Ezh2通过维持线粒体ATP合成功能促进骨骼肌细胞分化
2020-12-04武晓慧徐玉乔陈佳琪刘婉莹
武晓慧 ,李 青,徐玉乔,陈佳琪,杨 帆,刘婉莹,李 凯
(1.西安医学院肥胖与代谢病研究所,陕西西安 710021;2.空军军医大学第一附属医院病理科暨病理学教研室,陕西西安 710032;3.西安医学院临床医学院,陕西西安 710021)
骨骼肌是人体四大基本组织之一,骨骼肌病变严重影响人体运动功能,甚至危及生命[1]。近年来的研究表明,表观遗传学调控机制与骨骼肌细胞的分化调控及骨骼肌疾病如肌营养不良症、横纹肌肉瘤等有密切联系[2-5],了解骨骼肌细胞的分化调控机制可以为这些疾病的发病机制研究提供理论依据。Ezh2是多梳族抑制性复合物2(polycomb repressive complex 2,PRC2)的催化活性亚单位,由746 个氨基酸组成,可以催化组蛋白H3K27 的甲基化,形成H3K27me3。H3K27me3 一般沉默靶基因的表达[6]。现有的研究结果表明,Ezh2在E9.5 天的胚胎发育部位以及在未分化的骨骼肌成肌细胞中阳性表达,但随着胚胎的生长发育逐渐被抑制[7]。在肌卫星细胞系C2C12 成肌诱导分化过程中,Ezh2表现为减少趋势[7]。在小鼠的骨骼肌卫星细胞中敲除Ezh2,会导致骨骼肌分化不良,肌纤维间脂肪沉着和纤维素增加,小鼠存活率下降,但在分化终末阶段敲除Ezh2则不会明显影响骨骼肌细胞分化[8-9]。总之,目前Ezh2对骨骼肌细胞分化的影响研究结果是其在成肌细胞和肌卫星细胞阶段发挥作用,在分化终末阶段不发挥作用,但Ezh2在更早期的间充质干细胞阶段对骨骼肌细胞分化的具体影响尚不清楚[8,10]。本研究拟观察在间充质干细胞中敲除Ezh2的小鼠骨骼肌分化表型,明确此阶段Ezh2缺失对骨骼肌分化造成的影响。此外,本研究拟进一步观察骨骼肌收缩的能量供应细胞器-线粒体,在Ezh2缺失后是否受到相应的影响,从亚细胞水平探索Ezh2对骨骼肌细胞分化的影响。
1 材料与方法
1.1 实验动物
1.1.1Ezh2基因敲除小鼠的构建 采用Cre-loxp方法构建了Ezh2基因敲除小鼠。Ezh2flox/+雌雄小鼠(B6;129S1-Ezh2tm2Sho/J)及Prx1驱动Cre小鼠(B6.Cg-Tg(Prrx1-cre)1Cjt/J,间充质干细胞条件敲除工具鼠[11-12])由空军军医大学张丰副教授和叶菁教授惠赠。通过Prx1-Cre工具小鼠与Ezh2flox/+小鼠杂交,获得Ezh2flox/+Prx1-Cre+小鼠,再回交获得Ezh2flox/floxPrx1-Cre+小鼠(以下简写为Ezh2F/FPrx1-Cre)。同窝中基因型为Ezh2flox/+或Ezh2+/+Prx1-Cre+同性别小鼠作为对照组。小鼠为SPF 级,饲养于SPF 级屏障实验室,生产许可证编号:SCXK(陕)2019-001;使用许可证编号:SYXK(陕)2019-001。本实验中所有动物操作符合西安医学院和空军军医大学伦理委员会标准。
1.1.2 小鼠基因鉴定 提取鼠尾DNA 进行PCR,反应体系为20 μL,引物序列(5′-3′)如下:Ezh2Forward:CATGTGCAGCTTTCTGTTCA;Ezh2Reve⁃rse:CACAGCCTTTCTGCTCACTG;Prx1-CreForw⁃ard:TCTCTGGCTCTGATGTTGGCA;Prx1-CreRe⁃verse:CGCATAACCAGTGAAACAGC。PCR 结束后用EB 预染的1.6%琼脂糖凝胶100 V 电泳30 min分离DNA,紫外凝胶成像仪成像后根据条带位置判断小鼠基因型。
1.2 实验方法
1.2.1 小鼠骨骼肌组织化学染色 脱颈椎法处死小鼠,取小鼠小腿腓肠肌部位骨骼肌,立即进行冰冻切片,切片厚度10 μm,行苏木素-伊红染色(HE染色)、改良Gomori 三色染色(Modified Gomori Tri⁃chrome,MGT)及四氮唑还原酶(NADH-tetrarolium redutase,NADH-TR)染色。
HE 染色:用甲醛固定切片上的组织后进行常规染色,脱水、透明后树胶封片。
MGT 染色:Harris 苏木精染色10 min 后用水冲洗,Gomori 氏溶液浸泡30 min,0.2%醋酸浸洗1 min,酒精脱水,二甲苯透明,树胶封固。染色后,细胞核应呈红色,肌纤维呈暗绿色,胶原纤维呈亮绿色,线粒体呈红色[13]。
四氮唑还原酶(NADH-tetrarolium redutase,NADH-TR)染色:将切片放入37 ℃孵箱内孵育于含有Tric 缓冲液(PH7.4)、硝基四氮唑和NADH的染色液中30 min,蒸馏水洗,甘油明胶封固。染色后,Ⅰ型肌纤维呈蓝紫色,Ⅱ型肌纤维呈浅灰色[14-15]。
1.2.2 免疫组化染色 腓肠肌组织用40 g/L 甲醛溶液固定48 h,常规脱水、浸蜡、包埋、切片厚度3 μm,切片脱蜡后过梯度酒精至水,柠檬酸-柠檬酸钠缓冲液高压修复抗原(pH6.0);体积分数3%H2O2孵育10 min;体积分数5% 羊血清室温封闭15 min;用1∶300 稀释的细胞色素C(Cytochrome c,CytC,Abcam,美国),湿盒内4℃孵育过夜;生物素标记的羊抗兔IgG 二抗37 ℃孵育15 min;辣根酶标记链霉卵白素工作液37 ℃孵育15 min,DAB 显色,苏木素衬染核30 s,常规脱水透明,中性树胶封片。
1.2.3 电镜分析 取腓肠肌组织,2.5%戊二醛固定液固定24 h 后,将组织块切成1 mm3左右大小,注意纵横方向,送空军军医大学第一附属医院病理科电镜室制样,透射电镜观察并照相。
1.2.4 ATP 浓度测定 采用ATP 检测试剂盒(S0026B,碧云天)进行检测。样品每20 mg 组织加入100 μL 裂解液后进行匀浆。4 ℃13 800 ×g离心5 min,取上清,用于后续的测定。ATP 浓度的测定:加100 μL ATP 检测工作液到检测孔内,室温放置3 min,消耗本底ATP。在检测孔内加入20 μL 样品或标准品,迅速混匀,2 s 后用荧光酶标仪进行测定,根据标准曲线计算出样品中ATP 的浓度。用BCA 蛋白浓度测定试剂盒测定样品中的蛋白浓度。ATP 的浓度以nmol/ng 蛋白的形式表示。
1.2.5 Western blot 法检测蛋白表达量 取小鼠腓肠肌组织约100 mg,提取总蛋白,测定小鼠骨骼肌分化标志物Myogenin、Desmin 和Myosin[15-18]的蛋白表达量。采用BCA 法测蛋白浓度(上海,碧云天),样品上样量为20 μg/孔,电泳结束后转至PVDF 膜,将膜用50 g/L 脱脂奶粉室温封闭1 h,1×PBST 洗膜5 min×3 次,置于1 抗中4℃摇床孵育过夜(兔H3K27me3 抗体,Abcam(15 ku);兔Ezh2抗体,CST(98 ku);鼠β-Tubulin 抗体,天津三箭(55 ku);兔GAPDH 抗体,武汉三鹰(36 ku),兔Myosin 抗体,碧云天(19 ku);兔Desmin 抗体,碧云天(53 ku);小鼠Myogenin 抗体,SANTA CRUZ(34 ku);兔CytC 抗体,Abcam(14 ku)。1×PBST 洗膜5 min×3 次,将膜置于二抗(中衫金桥羊抗兔或羊抗鼠IgG,浓度均为1∶5 000)中室温孵育1 h,再用PBST 洗膜3 次,ECL 发光液发光。
1.2.6 逆转录荧光定量聚合酶链式反应(Real⁃time RT-qPCR)法检测基因相对表达量 检测骨骼肌细胞标志物Myogenin、Ckm、p16Ink4a[19]、Msx1[20]、Atf3[21]、Pax3[22]、MhcIIb和线粒体标志物COX1、CS、CytC、Tfam[23]、Pgc1α[24]、IDH3a的基因表达,取小鼠腓肠肌组织,使用RNA 提取试剂盒(日本TaKaRa)提取总RNA。反转录为cDNA,SYBR Green master mix(日本TaKaRa)催化下,在Real-Time PCR 仪(美国Bio-Rad)上进行扩增。待测基因的引物由大连宝生公司设计并合成,所测基因英文全称及引物序列信息见表1、2。以小鼠GAPDH 为内参。
1.3 统计学分析
采用SPSS 20.0 统计软件进行数据分析,计量数据呈正态分布时以均数±标准差()表示,两组间比较且方差齐时采用独立样本均数t检验,方差不齐的数据采用校正t检验。检验为双侧,以P<0.05 为差异有统计学意义。
2 结果
2.1 Ezh2 基因敲除小鼠骨骼肌细胞分化不良
经观察,Ezh2基因敲除小鼠的体型较小、体质量较轻,精神状态、毛发光泽度、饮食以及活动情况如常,但敲除小鼠出生后死亡率较同窝对照组高,繁殖能力下降。敲除小鼠前肢出现向内曲屈畸形。取6周龄小鼠进行实验。经检测,敲除组小鼠骨骼肌中Ezh2(t=3.327,P=0.029 2)和H3K27me3(t=4.330,P=0.012 3)的蛋白含量降低,表明成功构建了Ezh2基因敲除小鼠(图1A,均P< 0.05)。小鼠骨骼肌切片HE 染色结果显示,敲除组肌纤维粗细不等,核一般在周边,但部分细胞核发生内移现象,如图1B 右图绿色箭头所指处,此为病理性改变。小鼠骨骼肌分化相关基因中Myogenin(t=7.822,P=0.001 4,校正t检验)和MhcIIb(t=4.668,P=0.043 0,校正t检验)的mRNA 表达均明显减少(均P<0.05;图1C),差异有统计学意义,其余基因表达差异无统计学意义。敲除组Myogenin(t=9.126,P=0.000 8)蛋白表达量下降,差异有统计学意义,但Desmin(t=1.767,P=0.152 0)和Myo⁃sin(t=2.241,P=0.088 5)蛋白表达差异无统计学意义(图1D)。

表1 被测基因英文全称Table 1 Full English name of detected genes
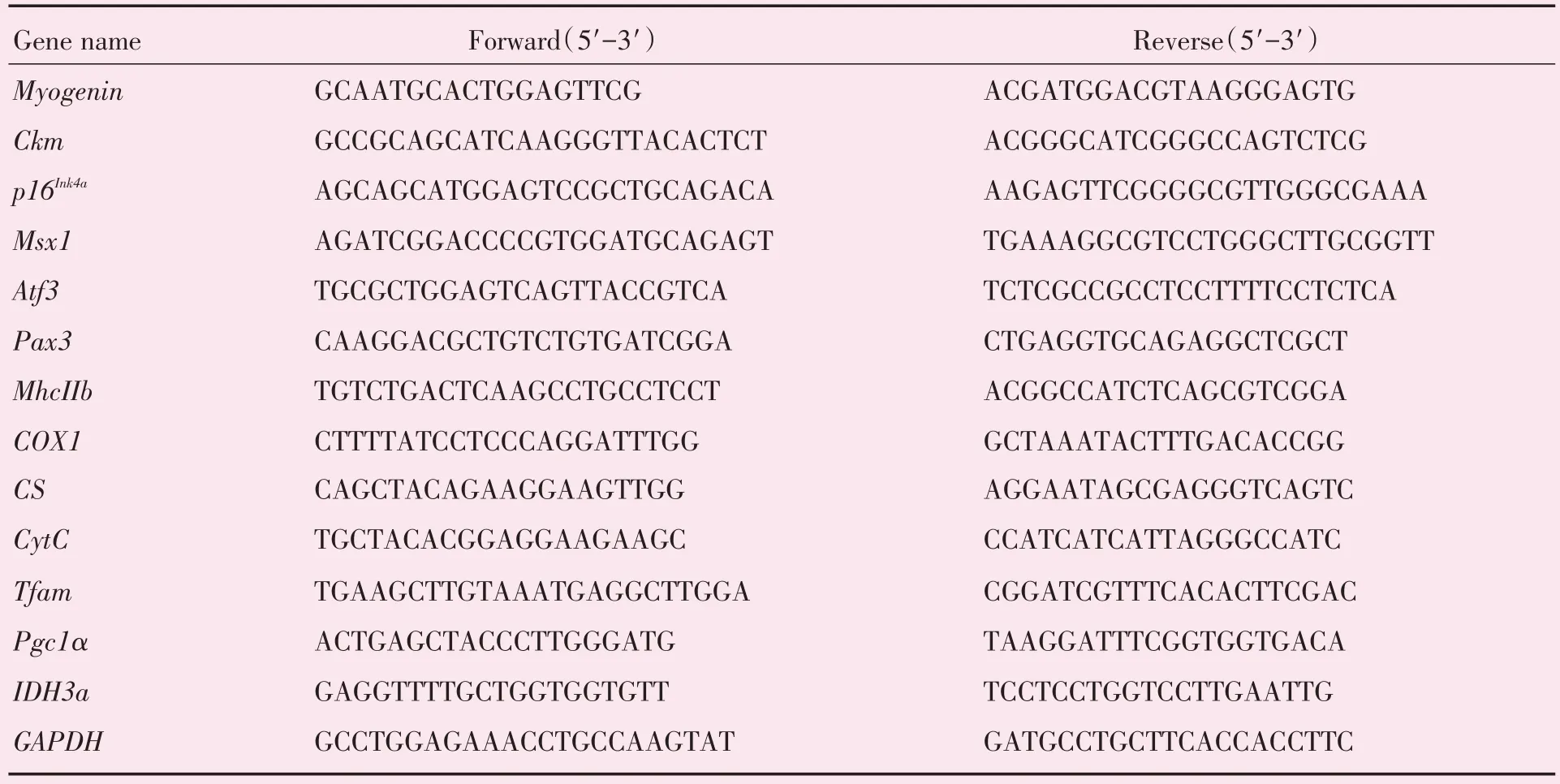
表2 被测基因引物序列信息Table 2 Primer sequence information of detected genes
2.2 Ezh2 敲除小鼠骨骼肌内线粒体数量增加
MGT 染色显示,骨骼肌细胞间无明显胶原纤维增生。NADH-TR 染色显示敲除组肌纤维粗细不等,I 型肌纤维内NADH 增多,提示线粒体数量增加或NADH 增加。细胞色素C 免疫组化染色显示敲除组骨骼肌I 型肌纤维内线粒体数量增加或CytC 表达增加(图2A)。为进一步确定表型,采用电镜在骨骼肌横纵两个切面上进行了观察,横断面上发现肌膜下线粒体数量明显增加,呈团块状堆积(图2B)。纵切面发现骨骼肌肌原纤维粗细不等,肌原纤维间的线粒体异常增加。未见线粒体水肿及明显的嵴断裂。对照组Z 线、明带和暗带清晰可见,敲除组明带清晰度略下降(图2C)。
2.3 Ezh2 敲除小鼠骨骼肌内线粒体可能为代偿性增加
图1 Ezh2 基因敲除小鼠骨骼肌分化表型Fig.1 Differentiation phenotype of skeletal muscle in Ezh2 knockout mice
为了判别敲除组骨骼肌细胞内线粒体增加的表型是Ezh2缺失后促进了线粒体增生还是线粒体功能异常所导致的代偿性增生,进行了一系列实验。经检测,线粒体结构和生成相关基因的表达有不同程度的升高,敲除组线粒体相关基因CytC(t=7.248,P=0.0185)、Tfam(t=4.927,P=0.0388,校正t检验)、Pgc1α(t=8.183,P=0.0012,校正t检验)的表达明显高于对照组(图3A),差异有统计学意义。细胞色素C 蛋白表达增加(t=3.009,P=0.0396;图3B),差异有统计学意义。经测定,敲除组骨骼肌内ATP 含量明显少于对照组(t=4.189,P=0.0138;图3C)。取2 周龄小鼠进行实验,此阶段小鼠刚开始进行自主活动,此时对骨骼肌的利用还不充分。HE 染色结果显示,2 周龄小鼠敲除组仍存在肌纤维粗细不等,直径较为纤细,提示骨骼肌纤维分化不良,但NADH-TR 染色两组无明显差别(图4A、B),说明线粒体增加并不明显。间充质干细胞是骨骼、脂肪、骨骼肌等细胞共同的前体细胞,本实验小鼠的脂肪组织也发生了Ezh2敲除(图4C)。因棕色脂肪富含线粒体,我们对6 周龄小鼠的棕色脂肪细胞中的线粒体进行了电镜观察,两组胞浆内均满布线粒体,并未发现敲除组线粒体数量的增加,线粒体形态无明显异常(图4D)。综合上述证据,推测Ezh2缺失后由于线粒体ATP 产生能力下降,机体为了满足运动需求而促使线粒体数量发生了代偿性增加。

图2 小鼠骨骼肌细胞特殊染色、免疫组化染色及电镜照片Fig.2 Special staining,immunohistochemical staining and electron micrograph of skeletal muscle fibers of mice
3 讨论
本研究利用在间充质干细胞中敲除Ezh2的小鼠,观察了骨骼肌细胞分化的表型。研究所用工具鼠的Cre 酶驱动基因Prx1在胚胎E10.5 天有明确的表达[25],与文献报道的Ezh2在胎鼠体节发育阳性表达最强阶段的E9.5 天时间接近,利用Prx1可以较完全地敲除胎鼠间充质干细胞中的Ezh2。实验结果表明,在出生后2 周龄和6 周龄的Ezh2敲除小鼠,均观察到了骨骼肌细胞分化不良,具体表现为肌纤维粗细不等,肌原纤维粗细不均,核内移等结构性异常。实验检测了与骨骼肌细胞分化调控密切相关的一部分基因的表达情况,结果显示,Myogenin和MhcIIb的mRNA 表达明显减少,Myogenin 蛋白表达量下降,提示这些重要的骨骼肌分化相关基因和蛋白的表达受到影响,与形态学表型相一致。这些研究结果表明,Ezh2在间充质干细胞阶段对于骨骼肌细胞的分化是必须存在的,否则将引起分化障碍。
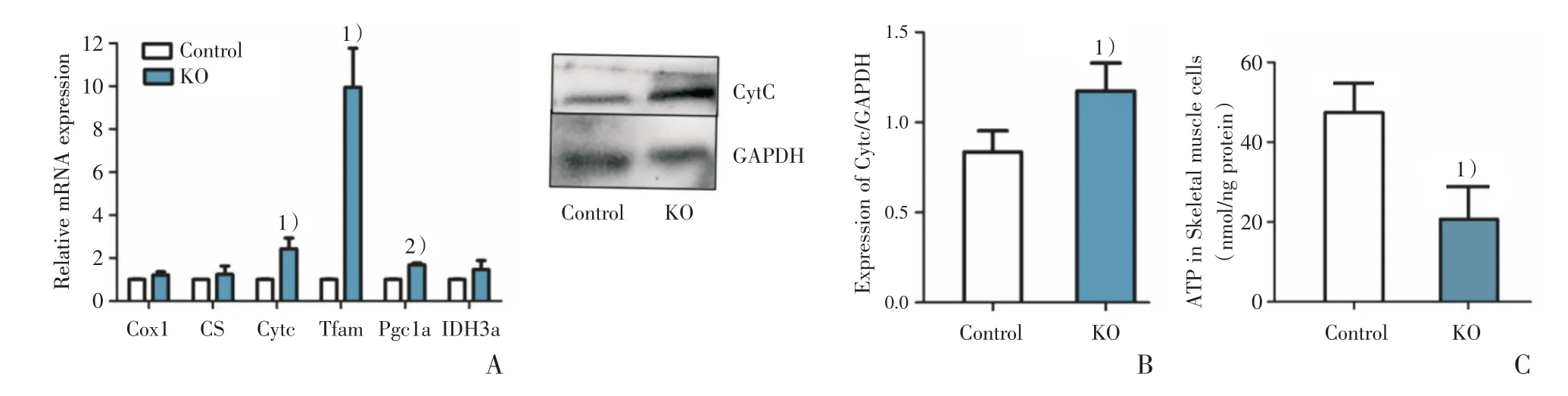
图3 骨骼肌线粒体相关基因表达、CytC 蛋白表达和ATP 含量测定结果Fig.3 Expression of mitochondrial related genes,CytC protein and ATP content in skeletal muscle

图4 2 周龄小鼠骨骼肌HE 和NADH-TR 染色及6 周龄小鼠背部棕色脂肪电镜照片Fig.4 HE and NADH-TR staining of skeletal muscle of 2-week-old mice and electron micrograph of brown adipocytes in the back of 6-week-old mice
骨骼肌是高能量需求组织,尤其是运动时,对能量的需求明显增多[25]。骨骼肌收缩时需要大量的能量用于离子泵、横桥摆动、细胞信号传递等,三磷酸腺苷(adenosine-triphosphate,ATP)为肌纤维收缩的直接能源物质,而线粒体是能量物质氧化产生ATP 的主要场所[26]。骨骼肌细胞的线粒体根据所在位置不同,一般分为肌膜下线粒体和肌原纤维间的线粒体,前者被认为与动脉血循环氧的提供有关,主要为保持肌膜完整性以及各种离子和代谢产物的跨膜转运提供能量,后者则与肌肉收缩时能量的提供有关[27]。既往的研究没有关注Ezh2缺失对于线粒体造成的影响。本研究通过骨骼肌特异性染色,线粒体氧化呼吸链结构蛋白CytC 的检测以及电子显微镜观察了线粒体的形态和分子表型。实验中我们观察到在敲除组Ⅰ型肌纤维(Ⅰ型肌纤维又称慢缩肌纤维,含有较多的线粒体;Ⅱ型肌纤维又称快缩肌纤维,含有较少的线粒体[14])中存在线粒体NADH 异常增多和标志物CytC 异常增多的现象。这促使我们通过进一步实验去寻找原因。在电镜下,我们观察到敲除组6 周龄小鼠肌膜下线粒体和肌原纤维间的线粒体数量均异常增加,呈团块状堆积,但未见线粒体水肿及明显的嵴断裂。线粒体生成相关基因Tfam、Pgc1α的表达均升高,免疫组化染色和West⁃ern blot 结果均证实敲除组的CytC 蛋白含量均加。假设增加的线粒体是功能正常的线粒体,那么不论是动脉血氧的利用还是ATP 的生成均应提高。但实验结果表明,敲除组小鼠骨骼肌内ATP含量是减少的,这提示实验中观察到的线粒体增加很可能是一种代偿性表现。为了观察是否在骨骼肌还未经运动刺激的幼龄动物体内是否有相同的现象,我们观察了2 周龄小鼠的骨骼肌,此时小鼠处在刚刚开始自主运动的发育时期。结果发现,2 周龄Ezh2缺失小鼠骨骼肌内NADH 与对照组并无明显差别。因本实验所用小鼠为间充质干细胞敲除模型,Ezh2敲除范围较为广泛,骨骼肌、骨、脂肪等间充质干细胞来源的组织内均发生了敲除,我们又选取了富含线粒体的棕色脂肪组织作为观察对象,观察线粒体表型。经观察,棕色脂肪细胞内线粒体并无增加现象,线粒体形态亦无明显异常。这说明,Ezh2缺失未能在其他组织细胞引起线粒体的增加,进一步间接地证明6 周龄小鼠骨骼肌内线粒体增加的原因很有可能是Ezh2缺失引起线粒体ATP 产生能力不足的情况下,机体为了满足自身的运动需求,出现的一种代偿性调节,使得线粒体数量增加[28]。目前可以确定,Ezh2的缺失损害了骨骼肌线粒体产生ATP 的能力,但因线粒体ATP 产生的关键环节非常多,具体哪个或哪些环节发生了障碍,尚有待进一步系统性的实验进行探索。
综上所述,本研究的结果证实,在小鼠间充质干细胞成肌分化阶段,Ezh2发挥了重要的作用,Ezh2在本阶段促进骨骼肌细胞的分化。本研究还发现Ezh2缺失后会对骨骼肌细胞内线粒体产生ATP 的能力造成不良影响,线粒体代偿性增加,具体环节有待进一步的研究。
