9株、丹东31株、营口2株、盘锦28株、铁岭34株、抚顺22株。
供试水稻鉴别品种:特特普、珍龙13、四丰43、东农363、关东51、合江18、丽江新团黑谷。
供试培养基:PDA培养基为马铃薯200 g,琼脂20 g,葡萄糖20 g,蒸馏水1000 mL;PD培养液为马铃薯200 g,葡萄糖20 g,蒸馏水1000 mL;TOA培养基为燕麦片30 g,番茄汁200 mL,琼脂30 g,蒸馏水1000 mL。
1.2 试验方法
1.2.1 供试菌株生理小种鉴定 将活化的供试菌株转接至TOA培养基进行产孢培养,培养5 d后,配成孢子悬浮液(每毫升1×106个孢子)备用。将7个供试鉴别水稻品种培养至3叶1心期,参照张海旺等[12](2014)的方法进行离体叶段划伤接种,放置于人工气候培养箱中,模拟7-8月水稻田间的气候条件,培养8 d后,采用全国稻瘟病菌生理小种联合试验组(1980 年)制订的统一标准进行调查,记载鉴别品种的抗(R)感(S)反应类型[13]。
1.2.2 供试菌株无毒基因鉴定 将活化的供试菌株转接至PD培养液中培养8 d,收集菌丝并干燥,利用OMEGA公司真菌基因组DNA提取试剂盒(D3390-01)进行供试菌株DNA提取。根据10个稻瘟病菌无毒基因序列,利用Primer 6和DNAMAN软件设计引物并检验,设计出10对不同无毒基因引物,引物序列见表1。PCR反应体系(30 μl):E-taq酶0.15 μl,10×E-TaqBuffer(MgCl2)3 μl,正向引物和反向引物各0.12 μl(100 μg·mL-1),dNTP 2 μl,DNA模板2 μl,ddH2O补足至30 μl。PCR反应程序:95 ℃预变性5 min,94 ℃变性30 s,55~61 ℃退火30 s,具体依据引物的G/C含量来定,72 ℃延伸30~60 s,具体依据目的片段长度而定,29个循环,72 ℃延伸5 min。琼脂糖凝胶电泳检测:取5 μl PCR产物与1.5×Loading buffer混合液(4∶1)混合后,在1 %琼脂糖凝胶中电泳,DuRed染色后,在凝胶成像系统下拍照。
1.3 数据分析
结合2010-2016年辽宁省稻瘟病菌生理小种种群分布和动态变化,对2017-2018年辽宁省稻瘟病菌生理小种进行分析,结合2014-2016年辽宁省稻瘟病菌无毒基因分布和动态变化,对2017-2018年辽宁省稻瘟病菌无毒基因进行分析。
2 结果与分析
2.1 2017-2018年辽宁省稻瘟病菌生理小种鉴定
由表2可知,供试151株稻瘟病菌经鉴定划分为6群41个生理小种。分别为ZA群的ZA1、ZA3、ZA5、ZA7、ZA9、ZA13、ZA17、ZA21、ZA25、ZA29、ZA31、ZA33、ZA37、ZA41、ZA49、ZA55、ZA57、ZA61,共18个小种;ZB群的ZB1、ZB3、ZB5、ZB9、ZB11、ZB13、ZB17、ZB19、ZB21、ZB23、ZB25、ZB29、ZB30、ZB31,共14个生理小种;ZC群的ZC1、ZC5、ZC11,共3个生理小种;ZD群的ZD1、ZD5、ZD7,共3个生理小种;ZE群的ZE1和ZE3,共2个生理小种;ZF群的ZF1,共1个生理小种。

表1 用于扩增无毒基因的引物
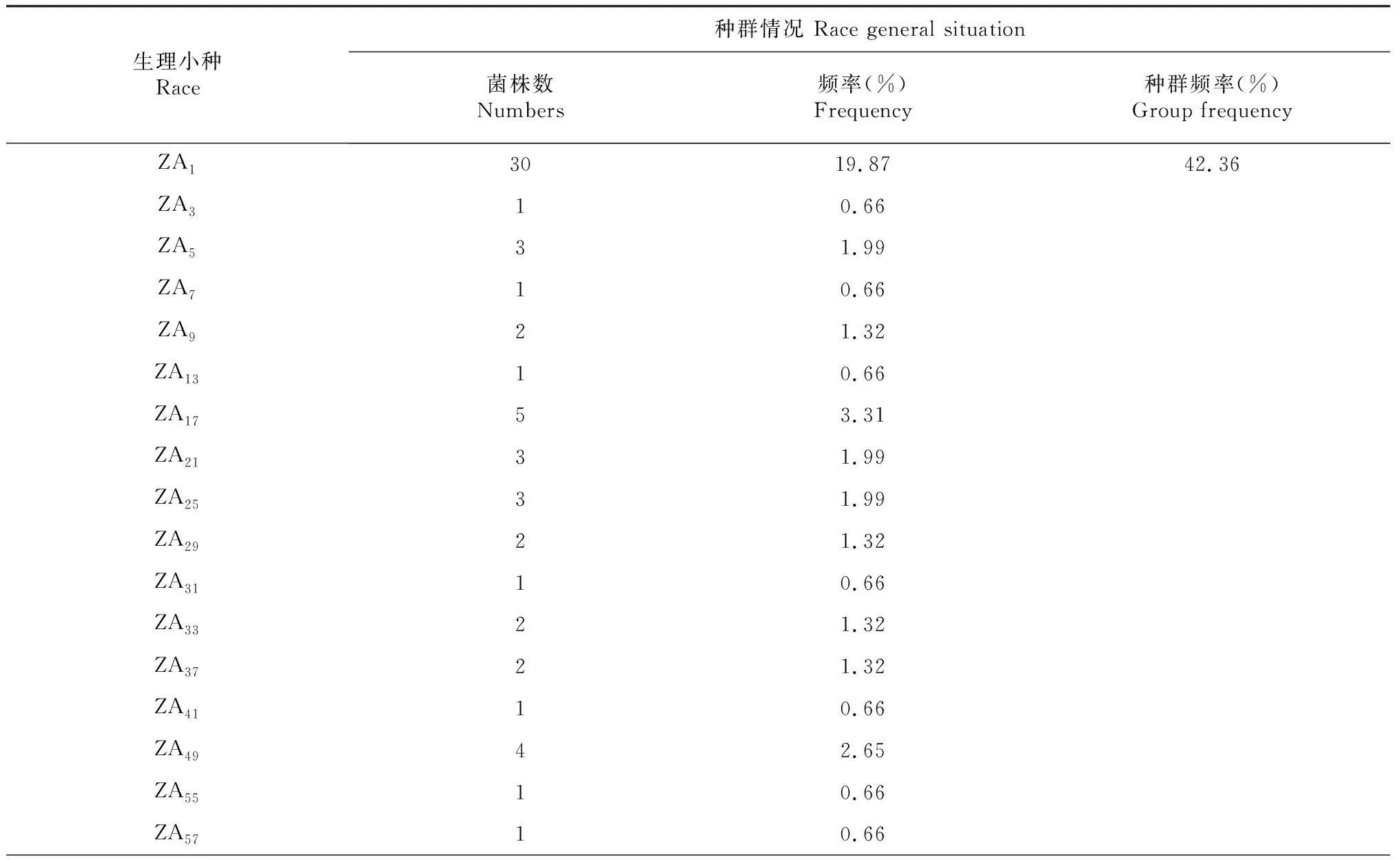
表2 2017-2018年辽宁省稻瘟病菌生理小种鉴定
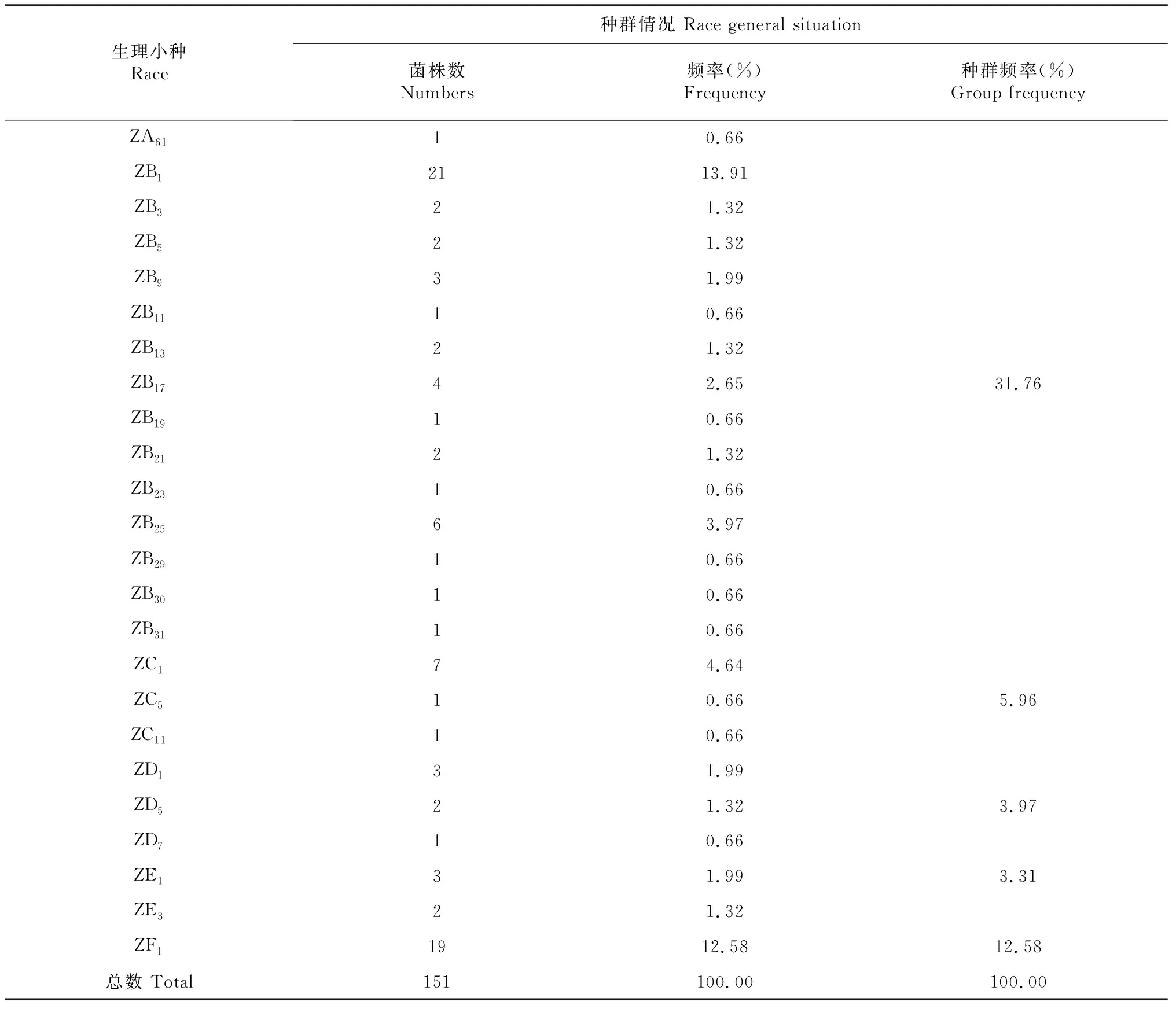
续表2 Continued table 2
2.2 2017-2018年辽宁省稻瘟病菌无毒基因型鉴定
通过设计10对无毒基因特异性引物对151株稻瘟病菌进行PCR检测,结果表明(表3):AVR1-CO39、Avr-pii和Avr-pia在辽宁省稻瘟病菌中未被发现,无毒基因Avr-Pita、AvrPiz-t、Avr-pik、PWL2和ACE1携带频率较高,携带频率分别为:90.07 %、94.04 %、93.38 %、92.72 %和98.68 %,无毒基因Avr-pib携带频率较低,仅为27.81 %。
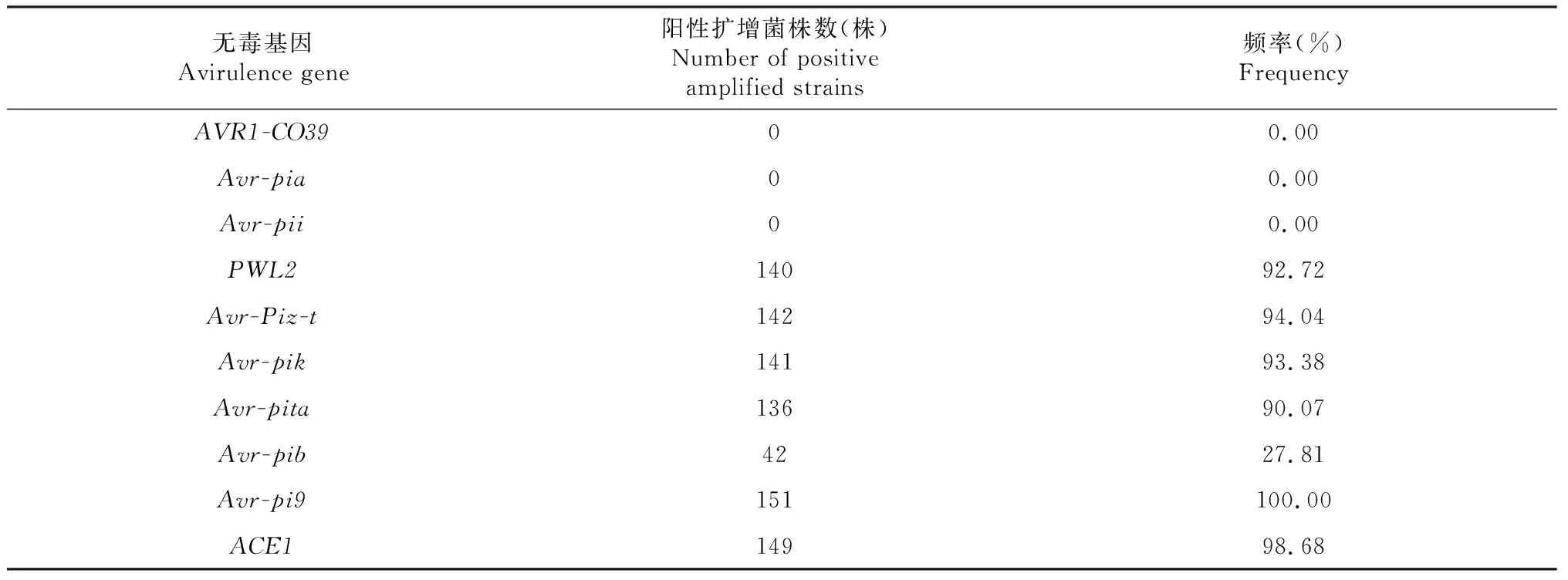
表3 2017-2018年辽宁省稻瘟病菌无毒基因鉴定
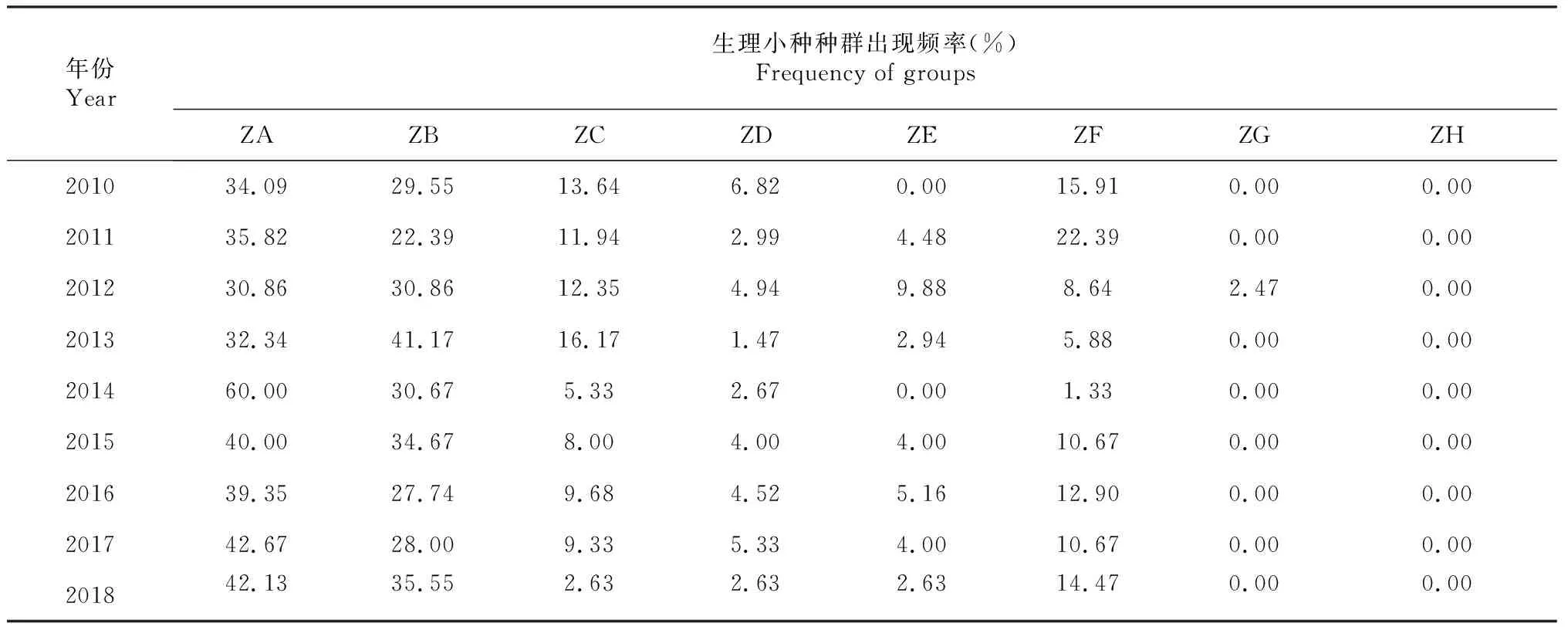
表4 2010-2018年辽宁省稻瘟病菌出现频率
2.3 2017-2018年辽宁省稻瘟病菌生理小种种群动态变化
由表4可知,2010-2018年辽宁省稻瘟病菌生理小种组成中,ZA种群和ZB种群为优势种群,除2013年外,ZA种群均为第一优势种群。多数年份,ZF种群占比仅次于ZA种群和ZB种群,而在2012-2014年ZF种群占比明显减少。ZC种群自2014年起占比明显降低。ZD种群和ZE种群虽然在一定的范围内进行波动,但占比率一直较低。ZG种群仅在2012年出现,ZH种群一直未出现。
2.4 2017-2018年辽宁省稻瘟病菌无毒基因动态变化
由表5可知,2014-2018年辽宁省稻瘟病菌无毒基因组成中,Avr-pi9、ACE1、Avr-Piz-t、Avr-pik、Avr-pita、PWL2携带频率较高。Avr-pi9在2015年携带频率有所下降,在其余年携带频率均在90 %以上。Avr-pib携带频率在2017年有较大幅度上升,为49.33 %,其2014、2015、2016和2018年出现频率分别为2.00 %、13.00 %、10.00 %和6.58 %。2014-2018年均未检测到无毒基因AVR1-CO39、Avr-pii和Avr-pia。
2.5 2017-2018年辽宁省稻瘟病菌生理小种种群分布
2017-2018年辽宁省稻瘟病菌种群监测结果(表6)表明:铁岭、沈阳、丹东、抚顺和盘锦地区稻瘟病菌生理小种种群组成复杂,分别为6群18个生理小种、6群18个生理小种、5群18个生理小种、5群11个生理小种和5群14个生理小种,大连和营口地区相对单一,分别为4群6个生理小种和1群2个生理小种。ZA种群和ZB种群不仅为辽宁地区的优势种群,在各个稻区也是优势种群。
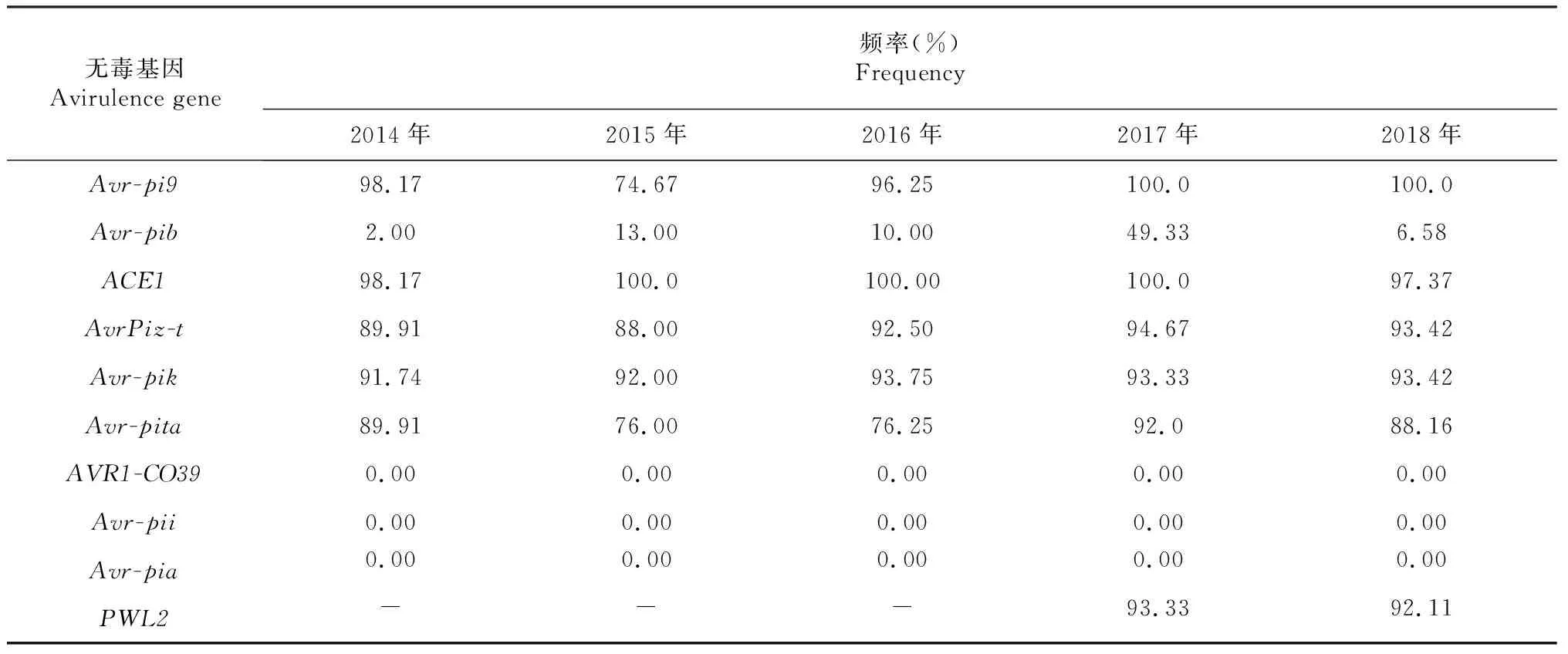
表5 2014-2018年辽宁省稻瘟病菌无毒基因出现频率

表6 2017-2018年辽宁省稻瘟病菌生理小种种群分布
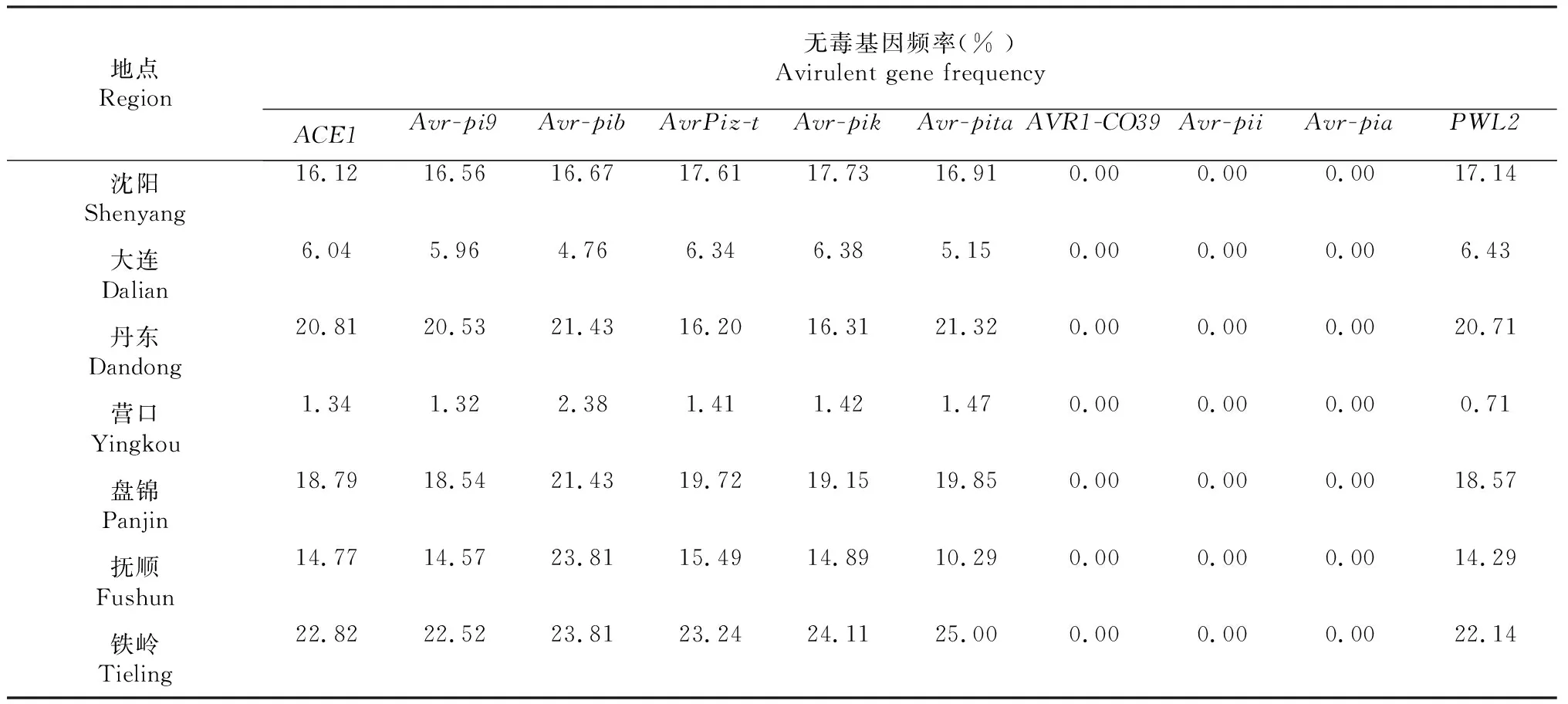
表7 2017-2018年辽宁省稻瘟病菌无毒基因型地理分布
2.6 2017-2018年辽宁省稻瘟病菌无毒基因分布
2017-2018年辽宁省稻瘟病菌无毒基因分布结果(表7)表明:所检测的10个无毒基因,除AVR1-CO39、Avr-pii和Avr-pia外,其它无毒基因在辽宁省各个稻区均检测到。同一地区的不同无毒基因的携带频率基本相同。
3 讨 论
选育和种植抗瘟品种是防治稻瘟病最经济有效的措施之一,但是单一抗病品种长时间大面积推广种植易对田间病原菌造成选择压力,促使生理小种发生变异,进而造成品种抗性减退甚至丧失[14-15]。鉴于稻瘟病菌及其寄主在世界范围内的重要影响,现在已经成为研究植物与病原菌间相互作用的模式生物。根据“基因对基因”学说,水稻中含有1个抗性基因(R基因),在稻瘟病菌中就存在1个相对应的无毒基因(Avr基因)。R基因通过识别Avr基因,启动防卫反应,从而激发水稻对稻瘟病菌的抗性。所以只有在水稻中含有R基因,而同时稻瘟病菌中含有相对应的Avr基因,水稻才表现抗瘟性。水稻抗瘟性研究需要从寄主的抗病基因和稻瘟病菌的无毒基因同时入手,才能系统地解决水稻抗稻瘟病的问题。本研究利用7个水稻鉴别品种对2017-2018年采自辽宁省各个稻区的151株稻瘟病菌进行生理小种鉴定,并根据已克隆的10个稻瘟病菌无毒基因序列设计引物,对供试菌株进行PCR检测。结果表明:供试菌株被划分为6群41个生理小种,ZA和ZB为优势种群,未检测到ZG种群。供试菌株全部携带无毒基因Avr-Pi9,大部分菌株携带无毒基因Avr-Pita、AvrPiz-t、Avr-pik、PWL2和ACE1,无毒基因Avr-pib携带频率较低,无毒基因Avr-Pii、Avr-CO39和Avr-Pia未检测到。
钟宝玉等[16]从2006-2015年近10年间收集并分离了广东各地稻瘟病菌3690个,经接种到中国稻瘟病菌鉴别品种上共鉴定出8群51个生理小种,其中以ZB群、ZC群和ZG群为主要优势种群;陈小林等[17]利用我国7个稻瘟病菌鉴别品种对广西2012-2014年分离获得的稻瘟病菌进行生理小种鉴定。结果表明:2012-2014年分离获得的142株稻瘟病菌菌株分为7群24个生理小种,其中优势种群为ZB,优势生理小种为ZB9和ZB13;沈乐融等[18]针对2013-2017年从江苏省采集标样并分离得到的共计621个稻瘟病菌菌株,利用10对稻瘟病菌无毒基因(PWL1、AVR1-CO39、Avr-Pita、ACE1、Avr-Piz-t、Avr-Pia、Avr-Pii、Avr-Pik、Avr-Pi9和Avr-Pib)特异性引物进行扩增,分析了江苏省稻瘟病菌无毒基因的分布及频率,结果表明:上述10个无毒基因在江苏省均有分布,但出现频率不同,其中平均扩增频率最高的为Avr-Pib,最低的是Avr-Pia;李祥晓等[19]采用7个稻瘟病菌无毒基因(AVR1-CO39、Avr-Pita、ACE1、Avr-Piz-t、Avr-Pia、Avr-Pit和Avr-Pik)特异性引物,对177个采自黑龙江省的稻瘟病菌菌株进行PCR检测,结果表明:7个无毒基因在黑龙江省不同地区均有分布,但出现频率不同,其中频率最高的为ACE1,达到最低的为AVR1-CO39。由以上可知,不同地区稻瘟病菌生理小种和无毒基因组成差异较大,这种不同是由于水稻种植品种变化、品种抗性变化及气候变化等综合因素影响的结果。
除了地区差异外,不同寄主的稻梨孢中无毒基因组成情况也具有较大差异,余欢等[20]对35株草类寄主的梨孢菌进行无毒基因型分析,结果表明:所有供试菌株均没有Avr-Pik/km/kp、PWL3和PWL4基因,以马唐为寄主的菌株中没有ACE1、Avr-Piz-t、Avr-Pita1和PWL1;以牛筋草为寄主的菌株中没有Avr-Pia、Avr-Pita1、Avr-Pita2和PWL2;以狗尾草为寄主的菌株中没有Avr-Pii、Avr-Pita、PWL1和PWL2。无毒基因Avr-CO39可与水稻抗性基因Pi-CO39(t)相互识别,引起水稻的过敏性坏死反应[21]。但Avr-CO39存在于稻瘟病菌的进化早期,并在稻属专化型的进化过程中由于发生转座子插入,使得稻瘟病菌丧失了Avr-CO39活性。稻瘟病菌中并未发现具有完整Avr-CO39序列的菌株。而其他寄主的梨孢菌具有完整的Avr-CO39结构[22-23]。
4 结 论
由于稻瘟病菌生理小种和无毒基因的多样性和易变性,极易导致优良抗瘟水稻品种丧失抗性,水稻品种的抗瘟持久性与其所在地区稻瘟菌生理小种和无毒基因的变化紧密相关[24]。因此,开展不同地区稻瘟病菌生理小种和无毒基因组成和动态变化监测,引导水稻品种的合理布局,是有效预防稻瘟病流行的重要手段,也是稻瘟病可持续治理的长期性工作。辽宁省稻瘟病菌生理小种和无毒基因组成结构复杂,在辽宁省水稻品种布局中,应推广含有Pita、Pi9、Pizt和Pik抗性基因的水稻品种。
