祁连山中部祁连圆柏年内径向生长对气候因子的响应
2020-11-24金敏艳李进军车宗玺张军周勾晓华
金敏艳,李进军,车宗玺,王 放,张军周,*,勾晓华
1 甘肃祁连山国家级自然保护区管理局, 张掖 734000 2 兰州大学资源环境学院西部环境教育部重点实验室, 兰州 730000
气候变化除表现出温度的快速升高外,区域降水模式也发生了显著变化,使干旱发生的频率及程度明显加剧[1]。树木生长对气候变化非常敏感[2],在一些高寒湿润地区,温度升高有利于树木径向生长加速[3]和生长季延长[4],使森林生产力增加[5]。但是温度的升高往往伴随着蒸散作用的增加,加剧一些区域的干旱程度,使树木生长速率减缓[6],甚至出现大面积的树木死亡现象[7]。因此,明确区域树木生长动态及其对气候变化的响应,有助于评估区域森林的变化规律及趋势,为森林保护和管理提供科学依据。
树木形成层活动监测是明确树木径向生长动态变化过程及其对气候因子响应的重要手段。已有研究表明,无论寒冷湿润地区(如欧洲阿尔卑斯山和北美高纬度地区)[8-9],还是温暖干旱地区(如欧洲阿尔卑斯山干旱沟谷和地中海地区)[10-11],温度对树木径向生长开始时间起着决定性作用。一些研究已经指出存在树木形成层活动开始时间的平均温度阈值(6—8℃)[12- 14]。然而,与生长开始时间不同,不同环境下影响树木生长结束时间的环境要素存在很大差异。比如,在寒冷湿润地区,树木径向生长结束时间主要受到温度的控制[9],但在其它北方山地森林生态系统中,生长季结束时间可能并不由温度控制[12],在低海拔区域[10]或极端干旱地区[15]也可能受到干旱的限制。
祁连山是我国东部湿润区、西北干旱区和青藏高寒区的过渡带和边缘区,是典型的寒冷干旱地区[16]。因其特殊的地理位置和地形特征,祁连山成为对气候变化最敏感的区域之一。过去50年,祁连山年平均气温升高速率为0.37℃/10 a,高达同期全球平均水平的2倍[17]。祁连圆柏(Juniperusprzewalskii)是我国特有树种,也是构成祁连山森林生态系统的主要树种,是祁连山生态屏障的重要组成部分,在维持森林生态系统稳定、涵养水源及缓解旱涝灾害等方面起着重要作用[18]。祁连圆柏生长对气候变化的响应及适应关乎我国西北地区森林健康状况和生态安全。尽管学者们在祁连山开展了一系列监测研究工作[19- 22],但对影响区域树木生长开始和结束时间的限制因子并没有达成共识。比如,一些研究指出水分对寒冷干旱地区祁连圆柏径向生长开始时间起着至关重要的作用,但对影响生长结束时间的限制因子并不明确[20-21]。相反,一些研究指出温度可能是影响该区域树木生长开始的主要因素,水分可能对径向生长结束时间起主要作用[6, 23]。因此,亟待进一步开展相关监测研究,明确祁连圆柏生长结束与气候因子的响应关系。
本研究通过在祁连山中部油葫芦沟设置监测样点,每两周连续采集微树芯,对祁连圆柏进行了连续两年的形成层活动监测研究,探讨了祁连圆柏年内径向生长动态及其对气候因子的响应,为区域森林保护和管理提供科学依据。
1 材料与方法
1.1 研究区概况和监测点设置
研究区位于祁连山中部黑河流域上游葫芦沟流域(99°46′E, 38°14′N;图1),海拔2960—4800 m[24]。区域植被分布呈明显的垂直分布特征,沿海拔从低到高分别为湿地、森林、灌丛、草甸、草原、寒漠、冰川等不同下垫面[25]。由祁连圆柏和青海云杉(Piceacrassifolia)两个主要树种组成的森林占总流域面积的0.7%[24],其中祁连圆柏主要分布于海拔3000—3800 m的阳坡和半阳坡区域,郁闭度较低,基本呈孤立木分布。我们于2012—2013年在海拔3250 m的区域设置了监测样点,随机选择5棵树龄相近、健康、直立、无明显干扰的祁连圆柏树木(树高:(10.7±2.2) m;胸径:(33.5±8.4) cm)进行监测研究。

图1 采样点位置Fig.1 Map of the monitoring site
1.2 气象要素
离监测点最近的气象站为祁连站(100°25′E, 38°18′N, 2787 m),距离监测点30 km。本研究使用了祁连站记录的1961—2015年的降水量、温度等日值气象数据。为了评估研究区不同监测年份的干旱程度,并充分考虑前两个月气候状况对当月树木生长的影响[26],本研究使用Thornthwaite法计算了区域1961—2015年以来3个月尺度的标准化降水蒸发指数(SPEI3),以量化两个观测年份的干旱程度[27]。
1.3 微树芯采集及室内实验
我们分别于2012年6—10月和2013年5—9月,每两周使用微树芯采样器(Trephor)在树木胸径处(1.3 m)采集1—2个微树芯,采集时每个微树芯之间的间隔至少大于3 cm,采集后用FAA 溶液迅速固定[28-29];在实验室内,采集的微树芯经甘油—乙醇溶液软化、不同浓度乙醇逐级脱水、二甲苯透明后用切片石蜡包埋,蜡块修整后用Leica RM2245石蜡切片机制备8—12 μm切片,切片经二甲苯脱蜡、复水、番红—固绿复染后封片[29];切片制备后,使用显微镜在正常光和偏振光下观察微树芯横切面的解剖结构,在不同的放大倍数(100、200、400倍)下进行拍照,然后用ImageJ软件处理和分析照片。
1.4 形成层物候及木质部分化过程的确定
形成层细胞排列整齐,细胞直径较小,细胞壁较薄,使用番红—固绿复染后呈蓝绿色。当至少观测到1个处于扩大阶段细胞时(扩大阶段的细胞直径为形成层细胞的2倍左右),形成层活动开始;当形成层不再分裂形成新的细胞时,细胞分裂结束。加厚阶段的细胞壁在偏光下明显发亮,可据此来判断。成熟细胞的液泡和原生质体退化,细胞壁被染色剂染成紫红色。所有细胞成熟时间为生长季结束时间。
在本研究中,我们统计了每棵监测样树的木质部分化各阶段发生的时间:细胞扩大阶段开始(1)和结束(2)的日期,细胞壁加厚阶段开始(3)和结束(4)的日期以及细胞成熟阶段开始(5)和结束(6)的日期,每个关键日期用序日(DOY)来表示。同时,对每棵样树木质部分化各阶段的细胞数量也进行了统计,每次统计时沿着径向进行3个重复并计算平均数。统计完成后对细胞数量进行标准化[30]:
nci=ncmi×rwm/rws
式中,nci为标准化的细胞数量,ncmi为实际测量的细胞数量,rwm为前一年所有样本的平均轮宽或细胞数量,rws为当前样本前一年的轮宽或细胞数量。
将5棵树的统计结果经算数平均后得到树木生长的关键日期和细胞数量,结果用平均值±标准误(Mean±SE)表示。本研究使用单因素方差分析确定木质部分化各阶段关键日期和细胞数量在不同监测年份间的差异。
本研究使用Gompertz函数拟合树木径向生长动态过程[31],确定细胞分裂速率及其发生的时间(拐点):
Y=Ae-e(β-κt)
式中,Y表示木质部细胞的累积量,t为时间,用序日(DOY)表示;A为上渐近线,表示形成层细胞分裂的木质部细胞总量(Ncell);β是x轴的截距参数,κ为变化速率参数。最大细胞分裂速率(Rmax)和曲线拐点 (tp)通过以下公式计算[31]:
tp=β/κ
Rmax=κA/e
2 结果
2.1 气象要素变化特征
根据祁连站1961—2015年的记录,研究区平均温度为1.17℃,降水量为410.2 mm。降水量年内分布极不均匀,5—9月降水量占全年总降水量的89.8%(图2)。研究区近50年来温度呈显著升高趋势,降水量也有显著增加的趋势,温度升高的速率为0.32 ℃/10 a,降水增加的速率为10.3 mm/10 a。SPEI3没有明显的变化趋势。2013年平均温度为2.38℃,比多年(1.17℃)和2012年平均温度(1.49℃)分别高1.21℃和0.89℃(表1)。尽管2013年的降水量(465.7 mm)高于2012年(395.8 mm),但2013年(-0.36)的SPEI3显著低于2012年(0.31);尤其在祁连圆柏生长季前期和主要生长期(3—6月),2013年SPEI3为-1.66,说明2013年生长季前期和主要生长期相对2012年显著干旱。

表1 研究区气候特征
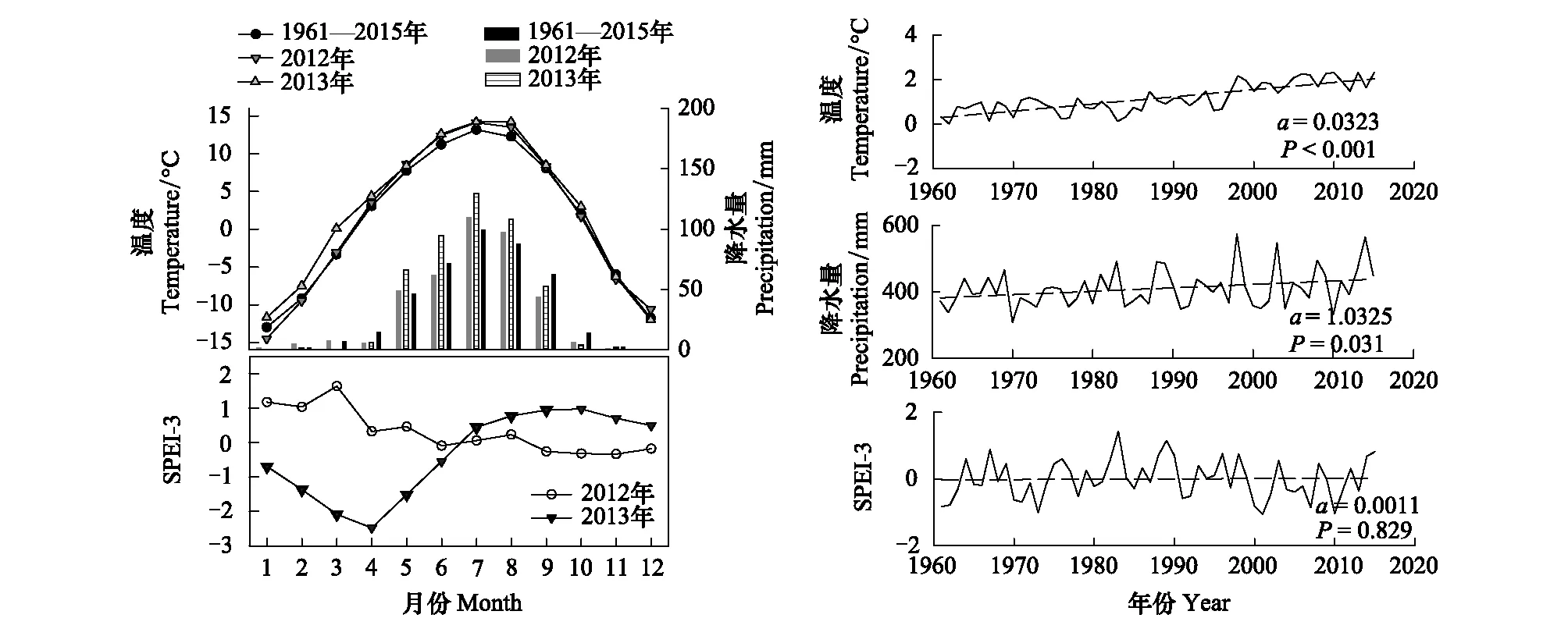
图2 研究区气候特征Fig.2 Climatic characteristics of the study area左图为祁连气象站记录的2012年、2013年及1961—2015年的温度、降水量及三个月尺度的标准化降水蒸发指数(SPEI3);右图表示1961—2015年祁连站的温度、降水、和SPEI3的变化趋势,虚线为趋势线
2.2 木质部分化各阶段开始和结束时间
由于2012年的第一次采样在6月上旬,此时已经观测到扩大细胞,因此无法判断形成层开始活动的具体时间(图3)。在2013年,第一个扩大细胞发现于5月31日(DOY152),说明此时祁连圆柏形成层开始活动(即生长季开始)。尽管细胞扩大阶段结束时间在两个监测年份中没有差异(P>0.05),但大多数监测树木的细胞扩大阶段结束时间在2013年(DOY190)比2012年(DOY197)早了一周左右。2012年细胞壁加厚阶段开始时间为6月26日(DOY178),比2013年的6月22日(DOY173)晚5 d,但在两个监测年份中没有显著差异(P<0.05)。细胞壁加厚阶段结束时间在2012年(8月8日,DOY221)和2013年(7月18日,DOY199)之间存在显著差异(P<0.05),2013年比2012年提前了22 d。木质化阶段开始和结束时间在两个监测年份中均存在显著差异(P<0.05)。其中2012年木质化阶段开始时间为7月23日(DOY205),2013年木质化开始时间为6月26日(DOY177),相差28 d;2012年木质化阶段结束时间(即生长季结束时间)为9月7日(DOY251),2013年木质化结束时间为8月15日(DOY227),相差24 d。2013年生长季持续时间为76 d(表2,图3)。

图3 2012和2013年木质部分化过程及细胞类型切片Fig.3 Types of xylem cells corresponding to different phases of xylem formation observed in 2012 and 2013 growing seasonCz:形成层区域Cambium zone;Ec:扩大细胞Enlarging cells;Wc:细胞壁加厚细胞Wall-thickening cells;Mc:成熟细胞Mature cells;刻度尺为100 μm
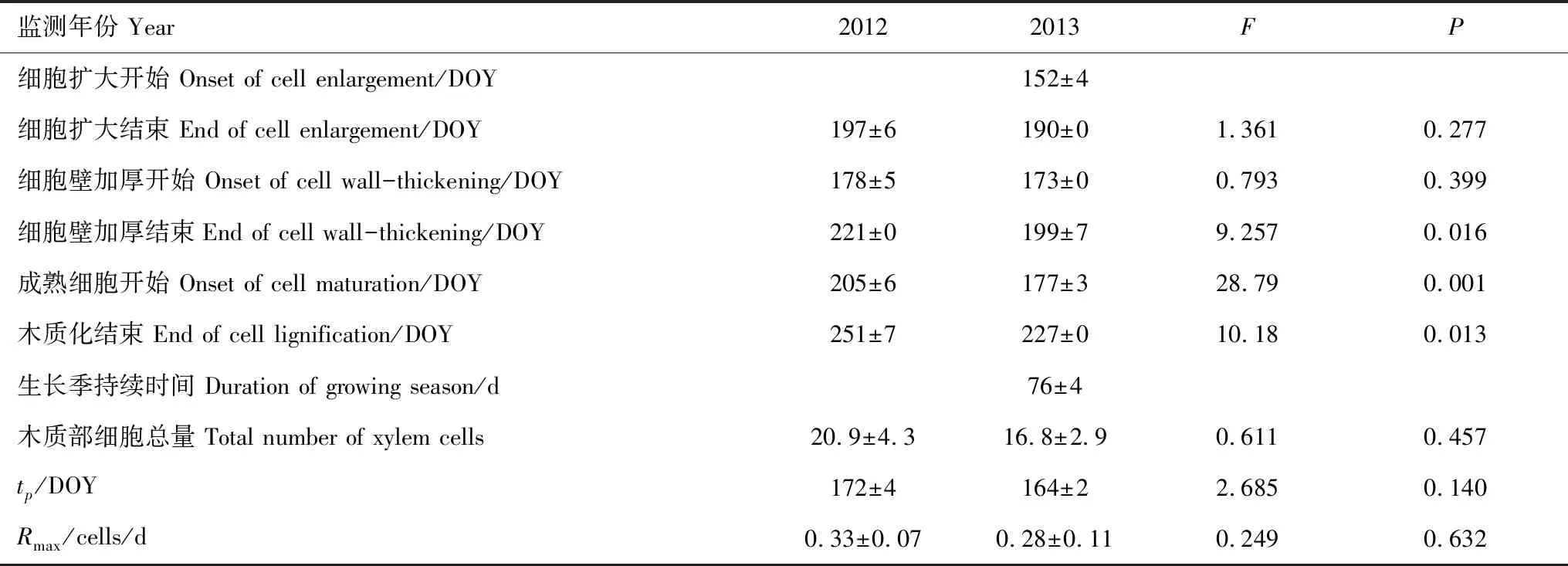
表2 不同监测年份树木形成层物候及木质部细胞产生量(平均值±标准误)
2.3 木质部细胞总量和生长速率
Gompertz函数很好地模拟了各监测树木在各监测年份的木质部细胞数量,拟合的解释量达到81%—99%(表3)。根据拟合结果,我们计算了2012和2013年各监测树木的木质部细胞最大分裂速率(Rmax),木质部细胞总量(Ncell)和最大速率发生时间(即拐点:tp)。通过对比发现,Ncell、Rmax和tp在两个监测年份中没有显著差异(P>0.05, 表2)。2012年,木质部细胞总量为20.9,细胞分裂速率为0.33 细胞/d。2013年的木质部细胞总量少于2012年,只有16.8个细胞,细胞分裂速率也小于2012年,为0.28 细胞/d。2012年拐点(最大生长速率)发生的时间为6月20日(DOY172),比夏至日只早1 d(DOY173);但2013年tp发生的时间为6月13日(DOY164),比夏至日早8 d(表2)。
2.4 木质部分化与气候因子的关系
监测结果表明,在温暖干燥的2013年,祁连圆柏木质部分化各阶段开始时间显著早于相对寒冷湿润的2012年,说明生长季前期的温度对木质部分化各阶段开始时间至关重要,而与水分状况没有关系(图4)。同时,祁连圆柏木质部分化各阶段结束时间在温暖干燥的2013年显著早于寒冷湿润的2012年,说明高温和干旱可能对祁连圆柏径向生长结束时间起着重要作用(图4)。此外,大多数树木的木质部细胞总量和生长速率在相对寒冷湿润的2012年大于相对温暖干旱的2013年,说明生长速率和木质部细胞总量可能与生长季早期干旱状况有关。

图4 2012和2013年木质部分化各阶段发生时间、木质部细胞数量与气候要素的关系Fig.4 Cambial phenology and wood production and associated climatic factors in 2012 and 2013

表3 运用Gompertz函数拟合的参数值及拟合解释量
3 讨论
3.1 形成层活动与气候因子的关系
尽管在2012年没有监测到祁连圆柏细胞扩大阶段开始的具体时间,但大多数树木细胞壁加厚阶段开始时间晚于2013年,同时所有树木的木质化阶段开始时间明显晚于2013年(两年相差达到28 d)。考虑到木质部分化各阶段开始时间具有较好的一致性[9],我们推测2012年细胞扩大阶段开始时间可能也晚于2013年。由气象记录可知,2012年为相对寒冷湿润年份,2013年为相对温暖干旱年份,暗示着2013年相对温暖的早春可能导致了树木生长季开始时间提前。在祁连山南坡,不同海拔梯度的监测研究表明祁连圆柏生长季开始时间随着海拔的升高明显延迟,说明温度差异是祁连圆柏生长开始时间的决定因素[6]。此外,很多基于控制实验[32- 35]和自然监测[4, 12, 14, 36]的研究也指出生长季早期升温会导致形成层活动开始时间提前。然而,一些学者在青藏高原东北部的监测表明径向生长开始时间受降水影响[20-21]。在本研究中,尽管2013年春季和夏季发生了严重的干旱,但细胞壁加厚和木质化阶段开始时间并没有晚于2012年,说明水分对形成层活动开始时间没有影响。综上所述,祁连山中部祁连圆柏形成层活动开始时间主要受温度影响。
寒冷环境中温度对生长季开始时间的重要作用已被大多数学者认可,但对决定生长季结束时间的气候因子还存在争议。在高纬度或高海拔湿润区域,研究指出温度不仅决定了生长季开始,对生长季结束也扮演着重要角色[4, 9, 13, 37]。然而在干旱半干旱区,树木生长结束与温度没有关系。比如,有研究指出低海拔树木生长结束时间与海拔决定的温度并没有关系,可能受到其它因素的控制[10]。同时,也有研究指出生长结束并不受外界环境的影响,可能受到树木本身的生理因素控制[38]。在本研究中,2013年(温暖年)的祁连圆柏细胞扩大、细胞壁加厚及木质化阶段结束时间明显早于2012年(寒冷年),说明较低的温度并不能使生长季结束时间提前,这与基于不同海拔梯度的树木生长监测研究结果一致[6]。相反,2013年的高温导致了夏季极端干旱事件的发生,对祁连圆柏径向生长结束时间产生了至关重要的影响,使其明显早于2012年。许多研究已经表明高温导致干旱胁迫对树木生长具有重要影响[2, 39-40]。我们在祁连山南坡的监测研究也表明水分胁迫对祁连圆柏径向生长结束时间具有重要影响[6]。此外,在北美南部干旱地区也有类似的研究结果[15]。因此,在干旱半干旱区,水分状况对生长季结束起着至关重要的作用。
3.2 树木年内径向生长对气候因子的响应
研究表明,在干旱地区,生长速率对木质部细胞总量起着决定作用[6, 41]。在干旱环境下,相对湿润年份能够增大形成层活动速率,产生较高的生长量[23]。在本研究中,我们发现在相对湿润的2012年,大多数监测树木的木质部细胞总量和生长速率大于相对干旱的2013年,支持了以上结论。监测表明生长季之前或生长季早期的降水对青藏高原东北部祁连圆柏的生长起着重要作用[21]。在祁连山北坡,祁连圆柏年内径向生长与干旱状况显著相关[19, 42]。此外,在祁连山及其周边地区,大多数基于祁连圆柏的树轮气候学研究结果也表明树轮宽度与生长季降水或干旱指数呈正相关[26, 43- 46],说明年际尺度的祁连圆柏树木生长也受到水分的限制。因此,生长季干旱状况是决定该区域祁连圆柏径向生长量最主要的气候因子。
一些研究认为,光周期和昼长也对形成层活动及径向生长量产生重要影响。比如,在寒冷环境中,树木形成层活动与昼长同步,在生长季早期快速分裂并在昼长最大的夏至日达到最大生长速率[38, 47]。本研究监测结果表明,2012年祁连圆柏细胞分裂最大速率发生的时间与夏至日吻合,说明光周期可能对树木生长产生一定的影响。然而,在2013年,祁连圆柏细胞分裂最大速率发生的时间比夏至日早了8 d,暗示昼长对树木径向生长的影响并不具有普遍性。同时,研究结果表明,祁连圆柏细胞分裂最大速率发生的时间在相对干旱的2013年明显早于相对湿润的2012年,进一步说明干旱对生长速率和木质部细胞总量具有重要影响。在相对湿润的2012年,由于干旱限制较弱,光周期对树木生长可能产生了一定的影响;但在干旱加剧的2013年,水分成为限制树木生长的主要限制因素,这与在祁连山其他区域的研究结果一致[23]。有研究已经指出,在干旱限制下,欧洲赤松(Pinussylvestris)的最大细胞分裂速率早于夏至日约4—6周[48],进一步证实了在干旱区,水分状况决定了最大径向生长速率及其发生的时间,进而决定了径向生长量。
4 结论
研究通过对祁连山中部祁连圆柏形成层活动和径向生长动态连续两年的监测研究,得到了以下主要结论:
(1)祁连圆柏的形成层活动对外界环境非常敏感,木质部分化各阶段发生的时间在不同监测年份中存在显著差异。其中,木质部分化各阶段开始时间在相对温暖的2013年显著早于相对寒冷的2012年;各阶段结束时间在相对干旱的2013年均早于相对湿润的2012年,说明温度和有效水分分别对祁连圆柏的生长开始和结束时间起着至关重要的作用。
(2)祁连圆柏的木质部细胞总量和细胞分裂速率在相对湿润的2012年大于相对干旱的2013年,说明水分对区域树木生长量及生长速率具有重要作用。这与年际尺度的树轮—气候响应结果一致。
尽管温度升高会导致寒冷地区的树木生长开始时间提前,生长季持续时间延长,但在干旱地区,升温是把双刃剑,除了导致生长季提前外,也导致了夏季干旱程度的加剧,使生长季提前结束。同时,升温导致的干旱胁迫对干旱区树木的生长速率产生了重要影响。由于干旱地区的树木径向生长量主要与生长速率有关,温度升高势必会导致该区域树木径向生长量降低。因此,在未来气候变暖情境下,升温导致的干旱加剧可能会使区域树木的生长速率降低,进而使其径向生长量下降。
