miR-520a-3p调控宫颈癌细胞因子分泌的信号通路*
2020-11-30刘红艳鲁启洪黄楚鹰
刘红艳, 鲁启洪, 黄楚鹰
(1.湖北省恩施土家族苗族自治州中心医院 肿瘤放疗中心, 恩施 445000;2.湖北省恩施土家族苗族自治州中心医院神经内科, 恩施 445000;3.湖北省恩施土家族苗族自治州中心医院肿瘤一科, 恩施 445000)
宫颈癌的发病率和死亡率在女性生殖道恶性肿瘤中排名第二[1]。每年全世界有近500,000例新病例,其中一半死于宫颈癌[2]。宫颈癌主要由持续感染高危人乳头瘤病毒(human papillomavirus, HPV)引起。在大多数情况下,HPV感染是自限性的,并且可以通过体液和细胞介导的免疫应答来根除。这表明免疫调节可能在宫颈癌发生中起重要作用。炎症是肿瘤进展的关键组成部分,因为癌症是从感染和炎症部位,尤其是慢性炎症部位生长的。已有研究表明宫颈癌细胞能够分泌细胞因子,并促进肿瘤的生存、迁移和增殖等过程[3]。因此,探讨宫颈癌细胞分泌细胞因子的分子机制对治疗肿瘤有积极意义。核因子-κB(nuclear factor-κB, NF-κB)是一种核转录因子,调节各种基因的表达,包括细胞因子,它们在细胞凋亡,肿瘤发生,各种自身免疫疾病和炎症中起关键作用[4]。miR-520a-3p能够调控多种癌细胞的增殖,凋亡和转移[5, 6]。此外,Zhu H等人[7]通过高通量测序发现miR-520a-3p在宫颈癌中表达量较低。然而,其在宫颈癌中的作用尚不明确。目前,从诊断到预后,可能在治疗和预防宫颈癌中发挥作用,所有抗癌方法都在评估细胞因子的作用[8-10]。基于此,本研究以宫颈癌HELA细胞为研究对象,旨在探讨miR-520a-3p调控宫颈癌细胞分泌粒细胞集落刺激因子(granulocyte colony stimulating factor, GCSF), 粒细胞巨噬细胞集落刺激因子(granulocyte-macrophage colony stimulating factor, GM-CSF), 白介素2(interleukin-2, IL-2), 白介素3(interleukin-3, IL-3), 白介素4(interleukin-4, IL-4), 白介素5(interleukin-5, IL-5), 白介素6(interleukin-6, IL-6), 白介素9(interleukin-9, IL-9), 白介素10(interleukin-10, IL-10), 白介素12 p40(interleukin-12 p40, IL-12 p40),白介素12 p70(interleukin-12 p70, IL-12 p70), 白介素13(interleukin-13, IL-13), 白介素17(interleukin-17, IL-17), 干扰素γ(interferon-γ, IFN-γ), 单核细胞趋化因子-1(monocyte chemotactic protein-1, MCP-1), 调控T细胞表达和分泌因子(regulated on activation normal T cell expressed and secreted, RANTES), 肿瘤坏死因子α(tumor necrosis factor α, TNF-α)等细胞因子的分子机制,为治疗宫颈癌提供理论基础。
1 材料与方法
1.1 材料
RELA, GAPDH抗体购自cst公司(货号:59674, 5174)。宫颈癌HELA细胞购自上海康朗生物科技有限公司(货号:Y-01663)。RPMI-1640培养基购自上海北诺生物科技有限公司(货号:R1145-500ML),胎牛血清购自Thermo Fisher Scientific公司(货号:10099-133)。pMir-Glo miRNA功能研究载体购自上海海吉浩格生物科技有限公司(货号:HH-LUC-016)。蛋白酶抑制剂Cocktail(不含EDTA, mini片剂)购自bimake公司(货号:B14011)。PBS购自生工生物工程(上海)股份有限公司(货号:E607008)。青链霉素混合液(100×)购自北京索莱宝科技有限公司(货号:P1400)。2×SDS 蛋白电泳上样缓冲液购自北京鼎国昌盛生物技术有限责任公司(货号:WB-0081)。双荧光素酶报告基因试剂盒购自北京冬歌博业生物科技有限公司(货号:SLDL-100)。蛋白裂解液购自碧云天生物技术有限公司(货号:P0013)。miRNA转染试剂购自上海卉深生物科技有限公司(货号:409-10)。LPS购自上海源叶生物科技有限公司(货号:S11060)。miR-520a-3p的qPCR引物,miRNA NC,miR-520a-3p mimics,miR-520a-3p inhibitor均有上海生工合成,序列参考Zhang R 等人[5]。GCSF, GM-CSF, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12 p40, IL-12 p70, IL-13, IL-17, IFN-γ, MCP-1, RANTES, TNFα的ELISA试剂盒购自上海乔羽生物科技有限公司(货号:QN-PS0040, QY-BM11154, QY-x0605P, QY-BM10220, QN-PS0051, QN-PS0050, QY-BM10211, QY-BM10205, QY-BM10156, QY-BM10178, QY-BM10175, QY-BM10172, QN-PS0179, QY-BM10168, QY-Q11142, QY-BM11185, QY-BM10200)。MCP-5的ELISA试剂盒购自上海恒斐生物科技有限公司(货号:SEC075Mu-1)。
1.2 细胞培养、转染与LPS处理
宫颈癌HELA细胞在含有10%FBS和1%P/S的RPMI-1640培养基中培养。将所有细胞维持在37℃的含5%CO2的培养箱中。将miR-520a-3p mimics或miR-520a-3p inhibitor与miRNA转染试剂混合,静止大约20 min后,缓缓滴入HELA细胞。将HELA细胞与500 ng/ml的LPS一起温育20 h后进行下游试验。其中,转染miRNA NC的为对照组(control)、转染miR-520a-3p mimics的为过表达组(overpression of miR-520a-3p)、LPS处理的为LPS处理组(LPS treatment)、转染miR-520a-3p mimics并用LPS处理的为LPS处理过表达组(LPS treatment + overpression of miR-520a-3p)、转染miR-520a-3p inhibitor的为过表达组(Knockdown of miR-520a-3p)、转染miR-520a-3p inhibitor并用LPS处理的为LPS处理敲低组(LPS treatment + Knockdown of miR-520a-3p)。
1.3 RNA抽取与实时荧光定量PCR
使用TRIzol试剂提取总RNA,并进行cDNA合成。用Bio-Rad CFX96实时系统进行qRT-PCR。将转录水平标准化为甘油醛-3-磷酸脱氢酶(GAPDH)的水平,检测基因表达水平。
1.4 双荧光素酶报告系统试验
将RELA的3’UTR克隆进pMir-Glo质粒,同时克隆RELA的3’UTR的UG突变进pMir-Glo质粒。将每孔总共5×104个HELA细胞接种在24孔板中。培养24 h后,用含有200 ng/ml双荧光素酶报告质粒和40 nmol/L miR-520a-3p mimics混合后转染细胞。使用荧光素酶报告检测试剂盒在转染后24 h通过酶标仪测量荧光素酶活性。所有转染独立重复至少三次。
1.5 免疫印迹试验
使用RIPA裂解缓冲液[50 mmol/l Tris-HCl(pH7.4),150 mmol/l NaCl,1%Nonidet P-40, 0.5%脱氧胆酸钠]从细胞中提取总蛋白质,并使用BCA蛋白质测定试剂盒进行定量[11]。使用多克隆抗RELA(1∶3 000), GAPDH(1∶6 000)抗体作为一抗、抗兔和抗小鼠IgG为二抗。使用ECL发光系统检测相应蛋白质表达水平。
1.6 酶联免疫吸附试验
用涂层缓冲液中的抗原/分析物溶液充分涂覆微量滴定板。盖上培养板,在4℃下孵育过夜。用300 μl洗涤缓冲液洗涤板三次。将阻断缓冲液添加到板中。在37℃孵育1 h。准备分析物抗体混合物用于样品和标准品。将分析物和标准混合物添加到选定的孔中。在37℃孵育1 h。用300 μl洗涤缓冲液洗涤三次。将酶缀合的二抗添加到每个孔中。在37℃孵育1 h。用300 μl洗涤缓冲液洗涤三次。将底物溶液添加到选定的孔中。在室温下孵育直至观察到所需的颜色变化。添加反应终止液。读取吸光度值。
1.7 统计学处理
2 结果
2.1 miR-520a-3p过表达后,野生型和突变型miR-520a-3p的3’UTR的荧光素酶活性
Targetscan在线工具分析结果表明,miR-520a-3p与RELA的3’UTR高度匹配,且匹配区域位于3’UTR的1268-1275位置(图1)。将RELA的3’UTR中与miR-520a-3p相匹配区域中的U均突变为C、C均突变为A并插入pMir-Glo质粒中,此为突变型。与突变型相比,野生型RELA的相对荧光素酶活性下降(100±6vs17±3,P<0.05,图2)。

Fig. 1 miR-520a-3p and NF-κB complex subunit RELA highly matched
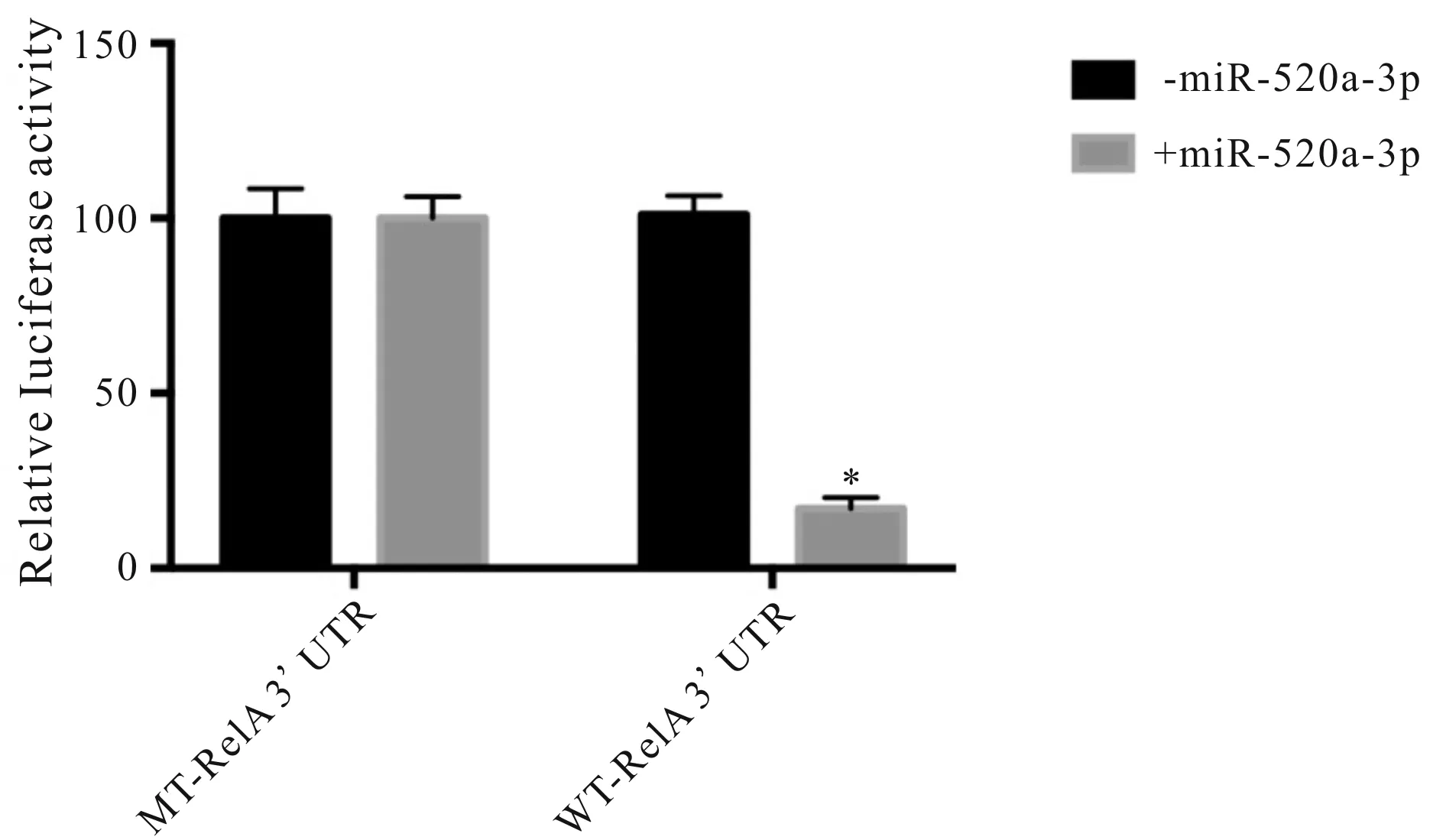
Fig. 2 Luciferase activity of 3 'UTR of wild and mutant miR-520a-3p after overexpression of miR-520a-3p
2.2 过表达miR-520a-3p对宫颈癌细胞HELA因子分泌的影响
在宫颈癌HELA细胞中转染miR-520a-3p mimics后,通过实时荧光定量PCR检测miR-520a-3p水平发现,与对照组相比,过表达miR-520a-3p组的miR-520a-3p基因表达水平上升(1.01±0.10vs3.59±0.66,P<0.05),同时通过免疫印迹检测发现,与对照组相比,过表达miR-520a-3p组的NF-kB复合体亚基RELA的蛋白表达水平下降(图3)。使用LPS激活NF-kB信号通路后发现,与对照组和过表达miR-520a-3p组相比,宫颈癌HELA细胞分泌的细胞因子GCSF, GM-CSF, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12 p40, IL-12 p70, IL-13, IL-17, IFN-γ, MCP-1, MCP-5, RANTES, TNF-α的蛋白表达水平上升(P<0.05),但是LPS处理的同时过表达miR-520a-3p后,相比于LPS处理组,宫颈癌HELA细胞分泌的这些细胞因子蛋白表达水平下降(P<0.05,表1)。

Fig. 3 Expression of the subunit RELA of the NF-kB complex after overexpression of miR-520a-3p
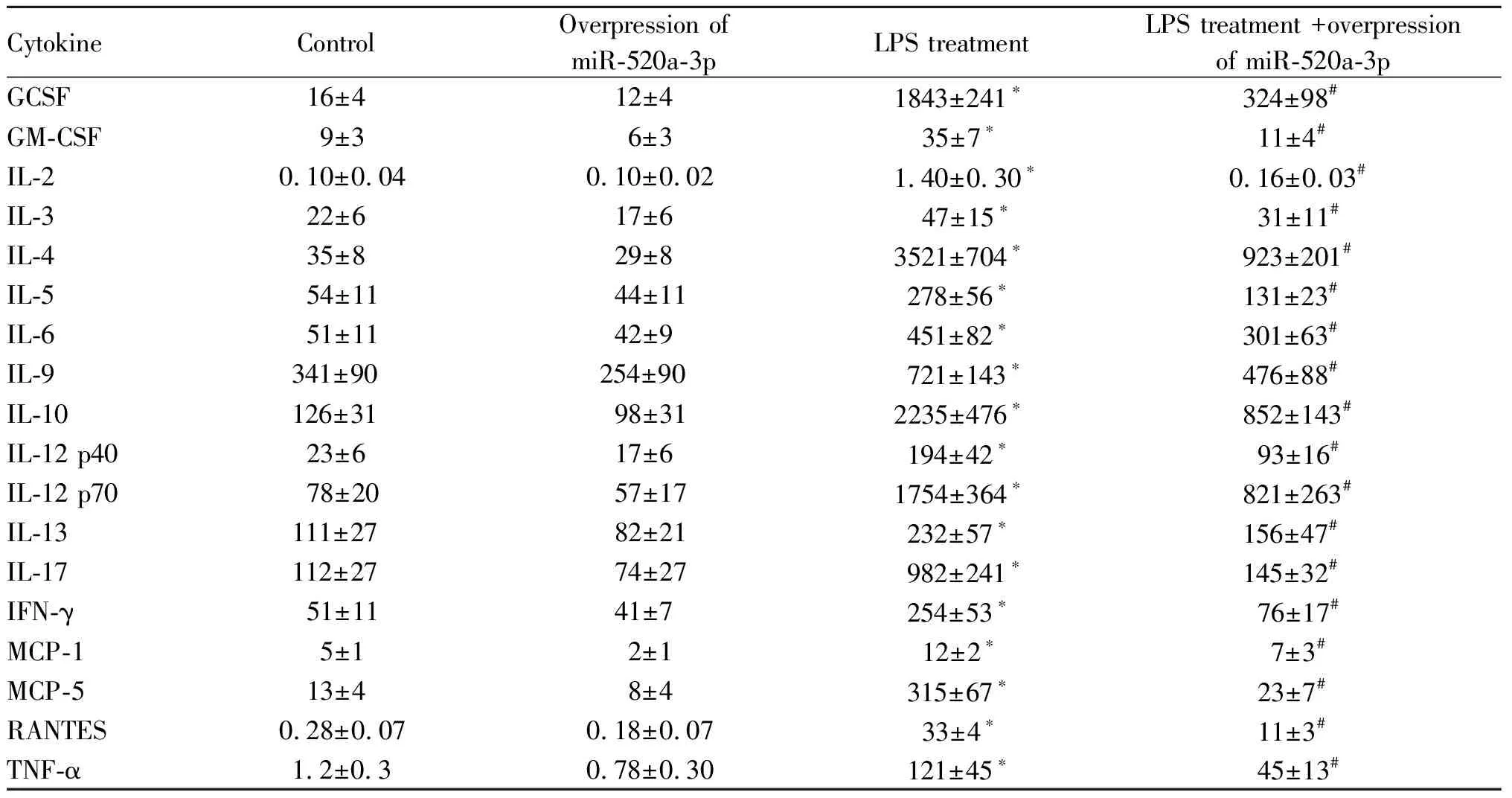
Tab. 1 Effects of overexpression of miR-520a-3p on HELA factor secretion in cervical cancer cells
2.3 敲低miR-520a-3p对宫颈癌细胞HELA因子分泌的影响
在宫颈癌HELA细胞中转染miR-520a-3p inhibitor后,实时荧光定量PCR检测miR-520a-3p水平结果表明,与对照组相比,敲低miR-520a-3p组的miR-520a-3p基因表达水平下降(0.99±0.11vs0.35±0.05,P<0.05),同时免疫印迹检测结果表明,与对照组相比,敲低miR-520a-3p组的NF-kB复合体亚基RELA的蛋白表达水平上升(图4)。使用LPS激活NF-kB信号通路后发现,与对照组和敲低miR-520a-3p组相比,宫颈癌HELA细胞分泌的细胞因子GCSF, GM-CSF, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12 p40, IL-12 p70, IL-13, IL-17, IFN-γ, MCP-1, MCP-5, RANTES, TNFα的蛋白表达水平上升(P<0.05),但是LPS处理的同时敲低miR-520a-3p后,相比于LPS处理组,宫颈癌HELA细胞分泌的这些细胞因子蛋白表达水平进一步上升(P<0.05,表2)。
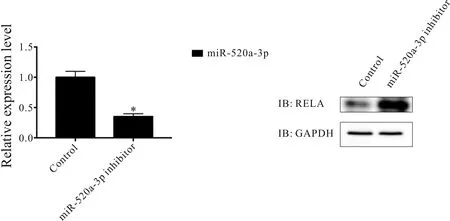
Fig. 4 Expression of subunit RELA in the NF-kB complex after miR-520a-3p knockdown
3 讨论
在本研究中发现,在宫颈癌HELA细胞中miR-520a-3p靶向RELA的3’UTR。Zhang R等人[5]报道miR-520a-3p能够靶向EGFR调控结直肠癌细胞迁移,细胞凋亡。Li J等人[12]发现,miR-520a-3p能够靶向CCND1和CD44调控乳腺癌细胞增殖,迁移和侵袭。因此,miR-520a-3p是一个与多种癌症相关的分子,能够调控癌细胞的生命活动。
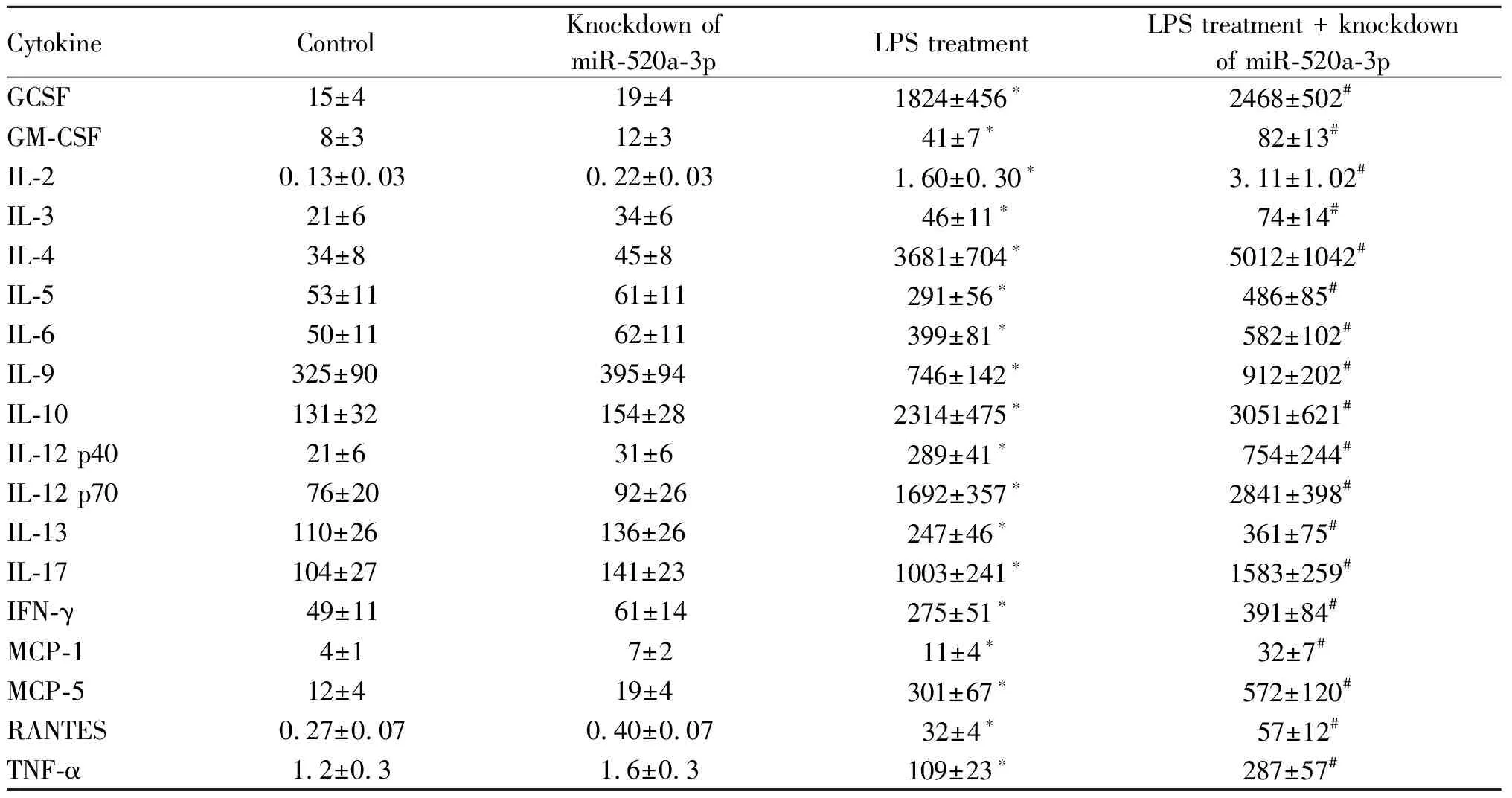
Tab. 2 Effect of miR-520a-3p knockdown on HELA factor secretion in cervical cancer cells
细胞因子也称为白细胞介素,单核因子,淋巴因子,趋化因子和生长因子[13-15]。本研究结果表明,LPS激活NF-kB信号通路后,宫颈癌HELA细胞分泌的细胞因子GCSF, GM-CSF, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12 p40, IL-12 p70, IL-13, IL-17, IFN-γ, MCP-1, MCP-5, RANTES, TNF-α的水平显著上升。NF-κB在大多数细胞中作为p50和p65亚基的同源二聚体或异二聚体复合物存在,并且在与NF-κB抑制蛋白(I-κB)相关的细胞的细胞质中保持无活性[16]。NF-κB被激活以响应各种炎症刺激分泌细胞因子,包括细菌LPS及其通过增加与细胞溶质IκB蛋白减少相关的核p65蛋白诱导的NF-κB活化。在多种疾病中,NF-κB能够调控多种炎症基因的转录,包括TNF-α、IL-1β、IL-2、IL-6、IL-8、IL-12、iNOS、COX2、趋化因子、粘附分子、集落刺激因子,因此NF-κB是治疗各种疾病的重要靶点。
在本研究中,发现过表达miR-520a-3p后,RELA的表达水平显著下降,宫颈癌HELA细胞分泌的细胞因子水平显著下降;敲低miR-520a-3p后,RELA的表达水平上升,宫颈癌HELA细胞分泌的细胞因子水平显著上升。最近的文献揭示一些细胞因子的失调与宫颈癌前病变的发生率,从癌前期到“原位”癌症的进展,进一步侵袭以及末期转移之间的显著关联[17]。最初,细胞因子被认为是免疫系统中的信使分子,将白细胞引导至炎症部位。然而,当失调时,细胞因子与大多数肿瘤组织的发生发展有密切关系,可能在恶性转化,增殖,存活,血管生成,侵袭和转移中起作用。现在已经鉴定近50种细胞因子,其中一些或它们的受体在宫颈癌的癌发生和转移中显著改变[18]。细胞因子如GCSF, GM-CSF, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12 p40, IL-12 p70, IL-13, IL-17, IFN-γ, MCP-1, MCP-5, RANTES, TNF-α等已被证明可作为潜在的生物标志物来评估侵袭性癌症和转移的风险[19]。许多细胞因子在宫颈癌前期和癌症中显著改变,在患有转移的晚期癌症中更是如此。其中一些例如IL-6,IL-17,IL-8与肿瘤生长相关[20],而一些与抑制HPV复制和抑制肿瘤相关,例如RANTES,TNFα,IFNγ。目前,几种miRNA靶向治疗已在临床应用,包括肿瘤抑制因子miRNA miR-34的模拟物,已经进行治疗癌症的I期临床试验,以及针对miR-122的antimiRs,已经进行治疗肝炎的II期临床试验[21]。因此,通过体外合成的miR-520a-3p模拟物可能抑制RELA的表达,抑制宫颈癌细胞因子的分泌,抑制肿瘤的生存、迁移和增殖等过程,延长宫颈癌患者生存率。
