亚热带常绿林不同冠层小枝叶面积-叶生物量关系研究
2020-11-24王满堂陈晓萍钟全林程栋梁
杨 力,王满堂,陈晓萍,孙 俊,钟全林,程栋梁,*
1 福建师范大学福建省植物生理生态重点实验室,福州 350007 2 福建师范大学地理研究所,福州 350007 3 枣庄学院城市与建筑工程学院,枣庄 277160
当年生小枝作为植物生长分支系统最活跃的部分,是木本植物重要的功能组成单元。研究当年生小枝叶片的性状特征,有助于深入了解当年生小枝内部的生物量分配[1]和植物的环境适应机制[2- 4]。
叶片是植物进行光合作用的主要器官,叶面积与叶生物量之间的关系不仅反映了植物的光截获能力,也反映了植物的环境适应能力[5- 6],对植物个体生长乃至群落生产力均有重要影响[7]。前期研究表明叶面积(Leaf Area, LA)与叶生物量(Leaf Mass, LM)存在显著的异速生长关系:LA=βLMα,其中β为异速生长常数,α为异速生长指数[8]。Niklas等[9]研究发现,植物的LA与LM之间的异速生长指数<1.0(即α<1.0),表明叶面积随着叶生物量的增加而降低,进而提出了“收益递减”假说(“Diminishing returns” hypothesis)。该假说认为植物叶面积随生物量的增加而降低,导致光合有效面积随叶构造成本的增加而降低。随后一些研究在不同尺度,不同区域对“收益递减”假说进行了探索[10- 11]。例如,Milla和Reich[12]对六个不同数量级上的叶面积与叶生物量的研究结果支持“收益递减”假说。相反Price等[13]的研究发现叶面积与叶生物量之间呈等速生长的关系。Pan等[14]对不同海拔植物群落水平的研究发现LA与LM之间的异速生长指数随着海拔高度的升高逐渐从α<1.0向α>1.0有规律变化。然而,Li等[15]对武夷山不同海拔黄山松小枝叶性状的研究,结果表明LA与LM之间的异速生长指数均<1.0,支持收益递减假说。这些不同的研究结果表明物种组成与环境可能会影响植物LA与LM之间的异速生长关系。
植物不同冠层光照的有效性是影响植物叶片性状的一个重要环境因素[16-17]。例如,Okobo等[18]对印度尼西亚混合林木本植物叶性状的研究,结果发现位于林下的比叶重(Leaf Mass Per Area, LMA)低于上部。Sterck和Bongers[19]的研究表明植物叶性状受到冠层高度光照影响,随着不同冠层高度光照有效性的降低,冠层宽度显著增加,叶片LMA显著降低。这些研究结果均表明冠层高度可能会影响植物LA与LM间的异速生长关系。由于目前大部分当年生小枝的叶面积-叶生物量的研究均是从树冠表面截取的当年生小枝[20- 22]。这些研究结果可能会忽略冠层梯度对LA与LM间异速生长关系的影响。
因此,本研究选取江西阳际峰自然保护区内中亚热带常绿阔叶林为研究区域,该区域保存了完好的武夷山中段西侧典型的中亚热带常绿阔叶林森林生态系统,具有重要的研究价值[23]。研究选取样地内落叶与常绿树种上、下冠层位置的当年生小枝进行采样,分析不同冠层高度和生活型植物叶面积与叶生物量是否存在差异;探讨叶面积与叶生物量的关系对冠层和生活型的响应。研究可为进一步了解冠层梯度和生活型叶面积与叶生物量的收益权衡提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于江西阳际峰国家级自然保护区(117°11′30″—117°28′40″E, 27°51′10″—28°02′20″N),地处北武夷山和南武夷山交界处,是连接武夷山各部的重要生态廊桥。该保护区具有典型的湿润中亚热带季风气候。年平均降水量为1870—2191 mm,其中4—6月份降水量72%—75%为降雨量,年平均气温11.4℃—18.5℃。地带性土壤是铝质湿润淋溶土[24]。
1.2 实验设计与样品采集
于2015年8月,建立一个25 hm2(500 m500 m)的大样地,样地森林群落基本特征见表1。对样地内所有胸径≥1 cm的乔木逐一挂牌,记录树种名、胸径及树高(表2)。在2017年8月,选取样地内的69种优势乔木树种进行采样,每个物种选取3株标准木,分别在每株标准木树冠上冠层(1/2冠层以上)完全暴露于太阳光照的位置及冠层下层(1/2冠层以下)完全阴影处的位置,随机选取中等大小、具有成熟叶片、没有明显叶面积损失的3—5个当年生末端小枝,并立即浸入水中,做好相关标记后带回实验室。

表1 阳际峰森林群落林分基本特征
1.3 指标测定
在样地内每个物种选取3株标准木,在每株标准木1/2冠层高度上下截取至少3个当年生小枝,在每个小枝上获取3—5个没有明显叶面积损坏的成熟叶片做好标记后带回室内,使用扫描仪(EPSONV39)[25]扫描后,运用ImageJ图形处理软件获取叶面积数据。将扫描后的叶片,在75℃下烘干48 h以上,使用电子天平(精度为0.001 g)进行称重。最后将扫描后的叶片,计算比叶重LMA=叶干重/叶面积(mg/mm2)。
1.4 数据分析
首先使用Excel 2016软件进行数据整理;其次使用SPSS 19.0对叶面积和叶干重的均值及标准差进行计算,并使用独立样本T检验对不同生活型植物检验分析处于上冠层和下冠层叶之间叶性状的差异,用字母标记组间显著性差异水平。然后对不同林木类型不同冠层的叶面积和叶生物量在R语言软件平台(R3.4.4)进行分析。使用R中的“smatr”程序包[26]分析叶片性状之间的关系。二者之间的关系采用异速生长方程LA=βLMα,其中β为异速常数,α为异速生长指数,LA和LM分别是叶面积和叶生物量。将该方程左右两边同时取对数转换为:logLA=logβ+αlogLM,当=1时,代表因变量和自变量呈等速生长关系,α>1或α<1时,两者表现为异速关系,使用模型II标准化主轴(standardized major axis estimation, SMA)回归分析方法计算异速生长指数和常数[27]。采用Origin 9.0软件进行绘图。
2 结果与分析
2.1 不同冠层位置常绿和落叶物种间叶面积与生物量特征
叶生物量在不同冠层位置和不同林木类型中均无显著差异(P>0.05,图1)。常绿树种上冠层与下冠层叶生物量平均值(mean±s.e.)分别为(216.60±39.28) mg和(214.25±47.14) mg(表3)。落叶树种上冠层与下冠层叶生物量分别为(214.53±47.03) mg和(182.41±31.06) mg(表3)。整体而言,上冠层和下冠层林木的叶生物量分别为(215.82±30.00) mg和(202.25±31.50) mg(表3)。
叶面积在不同冠层高度无显著差异(P>0.05,图1)。但在不同生活型之间存在显著差异(P<0.05,图1)。常绿树种上冠层与下冠层叶面积的平均值(mean±s.e.)分别为:(2317.85±309.88) mm2、(2334.29±327.74) mm2(表3)。落叶树种上冠层与下冠层叶面积分别为:(3027.96±575.61) mm2、(3149.86±484.77) mm2(表3)。整体而言,上冠层与下冠层叶片的叶面积分别为:(2585.43±290.86) mm2、(2641.27±276.04) mm2(表3)。
常绿树种上冠层的比叶重(LMA) (0.09±0.005) mg/mm2与下冠层(0.09±0.003) mg/mm2(表3)无显著性差异(P>0.05,图1)。落叶树种上冠层(0.07±0.003) mg/mm2(表3)与下冠层比叶重(0.06±0.003) mg/mm2有显著性差异(P<0.05,图1)。但上下冠层中常绿树种比叶重均显著高于落叶树种上(P<0.05,图1)。
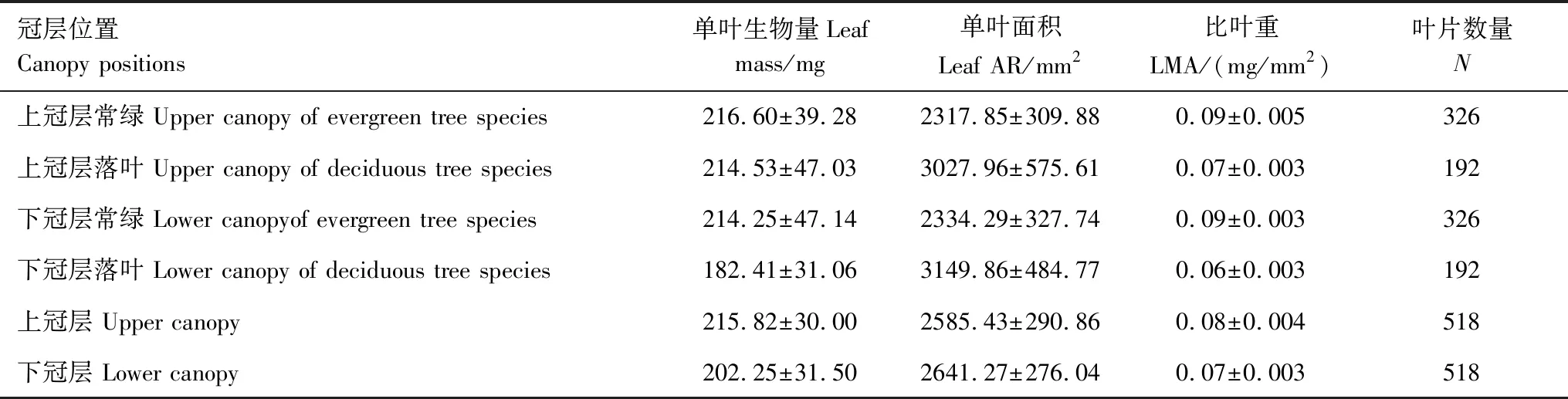
表3 69种亚热带木本植物不同冠层位置和生活型叶性状
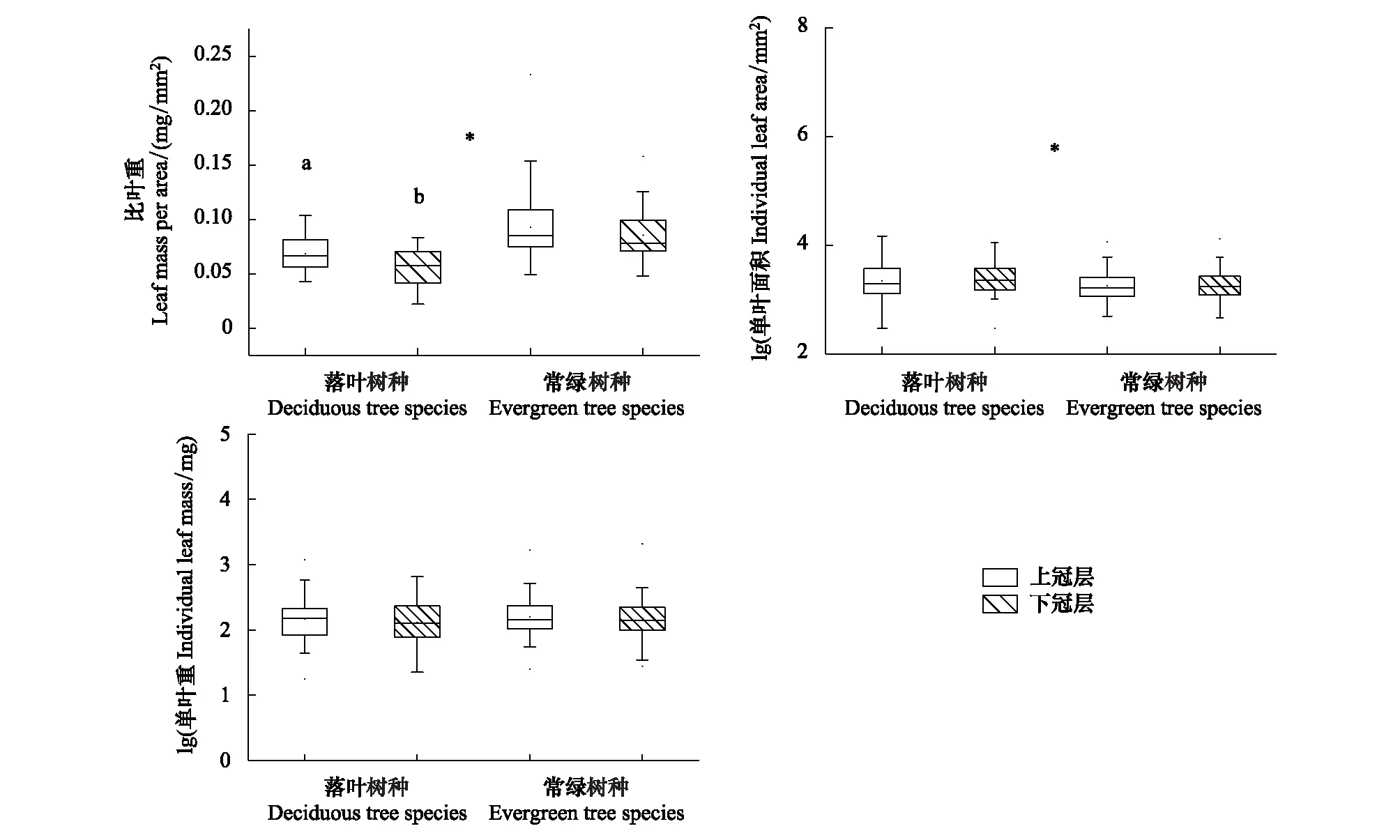
图1 不同冠层位置和生活型叶性状特征Fig.1 Leaf traits in different canopy heights and living form字母a和b表示上冠层与下冠层之间存在显著差异,*表示常绿和落叶植物之间存在显著差异(P<0.05)
2.2 物种水平叶面积-生物量关系
物种水平叶面积与叶生物量间异速生长指数均值为0.90±0.04(n=69)。43种常绿树种中有39种(90.7%)叶面积与叶生物量具有显著的关系(P<0.05,图2),2种(4.7%)林木的异速生长指数显著大于1.0(P<0.05),11种(25.6%)林木异速生长指数显著小于1.0,27种(60.5%)林木异速生长指数与1.0无显著性差异(图2)。26种落叶树种中有17种(65.4%)叶面积与叶生物量具有显著关系(P<0.05,图2)。26种落叶树种叶面积与叶生物量关系中,2种(7.7%)林木的异速生长指数显著大于1.0(P<0.05),6种(23.1%)林木的异速生长指数显著小于1.0(P<0.05),15种(57.7%)林木异速生长指数与1.0无显著性差异(P>0.05,图2)。
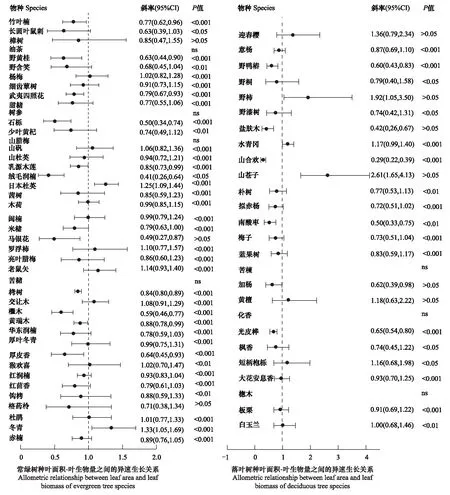
图2 不同生活型物种的叶片面积与叶片生物量异速生长指数Fig.2 Leaf area and leaf biomass allometric growth index of different living species
2.3 不同生活型叶面积与生物量关系
常绿树种与落叶树种不同冠层高度叶面积与生物量拥用共同斜率为0.92(P=0.96,95%CIs:0.86-0.97)(表4,图3),但截距存在显著差异 (P<0.001,95%CIs:1.21—1.46图3)。
单叶面积与单叶生物量之间存在显著的相关性(P<0.05,表4)。常绿树种上冠层与下冠层当年生小枝单叶面积与单叶生物量存在共同斜率为0.92(P=0.60,95%CIs:0.84—1.00),呈等速生长关系(P=0.60)(表4,图3),同时存在共同截距(P=0.30,95%CIs:1.09—1.43)(表4,图3)。落叶树种上冠层与下冠层单叶面积与单叶生物量共同斜率为0.91(P=0.81,95%CIs:0.83—1.01),呈等速生长关系(表4,图3),但不存在共同截距(P=0.01,95%CIs:1.22—1.59图3)。
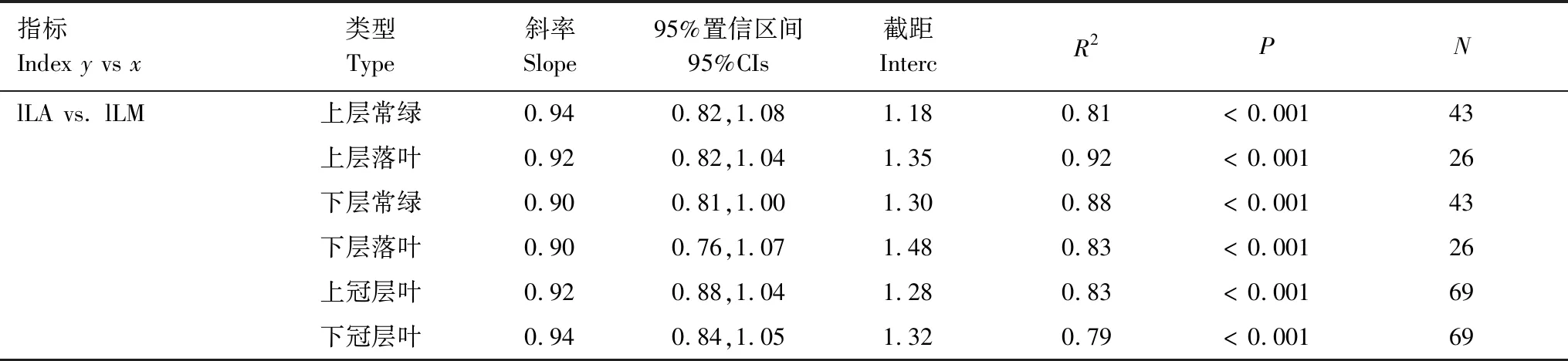
表4 不同冠层位置和生活型叶面积与生物量间的异速生长关系
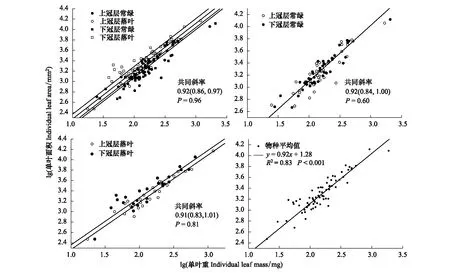
图3 单叶面积和单叶重异速生长关系Fig.3 The scaling relationships between the lndividual leaf area and lndividual leaf mass
3 讨论
木本植物在生长过程中会产生相对的冠层高度,不同生活型的物种在生存策略上存在差异,研究不同冠层高度和生活型植物叶面积-叶生物量之间的关系[28]。可以为叶的资源获取策略,生理协调和系统进化模式提供证据[29]。本研究表明冠层高度和生活型并未对当年生小枝的叶面积-叶生物量间的异速生长指数产生影响,但异速生长常数在不同冠层高度与生活型间存在差异。
3.1 不同冠层高度对叶面积-生物量关系的影响
本研究中,生活型相同的树种在不同冠层高度下当年生小枝叶面积-叶生物量无显著差异(P>0.05,图1)。即表明不同冠层高度并未对当年生小枝的叶面积和叶生物量产生影响。然而,Srikrishnah等[30]通过对斯里兰卡一种龙血树在不同高度光照水平下的研究中发现,处于不同高度光照水平下的叶面积和叶生物量存在显著差异(P<0.05)。原因可能是Srikrishnah等[30]的研究位于干旱的环境,水分是其生长的重要影响因素。不同的水分条件会改变植物的蒸腾耗水量,从而对叶面积产生影响,进而对叶面积和叶生物量在不同冠层高度产生影响。相反,本研究处于降水较多的亚热带地区,水分不是本区域植物生长的重要限制因子。另外,本研究发现落叶植物的比叶重在不同冠层高度存在显著差异(P<0.05,图1),常绿植物的比叶重无显著差异(P>0.05,图1)。然而Aclerly等[31]的研究发现,植物的LMA随着光照的增强而增多,但光照的有效性对单个物种的叶面积影响并不显著。本研究的原因首先可能是落叶树种在生长季更倾向于在生长季期间将生物量用于其叶面积的扩张,以得到更多的资源[32],而不同冠层高度的光照有效性导致落叶树种在上下冠层获取的资源存在差异。相反,常绿植物则倾向于更多的将生物量分配到支撑结构(根、茎)的生长,以便抵御非生长季外界恶劣的环境[33]。其次常绿和落叶植物树高和冠幅存在差异,导致在接收光照条件时可能无法严格体现冠层高度这一因素的限制。因此植物功能性状的平均值可能掩盖了性状变异在群落上的表现。
本研究中,89.9%(62/69)亚热带木本植物在不同冠层高度上具有显著的叶面积-叶生物量异速生长关系(图2)。一方面59.4%(41/69)的物种在ILA-ILM呈现出的异速生长指数与1.0无显著性差异(P>0.05图2),另一方面,24.6%(17/69)的木本植物叶面积-叶生物量异速生长指数显著小于(P<0.05,图2)1.0,5.8%(4/69)的木本植物叶面积-叶生物量异速生长指数显著大于(P>0.05图2)1.0。因此,研究结果总体上不支持个体水平的“收益递减”假说。本研究与姚婧等[34]对五角枫在幼树叶性状异速生长关系的结果一致。相反,Sun等[7]对中国东南武夷山不同海拔梯度和光照条件下竹子的叶大小和叶生物量之间的研究发现,海拔和光照条件未改变叶大小和叶生物量之间的异速生长关系(异速生长指数<1.0)。Pan等[14]对不同海拔植物叶大小和叶生物量关系进行分析发现,其叶大小与叶生物量的异速生长指数随着海拔的升高从<1.0向>1.0有规律变化。这些差异可能是两方面原因导致的首先在海拔梯度这一因素的影响下,植物的器官和生长可能受到环境因素影响[35],会使得植物的生物量分配产生变化。而本研究没有海拔的差异,虽然存在冠层高度的不同,但是并没有明显的改变植物的生境;另一方面,植物下冠层的叶片主要生活在光照不充足的环境条件下,为了更好的生长,植物可以将更多的生物量投资在叶片的扩大,以此增加光合有效面积来获取更多的资源[36]。而上冠层的叶片面临的环境胁迫增加(如,强风等),可能投入较多的生物量在起支撑作用的叶柄[37]。有研究表明,叶柄虽然在植物的叶中比例虽然较小,但却拥有很高的生物量分配[38-39]。因此叶面积与叶生物量的异速生长关系没有受到显著的影响。
3.2 不同生活型对叶面积-生物量关系的影响
本研究结果表明,江西亚热带不同生活型的木本植物在ILA-ILM二者关系上都拥有共同斜率,但在截距方面存在差异(图3)。落叶树种在不同冠层高度叶面积与叶生物量的截距存在差异(P<0.05),且下层落叶的截距大于上层落叶(图3)。常绿树种和落叶树种叶面积与叶生物量的截距也存在差异(图3a)。相反常绿树种的叶面积与叶生物量之间的截距不存在差异(P>0.05,图3)。另外本研究表明常绿植物和落叶植物的比叶重存在显著差异(P<0.05,图1),即常绿植物的比叶重显著大于落叶植物。这种结果的原因首先可能是常绿和落叶植物不同的生活史对策导致的。在整个生长季,常绿树种和落叶树种采取的生态策略不同,落叶植物倾向高生长效率的策略,而常绿植物倾向采取保守性的获取资源[40],这会影响常绿树种和落叶树种在生长速率和不同器官的生物量分配存在差异。其次可能是由于不同生活型植物的叶片对环境的响应机制不一样[41],植物LMA的变化主要是叶片厚度以及叶片密度的变化驱动[42]。常绿植物比落叶植物拥有更厚的叶片,叶片的厚度以及密度对植物叶片获取光资源存在限制[43-44],因此这些常绿植物和落叶植物之间生活史的差异,导致LMA存在显著差异。
另外本研究结果表明江西亚热带木本植物在不同生活型之间ILA-ILM的异速生长指数与1.0无显著性差异(P>0.05,表4),表明常绿植物和落叶植物并未对ILA-ILM的异速生长指数产生影响。60.5%(26/43)的常绿树种(图2)和57.7%(15/26)的落叶树种(图2)ILA-ILM呈等速生长关系。即随着叶生物量的增加,光合有效面积并没有降低。这与Pan等[14]的研究结果在中海拔地区保持一致。然而却与Milla和Reich等[12]的结果不一致,这可能由于本研究选取的是当年生小枝上的叶片,后者的研究则选取的成熟的叶片。叶片的大小具有差异,大叶往往能获得更多的光资源[45],但同时就需要在叶片的支撑结构需要投入更多的生物量,这就导致叶面积和叶生物量的投资比例失衡[46]。而小叶片只能通过其他部位(叶柄、叶角)和特征(叶形)来对光进行截留,以此活获得资源满足生长需要[47]。另外30.2%(13/43)(图2)的常绿树种和30.8%(8/26)(图2)的落叶树种ILA-ILM呈异速生长关系。这表明当年生小枝叶面积-叶生物量的异速生长关系在相同的生活型中具有物种特异性。原因可能是由于物种叶片细微结构(栅栏组织、气孔结构等)存在差异,随着当年生小枝叶片的生长,叶片形态改变的同时导致比叶重和光合速率产生变化[48]。从而在外界环境中获取资源适应生长。此外叶大小发生变化的时,叶片长宽比的变化能够明显影响叶面积-叶生物量之间的异速生长关系[49]。因此在相同生活型的物种之间,当年生小枝的叶面积与叶生物量之间的异速生长关系也会产生差异。
4 结论
研究结果表明,在江西亚热带常绿阔叶林森林生态系统中,不同冠层高度叶面积-叶生物量之间异速生长指数与1.0无显著差异。同样常绿树种和落叶树种中60.9%(42/69)木本植物叶面积-生物量间呈等速生长关系,不支持收益递减假说。表明冠层梯度和生活型并未显著影响江西亚热带林木本植物当年生小枝叶面积-叶生物量异速生长关系。研究结果可从不同冠层高度进一步了解植物的资源分配策略,进而丰富对植物个体生长发育对环境响应的理解。
