对香豆酸上调BDNF及改善慢性束缚应激诱导小鼠记忆障碍的作用*
2020-11-30肖楚丽谢一铭李金成彭丽文肖志勇于旭东
肖楚丽,谢一铭,李金成,彭丽文,肖志勇,于旭东 △
(1.邵阳学院基础医学院,湖南 邵阳 422000;2.南华大学附属第一医院,湖南 衡阳 421001)
应激是各种神经精神疾病的重要原因。随着生活节奏的不断加快,人类面临着越来越大的生理和心理应激[1]。慢性应激一方面可导致各种精神疾病如抑郁、焦虑,另一方面可导致神经退行性疾病如认知障碍[2,3]。虽然越来越多的人面临应激性记忆障碍的困扰,然而应激诱导记忆障碍的神经机制仍不清楚。
脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)及其原肌球蛋白激酶B(tyrosine kinase receptor B, TrkB)受体广泛分布于哺乳动物大脑中。越来越多的证据表明,BDNF/TrkB信号在神经可塑性和记忆形成中起着重要作用[4-7]。此外,BDNF/TrkB还影响与应激相关的记忆表现[8],如慢性应激导致BDNF表达降低,而药物发挥抗应激同时伴随BDNF的上调[9]。BDNF的作用主要通过TrkB介导。ANA-12是一种广泛应用的TrKB受体拮抗剂,可选择性直接与TrkB受体结合,抑制TrkB下游通路。其可阻断 BNDF与TrkB受体的结合从而降低BDNF的作用。如,ANA-12可防止脑源性神经营养因子(BDNF)介导的神经突增生;ANA-12可消除BDNF增加内向电流的作用[10]。
对香豆酸(p-Coumaric acid, p-CA)是一种衍生自芳香族氨基酸的苯丙酸[11],具有抗氧化和神经保护作用。最近研究表明,p-CA可上调BDNF并且改善抑郁样行为[12]。然而p-CA在学习记忆中的作用却研究滞缓,尤其是在应激性记忆障碍的作用并无报道。本文基于慢性束缚应激(chronic restraint stress , CRS)诱导的记忆障碍模型,探索p-CA对应激性记忆障碍的影响。此外,我们应用TrKB受体拮抗剂ANA-12观察其对p-CA促记忆作用的影响,以期明确BDNF/TrkB信号是否介导p-CA的这种作用。
1 材料与方法
1.1 实验动物
ICR小鼠(雄性,9周龄)购于湖南斯莱克有限公司,小鼠配对饲养在标准的环境下(温度为(25±1)℃,湿度为(50±5)%, 昼夜周期为12 h,其中8:00-20:00为明),小鼠自由饮水,摄食。实验开始前实验者抚摸小鼠7 d,每天3 min。所有实验程序均完全按照美国国家卫生研究院关于和使用实验室动物的指南进行。
1.2 实验仪器与试剂
Y型迷宫、新颖物体识别箱及其视频采集系统均购于北京众实迪创科技有限公司。BDNF 抗体、β-actin抗体均购于美国proteintech。基于之前的研究[12],p-CA(阿拉丁,上海)分散于10%的吐温80中,剂量为100 mg/kg;ANA-12(selleck,美国)分散于1% DMSO中,剂量为0.5 mg/kg[9];以上药物均经腹腔注射,药物及溶媒的注射体积为10 ml/kg。
1.3 实验设计
(1)p-CA对CRS应激小鼠学习记忆障碍的影响。小鼠随机分成对照组(Control),慢性束缚应激组(CRS)和慢性束缚应激+p-CA组(CRS+p-CA),每组8只,其中慢性束缚应激小鼠每天接受4 h的束缚应激,连续束缚10 d,而对照组小鼠留在笼中不被打扰。第11日小鼠腹腔注射溶媒(10%吐温80)或p-CA(100 mg/kg),注射后2 h进行Y迷宫测试,注射后24 h进行新颖物体识别采样,采样后2 h进行新颖物体识别测试。实验结束后断头取脑剥离海马并检测BDNF蛋白表达。
(2)ANA-12对p-CA改善CRS应激小鼠记忆障碍的影响。小鼠随机分成慢性束缚应激组(CRS),慢性束缚应激+ANA-12组(CRS+ANA-12)和慢性束缚应激+p-CA组(CRS+ p-CA)慢性束缚应激+p-CA +ANA-12组(CRS+ p-CA +ANA-12)。每组8只,4组小鼠每天接受4 h的束缚应激,连续束缚10 d。第11日小鼠腹腔注射溶媒(10%吐温80)或p-CA(100 mg/kg),ANA-12在p-CA注射前30 min给药。注射后2 h进行Y迷宫测试,注射后24 h进行新颖物体识别采样,采样后2 h进行新颖物体识别测试。
1.4 慢性束缚应激(CRS)
小鼠被放入50 ml的离心管,离心管均匀分布直径为4 mm的透气孔,每天上午10:00开始束缚,每天束缚4 h,连续束缚10 d。
1.5 新颖物体识别测试(NOR)
新颖物体识别测试分为采样和测试两个阶段,在采样阶段,小鼠自由探索两个物体5 min。采样结束后2 h进行测试阶段,采样阶段的物体被随机替换一个新的识别物体。小鼠自由探索两个物体5 min。小鼠行为通过箱顶的摄像头记录并由熟练者双盲分析。小鼠的鼻子距物体小于1 cm则视为探索。辨别指数(DI)=(新物体的探索时间-旧物体的探索时间)/(新物体的探索时间+旧物体的探索时间)。
1.6 Y迷宫测试
小鼠依次放入Y型迷宫(臂长40 cm,高15 cm)中5 min,迷宫上方摄像头记录小鼠行为。熟练者双盲统计小鼠5 min内入臂的次序,并计算出正确交替率。一次交替定义为小鼠连续的3 次进入三个不同的臂,中间无重复进入。最大交替次数为总入臂次数减去2,正确交替率为实际交替次数除以最大交替次数。
1.7 Western blot 检测BDNF蛋白表达
取海马组织,提取蛋白并使用BCA蛋白定量试剂盒测定蛋白浓度。电泳分离蛋白质,湿转法转膜,用1倍TBST配制5%脱脂奶粉,将膜浸入后,室温放置1.5 h。分别加入BDNF抗体(滴度1∶300)及β-actin 抗体(滴度1∶5 000),4℃孵育过夜。再加入辣根过氧化酶标记的二抗孵育2 h。用TBST 充分漂洗并与ECL化学发光试剂反应。曝光后扫描,并用quantity one专业灰度分析软件进行分析。
1.8 统计学处理
2 结果
2.1 对香豆酸改善慢性束缚应激诱导小鼠记忆障碍
2.1.1 Y迷宫 在Y迷宫测试中,与Control组相比,各组小鼠交替次数无显著性差异(P>0.05), 而CRS组小鼠正确交替率显著降低(P<0.05);与CRS组相比,CRS+ p-CA组小鼠正确交替率显著升高(P<0.05,表1);这些结果表明p-CA改善CRS应激诱导小鼠Y迷宫记忆障碍。
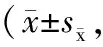
Tab. 1 Effect of p-CA on memory deficit of Y-maze test induced by CRS in mice n=8)
2.1.2 新颖物体识别(NOR)测试 在NOR测试中,与Control组相比,各组小鼠在NOR采样阶段对两物体的总探索时间和测试阶段穿越格子数均无显著性差异(P均>0.05)。而CRS组小鼠在测试阶段的辨别指数显著降低(P<0.05);与CRS组相比,CRS+ p-CA组小鼠辨别指数显著升高(P< 0.05,表2);这些结果表明p-CA改善CRS应激诱导小鼠NOR记忆障碍。
Tab. 2 Effect of p-CA on memory deficit of NOR test induced by CRS in mice n=8)
2.1.3 BDNF蛋白表达 Western blot结果显示,与Control组相比,CRS组小鼠海马内BDNF蛋白表达显著降低(P< 0.01);与CRS组相比,CRS+ p-CA组小鼠海马内BDNF蛋白表达显著升高(P<0.01,图1)。这些结果表明,p-CA上调CRS诱导小鼠海马BDNF蛋白表达降低。
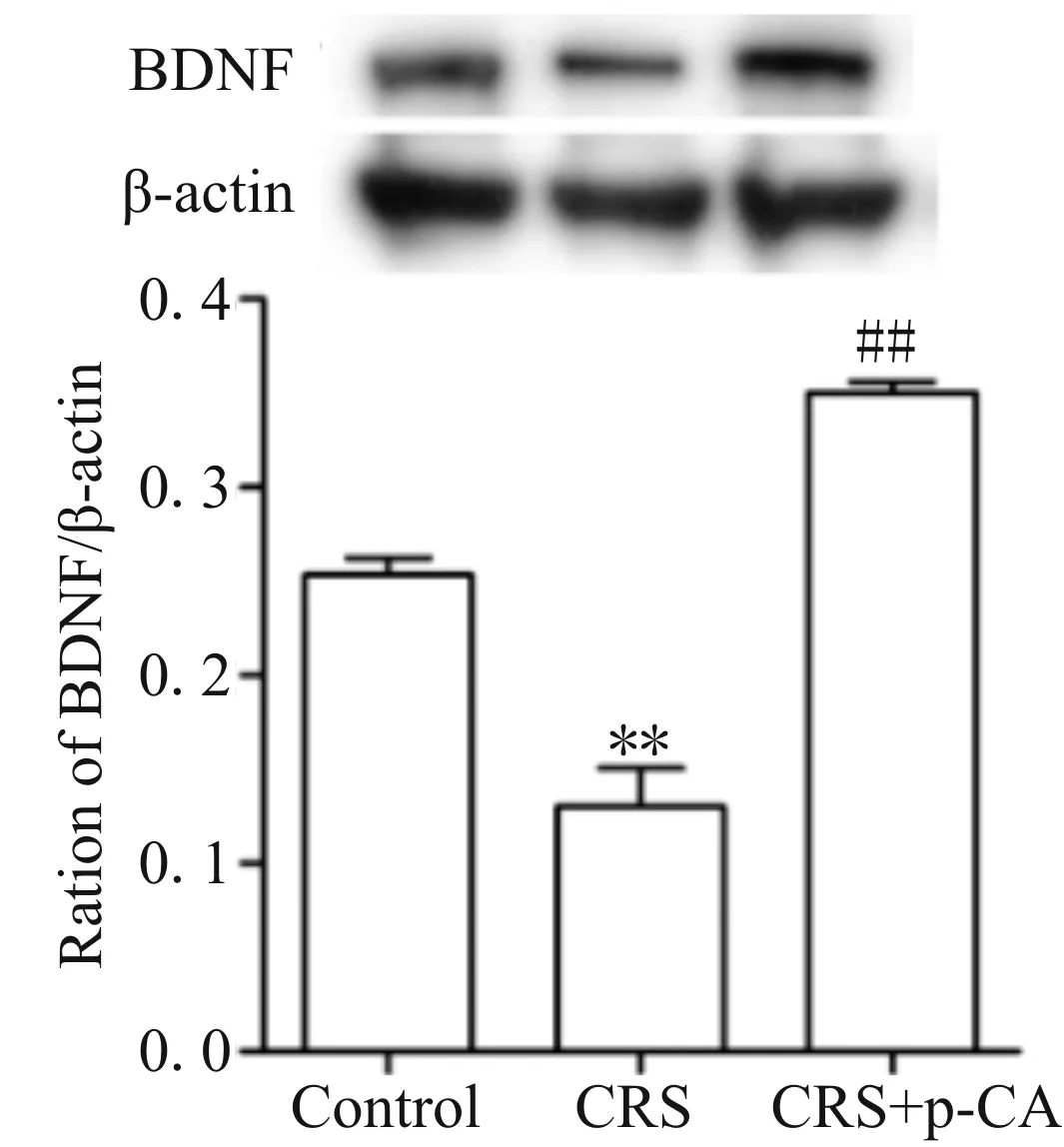
Fig. 1 p-CA reversed CRS-induced reduction of BDNF protein level n=3)
2.2 ANA-12阻断对香豆酸对慢性束缚应激诱导小鼠记忆障碍的改善作用
2.2.1 Y迷宫 在Y迷宫测试中,与CRS组相比,各组小鼠交替次数无显著性差异(P>0.05), 而CRS+ p-CA组小鼠正确交替率显著升高(P<0.05);与CRS+ p-CA组相比,CRS+ p-CA+ ANA-12组小鼠正确交替率显著降低(P<0.05,表3);这些结果表明ANA-12阻断p-CA对CRS应激诱导小鼠Y迷宫记忆障碍的改善作用。
Tab. 3 Blocking effects of ANA-12 on p-CA improved memory deficit of Y-maze test in CRS mice n=8)
2.2.2 新颖物体识别测试 在NOR测试中,与CRS组相比,各组小鼠在NOR采样阶段对两物体的总探索时间和测试阶段穿越格子数均无显著性差异(P均>0.05)。而CRS+ p-CA组小鼠在测试阶段的辨别指数显著升高(P<0.05);与CRS+ p-CA组相比,CRS+p-CA+ANA-12组小鼠辨别指数显著降低(P<0.05,表4);这些结果表明ANA-12阻断p-CA对CRS应激诱导小鼠NOR记忆障碍的改善作用。
Tab. 4 Blocking effects of ANA-12 on p-CA improved memory deficit of NOR test in CRS mice n=8)
3 讨论
当前研究发现P-CA可升高CRS小鼠Y迷宫测试中小鼠正确交替率以及NOR测试中小鼠辨别指数。此外,考虑到海马BDNF信号在学习记忆中的作用,本文检测BDNF在海马中的表达及TrkB受体拮抗剂ANA-12对P-CA改善CRS小鼠记忆障碍的作用。结果显示,P-CA改善CRS小鼠BDNF表达降低,并且ANA-12阻断P-CA对慢性束缚应激小鼠记忆障碍的改善作用。这些结果表明,P-CA改善慢性束缚应激诱导小鼠记忆障碍,并且这种作用依赖于BDNF的上调。
NOR测试利用动物对新物体的天然偏好来检测动物的识别记忆,Y型迷宫常用于检测动物的空间记忆。在慢性不可预测应激,慢性束缚应激均导致小鼠NOR与Y迷宫中小鼠学习记忆降低[13-15]。与这些发现一致,当前研究表明CRS导致小鼠NOR与Y迷宫中学习记忆障碍。P-CA是白藜芦醇和柚皮素等不同类黄酮的前体,在多种中枢疾病中发挥神经保护作用。在脑缺血模型中,P-CA可改善小鼠脑缺血/再灌注损伤[16]。在抑郁模型中,50 mg/kg,75 mg/kg和100 mg/kg的P-CA经腹腔一次注射均可改善LPS诱导的大鼠强迫游泳测试及悬尾测试中的抑郁样行为以及减轻炎症反应[11]。药代动力学上,p-CA在肠道中被快速吸收,与其他酚类化合物相比具有更高的生物利用度[17],此外,我们未公开的结果显示一次腹腔注射100 mg/kg的P-CA 30 min后可改善小鼠抑郁样行为。基于以上研究基础,本研究采用100 mg/kg剂量的P-CA经腹腔注射一次后2 h进行记忆测试。结果显示P-CA可改善CRS诱导小鼠NOR与Y迷宫中学习记忆障碍,这与P-CA改善东莨菪碱诱导大鼠被动回避测试和水迷宫测试中记忆障碍中的结果一致[18]。进一步支持P-CA的神经保护作用和扩展P-CA的促学习记忆作用。为排除活动度对实验结果的影响,在Y迷宫及NOR测试中分别检测小鼠的总交替次数和穿越格子数,结果显示p-CA不影响小鼠的活动水平,表明p-CA改善CRS诱导小鼠记忆障碍不是通过影响小鼠的活动水平。
BDNF通过与TrkB 受体结合在学习记忆中起到重要作用。例如,最近的一项研究表明BDNF杂合突变小鼠在NOR和Y迷宫中记忆障碍[19]。慢性应激可导致海马BDNF水平降低,而抗应激药物可上调BDNF并且改善应激诱导记忆障碍[13,20]。此外,BDNF抗体和TrkB信号拮抗剂均可阻断药物对应激诱导记忆障碍的改善作用[9,13]。与这些研究一致,当前研究表明慢性束缚应激诱导BDNF表达降低,而p-CA上调BDNF并且改善应激诱导记忆障碍。ANA-12是一种广泛应用的TrKB受体拮抗剂,其可阻断 BNDF与TrkB受体的结合从而降低BDNF的作用。如ANA-12可阻断Apelin-13上调BDNF后对应激性记忆障碍的改善作用[13]。一致的是,当前研究表明ANA-12阻断p-CA对慢性束缚小鼠学习记忆降低的改善作用,提示BDNF/TrkB信号参与p-CA的对应激小鼠学习记忆障碍的改善作用。然而,为进一步明确BDNF的作用,应在今后的研究中,中枢注射BDNF抗体探索其能否阻断p-CA对应激小鼠学习记忆障碍中的改善作用。
综上,本研究基于CRS模型,观察到p-CA可逆转CRS应激诱导认知障碍,这一结果可能对应激性记忆障碍的药物研发具有潜在价值;并且p-CA的这种作用可能依赖于BDNF蛋白表达的上调,但其更确切的机制仍需要在今后的研究中进一步探索。
