东北地区阔叶红松林典型群系植物多样性研究
2020-11-25朱玉鹏
朱玉鹏
(东北林业大学,黑龙江 哈尔滨 150040)
红松(Pinuskoraiensis) 要求温和凉爽的气候,在北半球中高纬度地区有较大面积的分布[1]。世界红松森林面积约50万km2,中国约占其中的60%[2-3]。红松通常与多种阔叶树构成混交林,称为阔叶红松林,分布于温带针阔混交林区域,植物种类丰富,且特征种与孑遗种较多,具有特殊的生物多样性保护价值[4-6]。生物多样性是指物种水平的生物多样性,研究范围涉及多样性的测度、关键种及其保护、干扰下的生态系统多样性以及动态监测等[7-9]。而植物群落物种多样性是生物多样性的重要组成部分,是森林群落健康的基础内容之一[10-14]。森林群落的植物物种多样性与土壤、地形、气候以及群落生产力等因子相关,也在一定程度上反映了森林植被的演化历史[15-17]。近几十年,东北地区的原始红松林遭到了严重破坏,植物多样性丧失[18-20]。在此背景下,阔叶红松林植物多样性的保护和恢复被广泛关注[21-23]。本研究以东北地区天然阔叶红松林为调查对象,利用双向指示种分析方法(TWINSPAN)并根据经验及相关记录进行校正,将调查区域分布的阔叶红松林划分为红松-红皮云杉林、红松-杉松林、红松-簇毛枫林等3种主要群系类型,通过对比3种群系类型内乔灌草各层的物种丰富度、Shannon-Wiener多样性指数、Simpson多样性指数和pielou均匀度指数,并分别计算各林型各层片间的物种相似度,探讨了东北地区不同的阔叶红松林群系类型的差异性和相似性,为阔叶红松林生物多样性的保护和恢复措施提供科学依据。
1 研究区域与研究方法
1.1 研究区域概况
丰林国家级自然保护区位于黑龙江省伊春市境内,地理坐标为E128°58′—129°15′、N 48°02′—48°12′,海拔为280—683m。年平均气温为0.2℃,年降水量为680—750mm,年蒸发量930mm,无霜期100—110d。
黑龙江凉水国家级自然保护区位于黑龙江省伊春市境内,地理坐标为E128°47′—128°57′,N47°06′—47°16′,海拔为280—707m。年平均气温为-0.3℃,年降水量为676mm,年蒸发量805mm,无霜期100—120d。
长白山自然保护区位于吉林省的东南部,地理坐标为E127°42′—128°16′,N41°41′—42°25′,海拔为691—2 691m。年均气温在3—7℃,年降水量为700—1 400mm,无霜期100d左右。
吉林省蛟河林业实验区管理局位于吉林省蛟河市境内,地理坐标为E127°35′—129°15′,N43°51′—44°05′,海拔为330—1 176m。年平均气温1.7℃,年降水量为800mm左右,无霜期120—150 d。
吉林露水河国家森林公园位于吉林省抚松县境内,E127°01′—128°06′,N41°42′—42°49′。年平均日照2 352.5h,日照百分率为53%,年降水量为800mm左右,无霜期变化较大。
白石砬子自然保护区位于宽甸满族自治县北部,地理坐标为E124°44′—124°57′,N40°50′—40°57′,海拔为404—1 271m。年平均气温6.4℃,年平均降水量1 158mm,年蒸发量885mm,无霜期132d左右。
老秃顶自然保护区位于辽宁省东部桓仁、新宾两县交界,地理坐标为E124°41′—125°05′,N41°11′—41°21′,海拔为465—1 358m。年平均气温6.2℃,无霜期139d左右。
1.2 研究方法
1.2.1 植被调查方法
(1)样地设置及群落调查。
在研究区域内,选择植被保存较完整的红松阔叶混交林群落进行研究。设20m×20m的乔木样地,在乔木样地的四角,设4个5m×5m的灌木样地和4个1m×1m的草本样地。
对乔木层进行每木检尺,记录胸径、树高、枝下高、冠幅。灌木层植株记录高度、株数、地径等。草本层记录株数、盖度、平均高。调查中将高度高于3m的大型灌木、低于3m的乔木幼树、低于0.5m的乔木和灌木的幼树或幼苗分别计入乔木层、灌木层和草本层。本研究共调查样方79块,其中红松-红皮云杉林26块;红松-簇毛枫林29块;红松-杉松林24块。
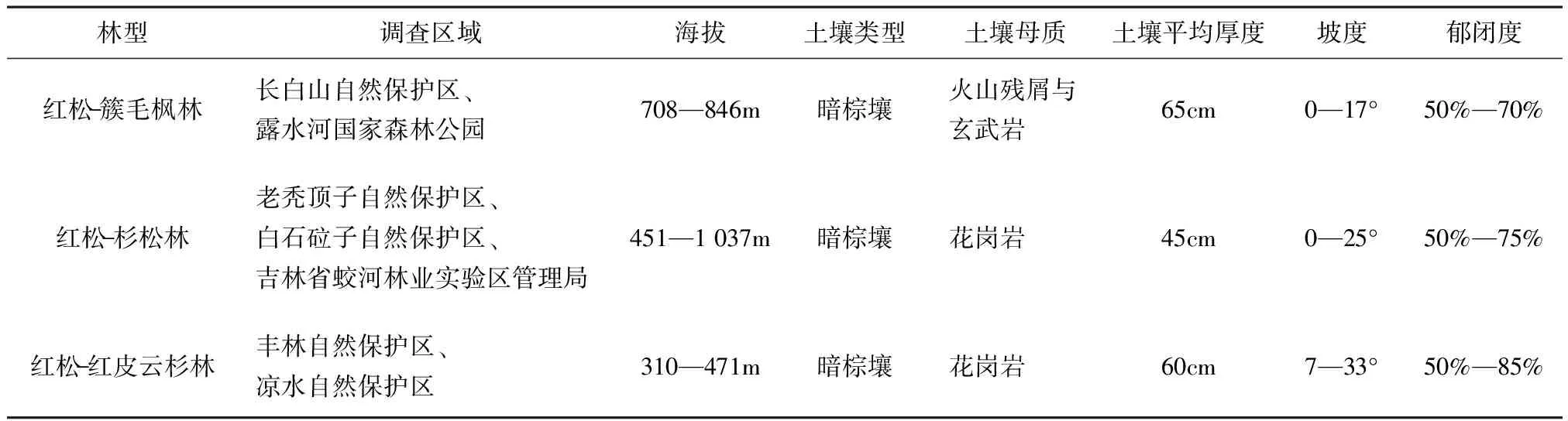
表1 样地基本情况
(2)环境因子调查。
记录样地内及周围的环境因子,包括海拔高度、坡度、坡向、坡位、人为干扰程度等情况。
(3)调查时间。
丰林自然保护区、凉水自然保护区、长白山自然保护区、露水河国家公园及老秃顶子自然保护区的调查时间为2018年的7月至9月,吉林省蛟河林业实验区管理局及白石砬子自然保护区的调查时间为2019年9月。
1.2.2 植被的分类及命名
特定物种组合或物种在一些群落中出现的概率较高,而在另一些群落中出现的概率较低,这种具有生境偏好的物种组合或物种,被称为特征种。在优势种有时不明显的混交林中,难以根据优势种划分出群落类型,在这种情况下,将特征种作为植物群落的划分依据具有很大的普适性。特征种同时是群落的优势种和常见种时,对群落的分类有重要意义,本文将这类物种的名称用于群落的命名。并利用软件JUICE7.0.84对样方采用双向指示种分析方法(TWINSPAN)根据乔木层物种组成对阔叶红松林进行数量分类并根据经验及相关记录校正,得到各群系类型的优势种、常见种和特征种,最终根据乔木层物种组合的不同将调查样地划分并命名为为红松-红皮云杉林、红松-杉松林、红松-簇毛枫林3种群系类型。
1.2.3 植被群落物种多样性计算
植被群落物种多样性采用以下四种多样性指数进行计算。公式分别为:
①丰富度指数:S=N
②Shannon-Wiener多样性指数(H′):
④pielou均匀度指数:E=H′/lnS
式中:S为样地中的物种总数,N为个物种全部植株个体总数,Ni为第i物种的植株个体数。a是群落1和群落2都有的物种数量,b是群落2有但群落1无的物种数量,c是群落1中有但群落2中无的物种数量。
2 结果与分析
2.1 物种组成
2.1.1 红松-杉松林的物种组成
林内共有维管束植物90种,隶属于43科67属。其中,蕨类植物5科8属11种,裸子植物1科2属3种,双子叶植物33科50属66种,单子叶植物4科7属10种(见表2)。仅含有1—2个种的科有30个,占总数的69.77%。仅含1个种的属有50个,占总数的74.63%。说明该区群落的科属组成结构相对分散。各层优势种均不显著,常见种有杉松(Abiesholophylla)、色木枫(Acerpictum)、千金榆(Carpinuscordata)、胡桃楸(Juglansmandshurica)、红松(Pinuskoraiensis)、紫椴(Tiliaamurensis)、裂叶榆(Ulmuslaciniata)、刺五加(Eleutherococcussenticosus)、金花忍冬(Lonicerachrysantha)、东北山梅花(Philadelphusschrenkii)、粗茎鳞毛蕨(Dryopteriscrassirhizoma)、荨麻叶龙头草(Meehaniaurticifolia)等,特征种有胡桃楸、杉松、千金榆、天女花(Oyamasieboldii)、戟叶耳蕨(Polystichumtripteron)、荨麻叶龙头草等。
2.1.2 红松-簇毛枫林的物种组成
林内共有维管束植物109种,隶属47科81属。其中,蕨类植物5科6属7种,裸子植物1科2属3种,双子叶植物36科65属89种,单子叶植物5科8属10种(表3)。仅含有1—2个种的科有34个,占总数的72.34%;仅含1个种的属有50个,占总数的80.25%。说明该区群落的科属组成结构相对分散。各层优势种均不显著,常见种有色木枫、簇毛枫(Acerbarbinerve)、青楷枫(Acertegmentosum)、紫花枫(Acerpseudosieboldianum)、红松、蒙栎(Quercusmongolica)、紫椴、东北山梅花、四花薹草(Carexquadriflora)、荨麻叶龙头草等;特征种有簇毛枫、紫花枫、蒙栎、早花忍冬(Lonicerapraeflorens)、木贼(Equisetumhyemale)、东北百合(Liliumdistichum)等。
2.1.3 红松-红皮云杉林植物的物种组成
林内共有维管束植物116种,隶属50科87属。其中,蕨类植物5科8属10种,裸子植物1科3属5种,双子叶植物41科70属94种,单子叶植物3科6属7种(表4)。仅含有1—2个种的科有34个,占总数的74%。仅含1个种的属有50个,占总数的77.01%。说明该区群落的科属组成结构相对分散。各层优势种均不显著,常见种有色木枫、花楷枫(Acerukurunduense)、红松、红皮云杉(Piceakoraiensis)、紫椴、春榆(Ulmusdavidianavar.japonica)、毛榛(Corylusmandshurica)、刺五加、金花忍冬、东北羊角芹(Aegopodiumalpestre)、东北蹄盖蕨(Athyriumbrevifrons)、毛缘薹草(Carexpilosa)、蚊子草(Filipendulapalmata)等;特征种有红皮云杉、臭冷杉(Abiesnephrolepis)、花楷枫、珍珠梅、金花忍冬、蚊子草、东北蹄盖蕨等。
2.2 种群系类型乔木层、灌木层和草本层物种多样性特征比较分析
图1分别对3种森林类型间乔木层、灌木层、草本层的物种多样性指数进行比较。
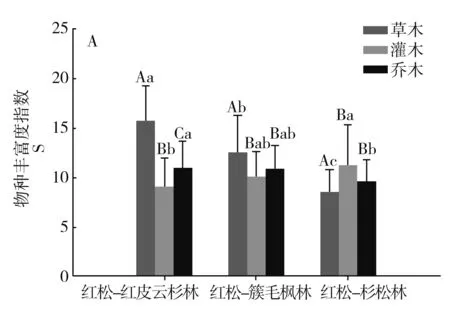
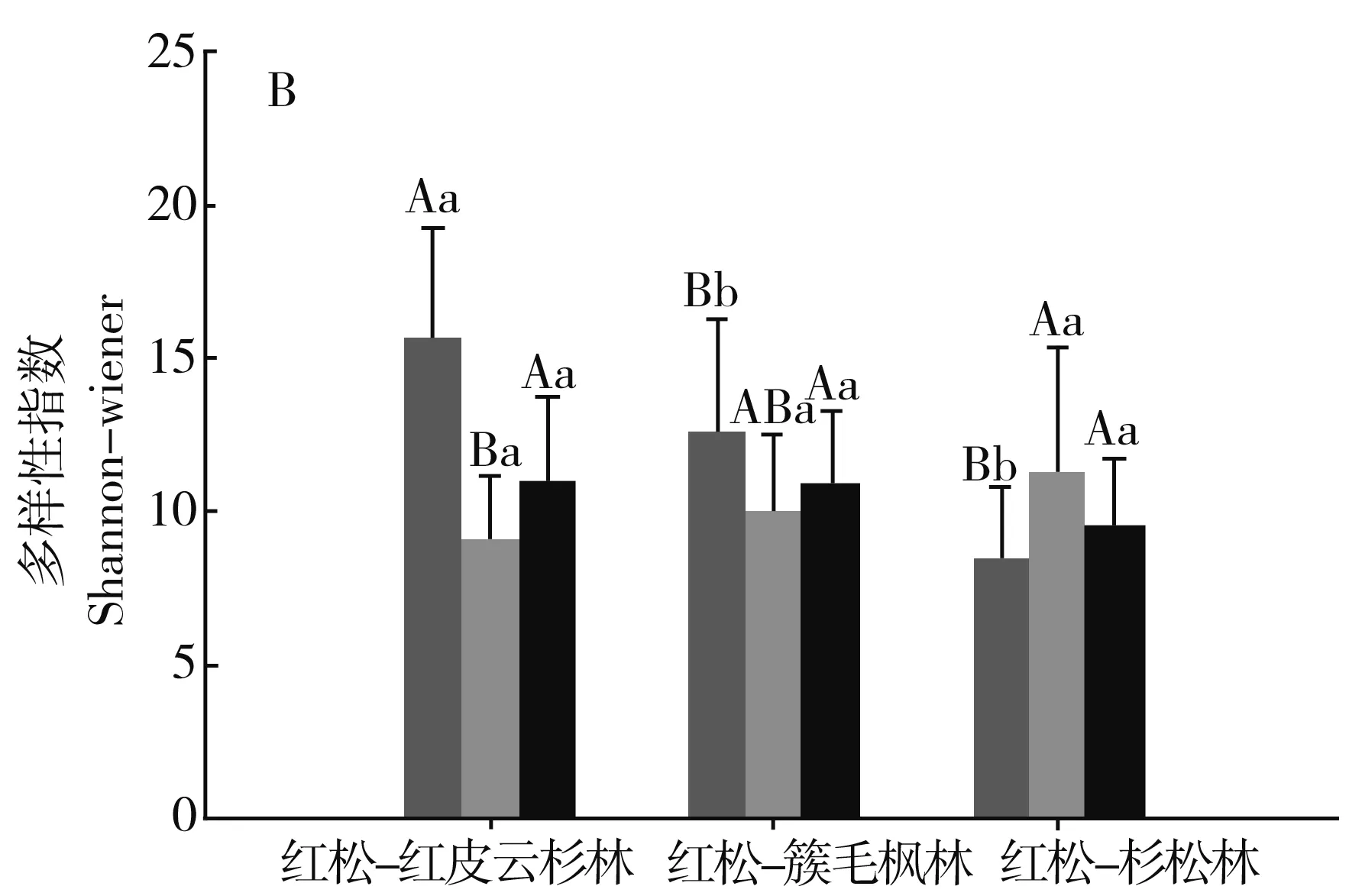
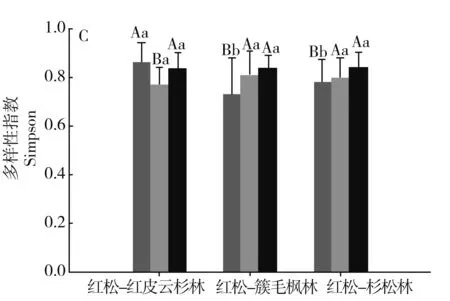
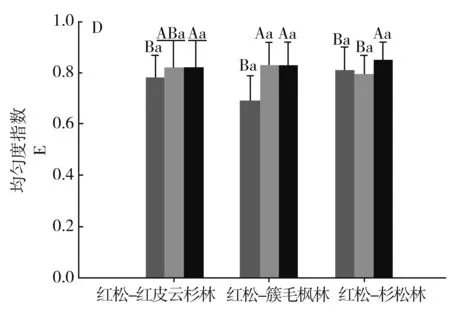
注:不同小写字母表示不同林型间差异显著,不同大写字母表示不同组分间差异显著。P<0.05。
3种群系类型乔木层的丰富度指数S表现为红松-红皮云杉林>红松-簇毛枫林>红松-杉松林,且红松-红皮云杉林与红松-杉松林的差异性达到显著水平;Shannon-Wiener(H′)、Simpson(λ)和均匀度指数(E)相近,且相互之间差异性均不显著。
3种群系类型灌木层的丰富度指数S表现为红松-红皮云杉林<红松-簇毛枫林<红松-杉松林,且红松-红皮云杉林与红松-杉松林的差异性达到显著水平,且红松-红皮云杉林与红松-杉松林的差异性达到显著水平;Shannon-Wiener(H′)、Simpson(λ)和均匀度指数(E)相近,且相互之间差异性均不显著。
3种群系类型草本层的丰富度指数S、Shannon-Wiener(H′)、Simpson(λ)表现为红松-红皮云杉林最高,且与红松-簇毛枫林、红松-杉松林的差异性达到显著水平,而红松-簇毛枫林与红松-杉松林相近,差异性不显著;均匀度指数(E)表现为红松-簇毛枫林最低,且与红松-红皮云杉林、红松-杉松林的差异性达到显著水平,而红松-红皮云杉林与红松-杉松林相近,差异性不显著。
相同群系类型不同的片层间,红松-红皮云杉林三个层片的丰富度指数S、Simpson(λ)、Shannon-Wiener(H′)均表现为草本层>乔木层>灌木层,且灌木层与草本层、乔木层的差异性显著。红松-簇毛枫林三个片层的丰富度指数S、Shannon-Wiener(H′)表现为草本层>乔木层>灌木层,且草本层与灌木层、乔木层的差异性显著,Simpson(λ)指数表现为草本层<灌木层<乔木层,且草本层与灌木层、乔木层的差异性显著,原因为部分样方拥有较为潮湿的林下环境,草本层的华金腰或木贼占有较大的数量优势。红松-红皮云杉林三个片层的丰富度指数S、Simpson(λ)Shannon-Wiener(H′)均表现为草本层<乔木层<灌木层,且草本层与灌木层、乔木层的差异性显著;Simpson(λ)指数表现为草本层<灌木层<乔木层,且草本层与灌木层、乔木层的差异性显著。
2.3 群落相似性
从表2、表3、表4可以看出,3种群系类型之间的乔木层、灌木层和草本层的群落相似系数均低于0.50。红松-红皮云杉林与红松-簇毛枫林、红松-杉松林乔木层的群落相似系数为0.40、0.47,红松-簇毛枫林与红松-杉松乔木层的群落相似系数为0.44,数值在0.25—0.50之间,为中等不相似。红松-红皮云杉林与红松-簇毛枫林、红松-杉松林灌木层的群落相似系数为0.43、0.38,红松-簇毛枫林与红松-杉松灌木层的群落相似系数为0.43,为中等不相似。红松-簇毛枫林与红松-杉松林、红松-红皮云杉林草本层的群落相似系数为0.34、0.28,为中等不相似。红松-簇毛枫林与红松-杉松草本层的群落相似系数为0.21,数值小于0.25,为极不相似。

表2 3种阔叶红松林群系乔木层植物群落相似性
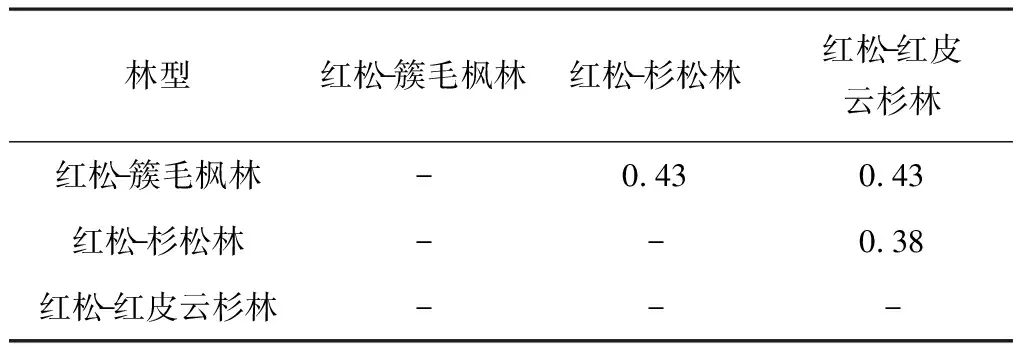
表3 3种阔叶红松林群系灌木层植物群落相似性

表4 3种阔叶红松林群系草本层植物群落相似性
3 讨论
本研究中,红松阔叶混交林的3个群系类型都具有较高的物种丰富度及多样性。王世雷等[24]的研究发现,针阔混交林同时具备了阔叶林和针叶林的林分特征,导致生境异质性的增加,使其具有了更高的物种丰富度与多样性,这与本研究调查结果相同。3种阔叶红松林林型中,科属组成结构都相对分散,这与于洋等[25]对小兴安岭原始红松林的研究结果相同。
3种群系类型中,除红松-红皮云杉林乔木层物种丰富度最高且显著高于红松-杉松林外,乔木层的Shannon-Wiener(H′)、Simpson(λ)均相近;而灌木层与草本层物种丰富度与多样性表现最好的林型分别为红松-杉松林和红松-红皮云杉林,红松-杉松林草本层物种丰富度与多样性表现最差;红松-簇毛枫林草本层的均匀度,显著低于另两种群系类型。3种群系类型之间,乔木层、灌木层和草本层的群落相似系数,均呈现为中等不相似或极不相似。
物种多样性依赖于空间异质性、演替阶段及演替过程对生境的改造作用,使生态系统承载力发生改变,是物种丰富度与多样性产生差异的原因[26-27]。前人基于大尺度的研究表明,气候因子对木本植物物种丰富度的影响显著。余敏[28]对山西灵空山林下草本群落的研究表明,冠层物种组成和冠层结构的不同,是林下草本群落生物多样性产生差异的重要原因。陆龙龙[29]对长白山地区阔叶红松林的研究显示,不同的演替阶段,植被各层的物种丰富度存在差异,乔木层的丰富度在演替初期最小,在中期及后期升高;灌木层的丰富度表现为先增大后减小,演替中期和演替中前期有较高的物种数;草本则表现为先减小再增大。除地理及环境因子的影响外,人为干扰也是影响森林群落结构和物种丰富度的重要影响因子[30]。马菁[30]对小兴安岭红松阔叶混交林的研究显示,阔叶红松林中物种丰富度、多样性指数各层均为草木层>灌木层>乔木层,物种多样性呈现出随植被所处林层高度的提升而递减的趋势。这与本次研究结果并不相符,原因可能为统计方法不同以及调查区域环境差异所致。马菁的研究中各片层物种主要根据生活型进行划分,而调查范围仅在小兴安岭地区;而本研究中,各片层物种划分除根据生活型之外也将高度作为划分标准,调查范围既包括小兴安岭也包括长白山及其数个支脉。郝占庆等[33]人的研究显示较低的乔木郁闭度在一定程度上有利于灌木层和草本层物种丰富度和多样性的增长。在本研究中,郁闭度最低的红松-簇毛枫林仅草本层的物种丰富度显著高于红松-杉松林,但显著低于红松-红皮云杉林,多样性指数并未明显高于另外两个林型,原因可能是土壤因子、气候因子、地形和冠层物种组成的影响所致。红松-杉松林分布区域原始阔叶红松林已经毁坏殆尽,红松-杉松林调查样地几乎全部为择伐之后经次生演替形成的次生林,其所处的演替阶段及演替过程可能是灌木层生物多样性表现较好而草本层表现差的重要因素。同时,林内灌木层较大的物种多样性,意味着林下的草本植被获得的光照减少,草本的生长受到抑制,使其生物多样性有所降低。
综上所述,东北地区不同群系类型的红松阔叶混交林之间,植物多样性之间存在的差异应为多种因素的共同作用下形成的。
对红松阔叶混交林植物多样性的正确认识,是保护该植被的重要前提。本次研究显示,红松-红皮云杉林、红松-簇毛枫林和红松-杉松林的物种丰富度、多样性指数和物种组成均存在着明显差异。由此可见,乔木层物种组成的不同也意味演替过程、立地条件等影响因子的不同,进而造成了灌木层和草本层物种丰富度、多样性指数和物种组成,因此对于不同的群系类型需要根据其特点进行更为科学有效的管理。
