脂质氧化对肉色影响的研究进展
2020-11-27刘文轩杨啸吟张一敏朱立贤毛衍伟梁荣蓉韩明山成海健
刘文轩,罗 欣,杨啸吟,张一敏,朱立贤,毛衍伟,梁荣蓉,*,韩明山,成海健
(1.山东农业大学食品科学与工程学院,山东 泰安 271018;2.国家肉牛牦牛产业技术体系通辽站,内蒙古 通辽 028100;3.国家肉牛牦牛产业技术体系济南站,山东 济南 250000)
肉色是消费者判断生鲜肉是否新鲜和决定购买与否的重要指标[1],消费者往往认为明亮的樱桃红色和真空包装的紫红色是肉制品新鲜的标志[2]。pH值、脂质氧化、包装方式、加热方式及生产过程中的不同处理等都会影响肌红蛋白的状态,从而影响肉色[3]。脂质氧化是影响肉色的重要因素,研究表明,脂质氧化会产生醛、酮等次级代谢产物,这些次级代谢产物能够促进肌红蛋白的氧化,给肉色带来不良影响[4-6]。该过程不但会影响生鲜肉的色泽,还会影响肉熟制后的色泽。肌红蛋白氧化后形成的高铁肌红蛋白热敏性高,容易造成肉制品熟制过程中的提前褐变(premature browning,PMB)现象[7],即加热的中心温度还未到达完全熟制的中心温度时,肉色提前产生熟肉色泽的现象。该现象容易使消费者产生错误的判断,导致购买并食用热杀菌不彻底的肉制品,带来食品安全问题。有研究表明,当气调包装体系中的氧气含量升高时,牛肉饼的脂质氧化程度加剧,PMB现象更明显[8]。
脂质氧化促进肌红蛋白氧化的主要原因是其产生的自由基等初级氧化产物和醛、酮等次级氧化产物能够促进肌红蛋白氧化。目前,关于脂质氧化初级代谢产物对肉色的影响机制较为明确,而次级代谢产物以及脂肪酸种类等对肉色的影响尚不完全清楚[6-7]。本文综述了脂质氧化途径及其与肌红蛋白氧化之间的关系,阐述了脂质氧化对牛肉肉色的影响及可能的机制,并综述了通过控制脂质氧化来改善牛肉肉色的措施及最新相关研究进展。旨在为脂质氧化在控制肉制品色泽中的应用提供理论参考。
1 肉中脂质氧化机制与途径
肉与肉制品是营养丰富的食品,含有丰富的蛋白质、脂肪、脂溶性维生素、矿物质和生物活性物质等。这些复杂的营养组分容易导致肉与肉制品在加工和贮藏过程中发生一系列的化学反应和微生物污染,最终导致产品品质劣变。而脂质氧化是这一过程中导致肉品劣变的最主要因素之一[9-10]。脂质的氧化最早开始于动物屠宰之前,屠宰后随着环境的改变和肉制品内部抗氧化能力的减弱,脂质氧化程度进一步增强。肉中脂质的氧化反应过程复杂,具有自发性,且不可避免。诸多因素参与并影响着脂质氧化的进程,包括宰前的饲养状况、营养水平、屠宰管理和宰后处理过程以及熟制过程等。肌肉内部的铁离子、肌红蛋白分子和双氧水等物质可以作为催化剂或者促进剂,促进活性氧的产生从而引起脂质氧化;而辐照或者光照等一些外界物理因素也可以引起脂质的氧化反应;此外,动物体内的脂肪氧合酶(lipoxygenase,LOX)也能加快氧气与不饱和脂肪酸反应的过程。因此,在宰后的生化体系中,脂质氧化一般通过自动氧化、酶促氧化和光氧化3 种反应途径进行[9],如图1[11]所示。
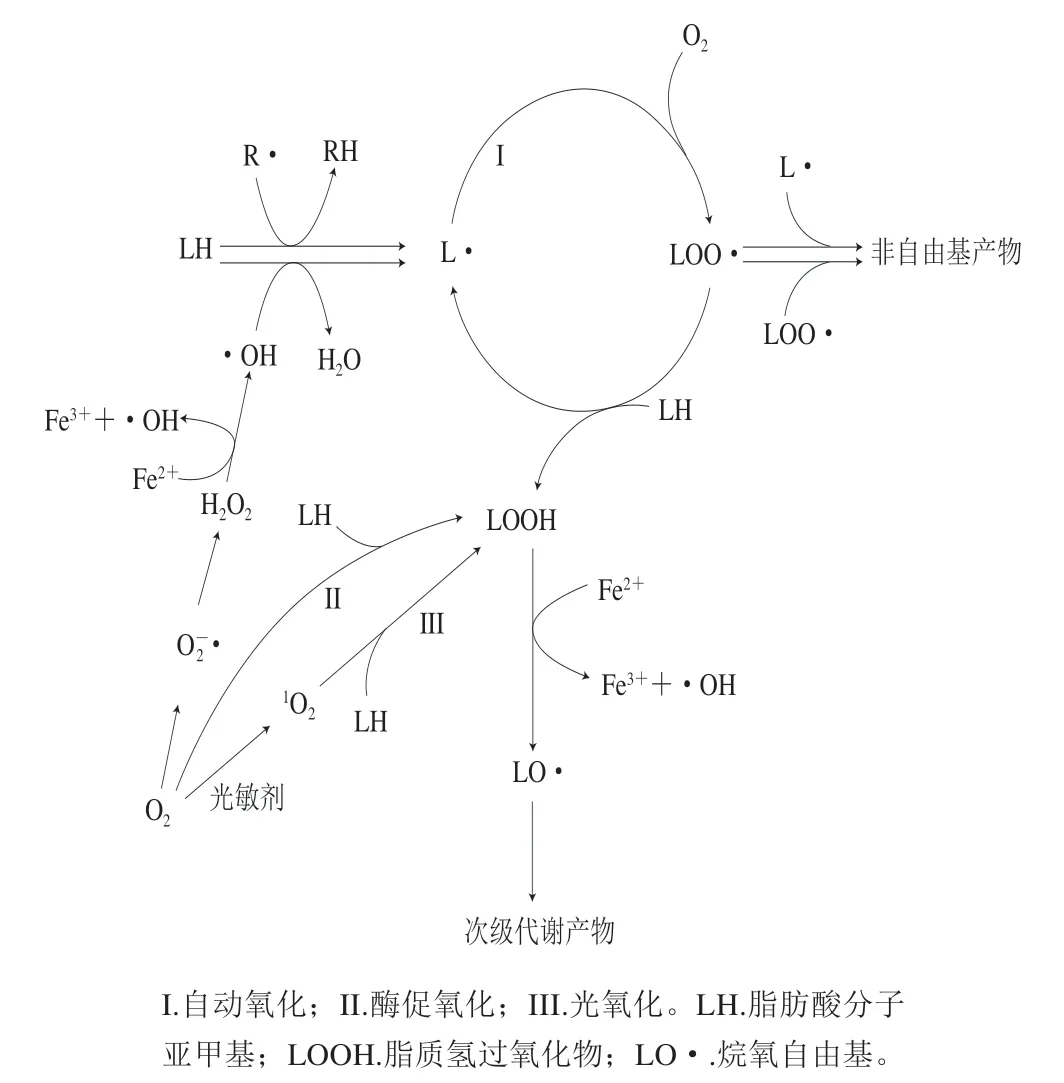
图1 脂质氧化的3 种途径[11]Fig. 1 Three pathways of lipid oxidation[11]
1.1 自动氧化
自动氧化是一个复杂的过程,是肉制品中最主要的氧化方式[12],包括诱导期、传递期和终止期3 个阶段[13]。它是由各种自由基攻击脂质底物所引起,受到自由基种类、金属离子、环境温度、脂质底物组成等多种复杂因素影响。该过程首先是一个自由基与脂肪酸烃链发生反应生成一系列过氧化物,这些过氧化物又进一步与其他脂肪酸烃链反应夺取其氢原子形成氢过氧化物,而被夺取氢原子的碳链则作为新的过氧化物进一步参与新的循环[9](图1)。该链式传播过程在终止以前大约能发生100 次以上。该过程中产生的氢过氧化物不稳定,最终降解为醛、酮等小分子化合物[14]。脂质氧化在诱导期、传递期产生初级代谢产物(如烷基、烷氧基、氧自由基),它们容易从相邻分子中夺取氢原子,形成的过氧化物经过分裂可以形成低分子质量的次级代谢产物,最具代表性的是4-羟基壬烯醛(4-hydroxynonenal,HNE)和丙二醛(malondialdehyde,MDA)。许多研究已证明这两种氧化产物能够促进肌红蛋白的氧化,造成肉色劣变[15-17]。同时,在医学领域已有研究表明,这一过程产生的自由基能够对线粒体膜造成损伤[18],而线粒体功能对肉色稳定性具有重要影响[19],因此,脂质氧化也会通过对线粒体膜的伤害对肉色造成不良影响。
动物屠宰以后,在肌肉向食用肉转变的过程中,随着各类生化机制的变化,机体的抗氧化能力显著下降,脂质的氧化反应开始加剧。同时,宰后pH值的降低也会加快脂质氧化的进程,因为H+会促进肌红蛋白分子的还原,加快其对脂质氧化的促进作用[9]。因此,随着宰后时间的延长,肉中脂质氧化逐渐加剧,其对肉色影响也更加显著。
1.2 酶促氧化
酶促氧化是动物体内的LOX加速氧气与不饱和脂肪酸反应的过程[20],氢过氧化物极不稳定,进一步反应能够形成一些低级脂肪酸、醛、酮等物质,是脂质氧化过程的一种重要途径。LOX存在于不同动物的肌肉组织中,它可以直接氧化不饱和脂肪酸形成脂质过氧化物,因此,LOX能参与动物宰后肉中的脂质氧化[21]。宰后LOX能够加速脂质的次级氧化,产生许多挥发性的风味物质[22]。LOX分子中间有一个非血红素铁,通常以Fe2+形式存在,此时LOX处于非激活状态,当LOX被脂肪酸氢过氧化物氧化,中间铁原子由+2价变为+3价时,LOX被激活[23]。激活态的LOX通过氧化多不饱和脂肪酸,使多不饱和脂肪酸亚甲基上的氢断裂,随后被氧化为含有共轭双键的氢过氧化物,最终生成一系列自由基和过氧化氢,从而影响肉色和肉色稳定性。酶促氧化的反应过程主要受LOX活性的影响,虽然是脂质氧化的一个重要途径,但是在肉制品的脂质氧化过程中不占主导优势[12]。
1.3 光氧化
光氧化是不饱和双键与单线态氧直接发生的氧化反应。肌肉中的铁卟啉、肌红蛋白和核黄素作为光敏剂[24]受到光照后将基态氧(3O2)转变为激发态氧(1O2)[25]。这种激发态氧具有较高的亲电性,使光氧化速率比自动氧化速率更快。史智佳等以金枪鱼为研究对象,探究了不同单色光照对脂质氧化的影响,结果表明,与暗室贮藏相比,单色光照能够显著增加金枪鱼脂质氧化程度,并且蓝色光照比红色光照导致金枪鱼脂质氧化的程度更大,这是因为蓝色光能量更高,更能促进单线态氧的产生,进而促进脂质氧化的发生[26]。
2 脂质氧化对肉色的影响
在众多影响肉色稳定性的因素中,脂质氧化程度与肉色稳定性密切相关,脂质氧化程度越低,肉色稳定性越高,反之,脂质氧化程度越高,肉色稳定性越低[27]。Inserra等探究了以柑橘干果肉替代部分大麦饲喂羔羊对羊肉脂质氧化的影响,结果表明,与对照组相比,添加柑橘干果肉的羔羊肉在有氧贮藏6 d后脂质氧化程度显著降低,肉色稳定性同时得到提高[28]。由此可见,降低肉制品的脂质氧化程度有利于肉色稳定性的提高。Wongwichian等研究了大眼竹荚鱼在冷藏过程中肌红蛋白氧化、脂质氧化与肉色的关系,结果表明,随着贮藏时间的延长,大眼竹荚鱼的硫代巴比妥酸(thiobarbituric acid reactive substances,TBARS)含量与高铁肌红蛋白含量显著增加,相对红度显著下降,脂质氧化与肉色稳定性显著相关[29]。
脂质氧化对肉色的影响机制主要有3 个途径。较为普遍的解释为脂质氧化产生的初级或次级代谢产物能促进肌红蛋白氧化,而肉色与肌红蛋白的状态密切相关;脂质氧化对线粒体活性的影响也是影响肉色稳定性的一个重要因素;此外,脂质氧化与肌红蛋白争夺体系内的氧气也是影响该过程的一个重要方面。
2.1 脂质氧化的初级和次级代谢产物对肌红蛋白氧化的影响
2.1.1 脂质氧化初级代谢产物对肉色的影响
脂质氧化的初级代谢产物是一系列自由基,这些自由基能够使肌红蛋白中的铁元素发生氧化反应[30]。肌红蛋白辅基中Fe2+被脂质氧化产生的自由基夺取电子形成Fe3+,从而加速了高铁肌红蛋白的形成。
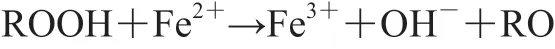
动物屠宰以后,肌肉细胞处于缺氧的状态,肌红蛋白中的Fe2+与肌肉中原有的H2O2发生Fenton反应[31]。该反应会加速肌红蛋白中的Fe2+向Fe3+的转变,从而导致肌红蛋白向高铁肌红蛋白的转变。

胴体分割以后,肉表面的肌肉细胞暴露在空气中,氧气逐渐渗入组织内部,组织内氧气含量逐渐升高,当组织中O2与H2O2的浓度比值大于1 000时,血红素铁与氧气形成铁氧络合物,通常以Fe2+-O2和Fe2+-O的形式存在,发生如下反应[13]。

这一反应过程产生的铁氧络合物Fe2+-O2、Fe2+-O和·是脂质氧化的引发剂,另外,肌红蛋白中游离的血红素铁也是脂质氧化的催化剂。由此可见,肌红蛋白中的铁元素可以促进脂质氧化,而脂质氧化又能促进铁元素的氧化,二者的氧化作用相互促进。这就导致了肉色稳定性的下降,加速肉品失色褐变[31]。Zhou Feibai等在猪背最长肌匀浆液中添加0~0.3 mmol Fe2+,探究Fe2+对脂质氧化和蛋白氧化的影响,分别在添加Fe2+前测定和在添加后80、200 min测定其氧合肌红蛋白、高铁肌红蛋白相对含量以及脂质氧化程度,与未添加Fe2+相比,添加Fe2+能够显著增加匀浆液的脂质氧化程度,添加0.3 mmol Fe2+孵育200 min时脂质氧化程度最高,此时高铁肌红蛋白相对含量显著升高至85%左右[32]。
2.1.2 脂质氧化次级代谢产物对肉色的影响脂质氧化产生的次级代谢产物主要以醛、酮为主,其中HNE是最活跃的物质之一[33]。HNE一方面能够与氧合肌红蛋白结合,促进氧合肌红蛋白的氧化;另一方面能够抑制高铁肌红蛋白还原酶的活性,从而抑制高铁肌红蛋白的还原[4]。Lynch等研究了脂质氧化产生的醛类化合物对肌红蛋白的影响,结果表明,与对照组相比,HNE能够显著加快氧合肌红蛋白的氧化速率,在孵育90 min后氧合肌红蛋白相对含量降低至65%左右,并且HNE能够促进亚铁血红素的释放,从而抑制高铁肌红蛋白的还原,高铁肌红蛋白的积累造成了肉色褐变[4]。Yin Shuang等研究了HNE对绵羊肌红蛋白稳定性的影响,得出与文献[4]相同的结论,并且在添加抑制脂质氧化次级代谢产物的抗氧化剂α-生育酚的微粒体模型中,发现HNE对肌红蛋白的氧化还原稳定性没有影响,这说明添加α-生育酚能抑制脂质氧化次级代谢产物对肌红蛋白的攻击,有利于绵羊肉色稳定性的保持[16],这也进一步说明了HNE是导致肌红蛋白氧化的一个重要物质。
MDA是脂质氧化的另一种典型的次级代谢产物,它分子质量小,但与肌红蛋白反应活性较强,能促进肌红蛋白的氧化。另外,MDA也能够抑制线粒体活性,使高铁肌红蛋白的还原能力降低[34]。Wang Zhaoming等研究了MDA对兔肉肌红蛋白氧化的影响,发现MDA能够促进高铁肌红蛋白的形成,1 mmol MDA孵育8 h后,高铁肌红蛋白的相对含量达到60%左右,导致肉品的褐变失色[17]。
肉在贮藏加热过程中,脂质不断氧化,除了HNE和MDA外,还产生许多其他的二级氧化产物,如α-和β-不饱和醛等,这些物质是否会促进肌红蛋白的氧化还需要进一步的研究。此外,脂质氧化次级代谢产物除了醛类物质外,还有酮类、酸类物质以及醇类物质,这些不同于醛类物质的次级代谢产物是否也会对肉色造成不良影响,目前尚不明确。因此,脂质氧化对肉色的影响仍然是值得探究的问题。
2.2 脂质氧化对线粒体活性的影响
近年来,人们在线粒体活性对肉色的影响方面已进行了大量研究[19]。线粒体主要通过两种机制来影响肉色:1)线粒体呼吸作用消耗氧气,使肌红蛋白保持脱氧肌红蛋白的状态,而脱氧肌红蛋白易被氧化为高铁肌红蛋白,造成肉色劣变;2)肌肉的高铁肌红蛋白还原能力与线粒体活性密切相关,线粒体活性的降低会导致高铁肌红蛋白还原能力的下降,从而造成高铁肌红蛋白的积累[35]。线粒体膜是磷脂双分子层结构,易被自由基和活性氧攻击而受到破坏,造成线粒体的氧化损伤,使线粒体对高铁肌红蛋白的还原能力减弱,对肉色稳定性造成不良影响[36],因此,线粒体膜是脂质氧化的一个重点部位,也是人们研究肉色稳定性的一个重要出发点。
大部分研究认为,脂质氧化会对线粒体活性造成不良影响,主要表现为线粒体膜易发生脂质氧化,导致其通透性下降,从而导致其生物活性的丧失。Chen Cheng等提取牛背最长肌线粒体,研究脂质代谢产物对线粒体膜通透性的影响,发现与对照组相比,添加脂质代谢产物HNE和己烯醛的线粒体悬浊液孵育1 h后,在520 nm波长处的吸光度分别显著下降0.037和0.039[6]。Halestrap研究表明线粒体膜的通透性与A520nm呈负相关,这说明脂质代谢产物能够显著改变线粒体膜的通透性,对肉色稳定性造成不良影响[37]。Chen Cheng等发现HNE和己烯醛不仅能够加速氧合肌红蛋白向高铁肌红蛋白的氧化,同时还能够使线粒体膜发生损伤,抑制线粒体介导的高铁肌红蛋白还原,对肉色稳定性造成不良影响[6]。另外,MDA也能够抑制线粒体活性,使高铁肌红蛋白的还原能力降低[34]。Long Jiangang等研究了MDA对大鼠离体肝细胞线粒体活性的影响,结果表明,MDA能够抑制线粒体膜上的烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)-泛醌还原酶和琥珀酸-泛醌还原酶的活性,造成线粒体氧化呼吸链功能的破坏[34]。
2.3 脂质氧化过程通过争夺体系内的氧气对肌红蛋白氧化的影响
目前已有大量的研究表明脂质氧化的代谢产物会对肉色稳定性造成不良影响。脂质氧化的次级代谢产物HNE、MDA也被逐步证明会降低肉色稳定性。然而,肌红蛋白的氧化需要氧气的参与,而脂质氧化也是一个耗氧反应,因此,许多学者对脂质氧化消耗溶解氧与脂质氧化代谢消耗氧哪种方式对肌红蛋白氧化影响程度更大展开了研究。
O’Grady等在敞口容器中以牛背最长肌匀浆液为研究对象,发现脂质氧化的初级代谢产物能够促进肌红蛋白氧化,影响肉色稳定性,而次级代谢产物HNE、MDA等对肌红蛋白氧化没有显著影响,这与之前的诸多研究结果不一致,可能是因为脂质氧化是一个耗氧的反应,同一个体系条件下,它会争夺体系内的氧气,使体系内的氧气浓度降低,而低氧气浓度能够促进高铁肌红蛋白的形成[38],使敞口容器中脂质氧化消耗的氧气得到补充,匀浆液中始终维持较高的溶解氧水平,因此,肌红蛋白氧化程度低[39]。Monahan等以牛背最长肌匀浆液为研究对象,分别进行有氧搅拌、无氧搅拌以及不进行处理,结果表明,有氧搅拌中脂质氧化程度高,但随着搅拌的进行,体系中的氧气不断得到补充,导致肌红蛋白氧化程度低;无氧搅拌中脂质氧化程度低,但肌红蛋白氧化程度高,这是由于体系中氧气含量低,促进了高铁肌红蛋白的形成[40]。以上研究都表明了脂质氧化消耗溶解氧的程度对肌红蛋白氧化的影响大于脂质氧化代谢产物对肌红蛋白氧化的影响。而肌肉组织中也会存在争夺氧的现象,但是否也存在脂质氧化消耗溶解氧对肌红蛋白氧化程度影响更大的现象,还需要进一步的研究证实。
3 影响脂质氧化的因素
脂质氧化过程复杂,受很多因素的影响,这些因素可以归纳为内部因素和外部因素[20]。内部因素包括动物脂肪含量及脂肪酸组成、LOX活性、金属元素、蛋白氧化及肌红蛋白氧化、pH值等。外部因素包括贮藏温度、包装方式、盐类的添加等[9-10,41]。
3.1 内部因素
3.1.1 动物脂肪含量及脂肪酸组成
肉制品中的脂肪酸含量及组成是影响脂质氧化稳定性的重要因素之一。一般来说,饱和脂肪酸的氧化稳定性高,脂质氧化一般是从不饱和脂肪酸开始的,脂肪酸分子的不饱和程度越高,越容易发生氧化[42]。有研究表明,非反刍动物的肉中含有更高含量的不饱和脂肪酸,比反刍动物更易发生氧化[43]。刘永祥等以肉鸡为研究对象,探究饲料中添加共轭亚油酸和鱼油对脂肪酸组成和脂质氧化状态的影响,结果表明,添加共轭亚油酸有利于C16:0和C18:0等饱和脂肪酸含量的增加,脂质氧化水平显著降低,而添加鱼油组中n-3多不饱和脂肪酸含量显著升高,脂质氧化稳定性显著降低。这说明饱和脂肪酸含量越高,脂质氧化稳定性越高[44]。
3.1.2 LOX活性
如1.2节中所阐述,LOX主要对酶促氧化中不饱和脂肪酸的氧化有促进作用,LOX活性能够影响脂质氧化的程度。Jin Guofeng等研究了培根在腌制和熟化过程中LOX活性与脂质氧化的关系,发现随着腌制时间的延长,LOX活性显著升高,TBARS含量显著上升;在成熟过程中,LOX活性和TBARS含量均显著下降[24]。由此可见,脂质氧化与LOX活性呈正相关,LOX活性越强,脂质氧化程度越大。
3.1.3 金属元素
金属元素在自动氧化生成氢过氧化物的过程中起到催化剂的作用[45]。金属元素如铁元素,广泛存在于动物体内,在肉制品加工过程不易被消除,是造成肉制品脂质氧化的一个关键因素[46]。在过渡态金属离子中,Fe2+对脂质氧化的诱导效果最强,而Cu+、Cu2+、Cd2+可以抑制脂质氧化[47]。Shahidi等的研究表明,肌红蛋白变性释放的铁元素能够明显促进脂质氧化,而添加金属离子螯合剂后促氧化作用则显著下降[48]。Carlsen等综述了亚铁血红素与脂质氧化的关系,认为非血红素铁对脂质氧化有显著的促进作用,Fe2+比Fe3+对脂质氧化的促进效果更明显[11]。Zhou Feibai等以猪背最长肌匀浆液为研究对象,探究Fe2+对肌肉氧气消耗率和脂质氧化的影响,发现添加FeSO4·7H2O的处理组比对照组氧气消耗率更大,脂质氧化程度更高[32]。
3.1.4 蛋白氧化及肌红蛋白氧化
在肉制品加工过程中,脂质与蛋白质作用形成蛋白质-脂质复合物,使二者的氧化进程相互促进[49]。Viljanen研究了不同食物模型中蛋白质氧化与脂质氧化的关系,发现TBARS含量与蛋白质羰基含量显著相关,这说明脂质氧化与蛋白质氧化能够相互促进[50]。Utrera等以牛肉饼为研究对象,探究蛋白氧化与脂质氧化的关系,结果表明,蛋白质氧化与脂质氧化之间存在相互作用,一种物质的氧化将会促进另一种物质的氧化[51]。除此之外,肌红蛋白的氧化也会促进脂质氧化的发生,研究表明,在氧合肌红蛋白氧化成高铁肌红蛋白的过程中会产生O2-·等一些活性物质,这些活性物质能够进一步促进氧合肌红蛋白的氧化,并且通过促进不饱和脂肪酸的氧化加速脂质氧化[52]。O2-·迅速分解为H2O2,它与高铁肌红蛋白反应形成一种高铁肌红蛋白复合物,这种复合物能够促进脂质氧化[48]。肌红蛋白浓度越高,脂质氧化程度越高[53],二者能够相互促进,使肉色稳定性降低。
3.1.5 pH值
pH值也是影响脂质氧化的一个重要因素。动物屠宰以后,pH值下降产生的H+能够促进肌红蛋白的氧化以及脂质氧化的发生[9]。郭建凤等研究了不同贮藏条件下猪肌肉pH值和TBARS含量的关系,结果表明,在-20 ℃的贮藏条件下,pH值与TBARS含量呈显著负相关,这说明较高的pH值条件下脂质氧化程度较低[54]。同样,Zhang Yimin等研究了气调包装对不同极限pH值牛肉肉色的影响,发现在高氧气调包装中,中间pH值(5.8~6.2)组和高pH值(6.2以上)组牛排的脂质氧化程度显著低于低pH值(5.4~5.8)组[55],这与文献[54]的研究结果一致,这说明较高的pH值条件下脂质氧化程度较低,能维持较高的肉色稳定性。
3.2 外部因素
3.2.1 贮藏温度
温度是影响脂质氧化的重要外部因素之一。温度对脂质氧化的影响主要体现在两个方面。一方面是低温贮藏能够有效抑制脂质氧化[57]。张海容以牦牛脂肪为研究对象,探究温度对牦牛脂质氧化的影响,发现(4±1)℃贮藏条件下脂质氧化程度显著低于(9±1)℃和(15±1)℃的脂质氧化程度[56]。Wang Zhaoming等探究发现,冰温贮藏(-3.5±0.5)℃下的兔肉的脂质氧化、蛋白氧化程度和高铁肌红蛋白的积累速率显著低于传统冷藏条件(4±1)℃[17]。所以对生鲜肉来说,冰温贮藏是抑制脂质氧化的一个重要因素。
温度影响脂质氧化的另一种情况是在肉制品加热过程中,肌红蛋白结构遭到破坏,大量的铁离子释放出来,这个过程促进了脂质的氧化。由于传统的加热方式会使肉制品发生剧烈的氧化,近年来,真空低温烹调、微波加热以及欧姆加热的出现,也为肉制品加热提供了新思路。Rasinska等研究了不同加热方式对兔肉脂质氧化的影响,发现与传统的烤制相比,真空低温烹调能够显著降低兔肉熟制过程中的过氧化值和TBARS含量[57]。Alfaia等研究了不同烹饪方式对牛肉脂肪酸和营养品质的影响,结果表明,水煮牛肉比微波加热的牛肉有更高的MDA含量,微波加热更有利于减少肉制品在加热过程中的脂质氧化程度[58]。Tian Xiaojing等发现欧姆加热熟制(中心温度72 ℃)的牛肉中心红度显著高于传统水浴熟制的牛肉,这说明在欧姆加热条件下,肌红蛋白氧化程度降低[59]。此外,戴妍发现,欧姆加热的猪肉块中TBARS含量显著低于传统水浴加热,同时肉块贮藏期间的红度也均显著高于水浴加热组,这说明欧姆加热可以降低熟制过程中的脂质氧化程度[60]。
3.2.2 包装方式
不同的包装方式能起到促进或抑制脂质氧化的作用。研究表明,在包装体系中,当氧气体积分数超过21%时,容易发生脂质氧化[61]。Park等通过研究不同包装对猪肉脂质氧化的影响,发现真空包装有利于抑制脂质氧化,有氧包装则能够促进脂质氧化[62]。Mancini等研究了琥珀酸盐对绞碎牛肉脂质氧化的影响,发现与真空包装相比,高氧气调包装中的牛肉饼脂质氧化程度更高,而添加琥珀酸盐能够降低TBARS含量[63]。这说明真空包装能够有效抑制脂质氧化,若采用高氧气调包装,可以适当添加抗氧化剂抑制脂质氧化。
3.2.3 盐类的添加
除以上因素外,盐类也是影响脂质氧化的一个重要因素。在肉制品加工过程中,食盐是一种重要的调味品,能够对肉制品的风味产生很大的影响。有研究认为,在肉制品加工过程中加入食盐能促进脂质氧化[9]。但Kong Fanbin等研究表明,添加质量分数1.5% NaCl对于三文鱼脂质氧化并没有显著影响,这可能是由于食盐在一定的浓度范围内能够促进脂质氧化,添加过多能够抑制脂质氧化[64]。
除食盐外,钙盐和亚硝酸盐等其他盐类也会影响脂质的氧化。在肉制品加工过程中,钙盐的添加会促进脂质氧化的发生[65]。Harris等研究了饲料中添加VE对宰后注射氯化钙的牛排肉色的影响,与未注射氯化钙组和添加VE组相比,贮藏7 d后,注射氯化钙的牛排TBARS含量显著升高,脂质氧化程度更高,并且在放置7 d后,牛排的a*值显著下降[66]。亚硝酸盐往往作为发色剂在腌肉制品的加工过程中添加使用。研究表明,亚硝酸钠的添加能够减少MDA的形成,显著抑制干腌肉制品中的脂质氧化程度[67]。Šojić等研究了添加香菜精油对不同含量亚硝酸盐的熟猪肉肠品质和安全性的影响,也得到了相同的结果,在对照处理(不添加香菜精油)组中,不添加亚硝酸盐的熟猪肉肠在贮藏60 d后,MDA含量达到0.5 mg/kg,当亚硝酸盐添加量分别为50、100 mg/kg时,MDA含量则分别降为0.37、0.2 mg/kg[68]。
4 脂质氧化的控制措施
脂质氧化一方面会加速肉与肉制品失色褐变,降低消费者的购买欲,造成经济损失;另一方面,它还会产生许多不良风味,对肉与肉制品的品质产生不良影响;此外,脂质氧化还会促进蛋白的氧化,造成肉品品质劣变。因此,肉品行业亟需采取有效的措施来抑制脂质氧化,维持产品贮藏期间内的肉色和品质。目前,在肉制品工业中,通常采用低温保藏、气调包装或真空包装、低温贮藏结合不同的包装方式和添加抗氧化剂等措施来抑制肉中的脂质氧化。其中应用较多的是在低温贮藏条件下改善包装方式或在肉制品中添加抗氧化剂等。
4.1 改善包装方式
目前市场上生鲜肉传统的包装方式有托盘包装、真空包装、气调包装等。真空包装中氧气含量极低,在贮藏期间肌红蛋白氧化和脂质氧化程度较低,有利于肉色稳定性的保持。研究表明,高氧气调包装只能在短期保持良好的肉色,随着贮藏时间的延长,氧气含量升高会加速肌红蛋白氧化,使肉色稳定性降低[69]。同时,高氧气调包装能够促进脂质氧化,脂质氧化产生的醛类物质能够对肉色稳定性造成不良影响。Yuan Lu等研究了高氧气调包装和真空包装对冷鲜猪肉脂质氧化的影响,结果表明真空包装中的脂质氧化程度显著低于高氧气调包装[70]。
除此之外,食品工业领域研究人员多年来致力于新包装系统的开发,这种新技术主要是在包装材料中加入活性物质从而达到抑制脂质氧化的目的。近年来,随着天然抗氧化剂的研究与开发,以及消费者对天然成分的青睐,许多学者尝试在包装材料中加入天然抗氧化物质开发活性包装。为了提高牛肉的抗氧化性,Letricia等研发了一种在低密度聚乙烯材料中添加质量分数为3%、10%、20%的迷迭香萃取物的新型活性包装,这种活性包装显著抑制了牛肉在贮藏期间的TBARS含量,起到良好的脂质氧化抑制效果[71]。Nisa等以马铃薯淀粉为基质生物聚合膜,添加5%的绿茶制成活性包装膜包裹新鲜牛肉,与对照组相比,在-4 ℃贮藏10 d显著降低了高铁肌红蛋白含量和TBARS含量[72]。这说明添加天然抗氧化成分的活性包装能在一定程度上起到良好的抗氧化和护色效果。
因此,无氧包装能够有效抑制脂质氧化,保持肉色稳定性,而高氧气调包装可以在较短的贮藏时间内保持良好的肉色。对于脂肪含量较高的高档牛肉,更适宜采用真空包装,防止由于脂质氧化造成的肉品失色褐变。新型活性包装的研发也为控制肉制品脂质氧化、保持肉色稳定性提供了新的思路,与传统的气调包装相比,新型活性包装能够抑制脂质氧化、延长货架期,但是新型活性包装的研究尚不完善,有可能给消费者带来潜在的健康危害。
4.2 添加抗氧化剂
近年来,天然抗氧化剂在肉制品中的使用越来越广泛。其使用方法不尽相同,可直接在肉制品加工过程中添加,也有许多研究表明,在饲料中添加天然抗氧化剂,不但可以有效防止屠宰后肌肉的氧化过程,而且能够提高肉类的食用品质[73]。Zhao Junxing等在绵羊饲料中添加酿酒葡萄皮渣来研究其对绵羊肉抗氧化活性的影响,酿酒葡萄皮渣含有丰富的多酚类物质,具有很强的抗氧化能力。研究发现,日粮中添加5%、10%的酿酒葡萄皮渣可以显著降低绵羊肉中MDA的含量,对脂质氧化起到良好的抑制作用[74]。
许多学者在肉制品中直接添加天然抗氧化剂,发现多种天然产物具有较好的抗氧化效果。在新鲜的绞碎牛肉中添加茼蒿、南瓜、韭菜、刺五加、大豆、蜂斗菜等能够显著降低绞碎牛肉的TBARS含量[75-76]。此外,在牛肉饼中添加不同浓度的橄榄油副产物,发现4%的添加量能够抑制脂质氧化[77]。Zhou Feibai等研究了所添加绿茶、迷迭香、马蹄提取物在总酚含量为25~250 mg/kg时对猪肉的抗氧化保护作用,结果表明,这3 种物质都能抑制脂质氧化,其中迷迭香的抗氧化效果最佳,但迷迭香对肌红蛋白氧化的抑制作用不明显,可能是由于迷迭香中含有较多的疏水性多酚,由于硫醇-醌的相互作用而导致硫醇基的损失,从而使肌红蛋白氧化;而绿茶和马蹄提取物对脂质氧化和肌红蛋白氧化均表现出较好的抑制作用,提高了肌红蛋白的稳定性[78]。
近年来,从水果中提取的天然抗氧化剂在肉制品中的应用得到了广泛的研究。荔枝果皮中含有大量多酚、黄酮以及花青素,具有良好的抗氧化活性。Das等研究了荔枝果皮提取物对熟制羊肉块脂质氧化的影响,结果表明,与对照组相比,添加1.0 g/100 g和1.5 g/100 g的荔枝果皮提取物抑制了整个贮藏期间的脂质氧化[79]。Qi Suijian等在肉酱中添加体积分数0.1%、0.5%、1.0%的荔枝籽提取液也能够显著抑制肉酱的脂质氧化[80]。
4.3 新型低温贮藏技术
温度是影响脂质氧化的重要因素,在肉制品贮藏过程中,低温可以有效抑制脂质氧化的发生。冷冻是目前市场上常见的鲜肉贮藏和运输方式。Xia Xiufang等认为传统的冷冻再解冻过程会加速肉制品脂质氧化的发生[81]。Benjakul等认为冷冻过程形成的冰晶会对细胞膜造成损伤,释放出的非血红素铁能够促进脂质氧化的发生[82]。为改进传统冷冻的缺陷,近年来,研究人员开发了一些新型低温保藏技术,如冰温保鲜技术、微冻技术、高压冷冻技术、电场辅助冻结技术及微波辅助冷冻技术等。与传统的冷冻方式相比,新兴冷冻技术可以改善冷冻生鲜肉品的某些品质或提高冷冻效率,其中磁场辅助冷冻、电场辅助冷冻、射频辅助冷冻均可以改善冷冻过程对肉色的不利影响[83]。随着冰温保鲜和微冻技术研究的逐渐深入,人们发现这两种低温贮藏方式对脂质氧化有较好的抑制效果[84]。研究表明,冰温条件下肌红蛋白氧化速率显著降低,是因为冰温能够减少血红素和原卟啉这两种脂质氧化催化剂的释放,从而对脂质氧化产生良好的抑制效果[85]。程玉平等以猪背最长肌为研究对象,探究新型微冻液冷冻法和传统冷冻法对猪肉饼品质的影响,发现新型微冻液冷冻的猪肉饼TBARS含量显著低于传统冷冻法[86]。由此可见,与传统的冷冻技术相比,微冻技术和冰温保鲜技术能够显著抑制脂质氧化,使肉类保持良好的食用品质。随着肉制品行业的不断发展和市场需求的扩大,这两项低温保藏技术也将得到快速的发展。
5 结 语
肉色是评价肉类品质的一个重要指标,如何维持良好肉色始终是行业关注的焦点。而脂质氧化是影响肉色的一个重要因素,虽然脂质氧化初级代谢产物对肉色稳定性的影响基本明确,但除了HNE和MDA这两种脂质氧化次级代谢产物外,其他大量的脂质氧化次级代谢产物对肉色稳定性的影响尚不完全明确。除脂质氧化与肌红蛋白状态和线粒体功能的关系外,肌肉系统中脂质氧化耗氧程度是否会对肌红蛋白稳定性造成更大的影响,仍然是一个值得探究的问题。
脂质氧化不但会造成生肉褐变失色,在肉制品熟制过程中也会发生PMB现象。该现象容易使消费者造成错误的判断,导致食用热杀菌不彻底的产品。这些问题的存在促使适宜的脂质氧化抑制措施的开发,以维持较好的肉色和肉色稳定性。近年来,蛋白质组学和脂质组学的发展为利用分子技术分析脂质氧化使肉色劣变的机制提供了可能,也为进一步开发有效的抗氧化措施提供理论基础和科学依据。
