姜黄素调节SREBP和FAS改善非酒精性脂肪性肝病的研究
2020-11-23陈仲生阎文柱
陈仲生?阎文柱


【摘要】目的 探究姜黄素对非酒精性脂肪性肝病(NAFLD)的作用和可能机制。方法 18只SD大鼠随机分为对照组、模型组和姜黄素组,每组各6只。模型组和姜黄素组予以高脂高胆固醇食物10 ml/kg灌胃,对照组给予等量生理盐水灌胃。建模成功后,姜黄素组每日给予姜黄素100 mg/kg灌胃,对照组和模型组给予等量的羧甲基纤维素钠。治疗4周后,检测大鼠血清ALT、AST、甘油三酯(TG)和总胆固醇(TC)水平,HE染色观察大鼠肝脏组织病理学变化,免疫组织化学法检测大鼠肝组织固醇调节元件结合蛋白(SREBP)和脂肪酸合成酶(FAS)蛋白的表达情况,蛋白免疫印迹法检测沉默调节蛋白6(Sirt6)和磷酸化腺苷酸活化蛋白激酶(p-AMPK)蛋白表达变化。结果 模型组ALT、AST、TG、TC、SREBP和FAS蛋白表达均高于对照组(P均< 0.05);姜黄素组ALT、AST、TG、TC、SREBP和FAS蛋白表达均低于模型组(P均< 0.05)。模型组Sirt6、p-AMPK蛋白表达均低于对照组(P均< 0.05);姜黄素组Sirt6、p-AMPK蛋白表达均高于模型组(P均< 0.05)。结论 姜黄素可以减轻大鼠NALFD的脂肪沉积和肝损伤,可能与Sirt6、p-AMPK的表达上调相关。
【關键字】非酒精性脂肪性肝病;固醇调节元件结合蛋白;脂肪酸合成酶;姜黄素;
沉默调节蛋白6;磷酸化腺苷酸活化蛋白激酶
Curcumin alleviates non-alcoholic fatty liver disease by regulating SREBP and FAS Chen Zhongsheng, Yan Wenzhu. Department of Gastroenterology, the Third Affiliated Hospital of Jinzhou Medical University, Jinzhou 121000, China
Corresponding author, Yan Wenzhu
【Abstract】Objective To explore the regulatory mechanism of curcumin in the treatment of non-alcoholic fatty liver disease (NAFLD). Methods Eighteen male SD rats were randomly divided into the control, model and curcumin treatment groups, 6 rats in each group. In the model and curcumin treatment groups, NAFLD rat models were established by gavage with high-fat and high-cholesterol diet, and an equivalent amount of normal saline was given in the control group. After the model establishment, the rats were given with gavage with 100 mg/kg curcumin in the curcumin treatment group, and an equivalent amount of sodium carboxymethyl cellulose was given in the control and model groups. At 4 weeks after treatment, the serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), triglyceride (TG) and total cholesterol (TC) were detected. Pathological changes of liver tissues were observed by HE staining. The expression levels of sterol regulatory element-binding protein (SREBP) and fatty acid synthase (FAS) proteins in liver tissues were characterized by immunohistochemical staining. The expression levels of silent information regulator 6 (Sirt6) and phosphorylated adenylate-activated protein kinase (p-AMPK) proteins were measured by Western blot. Results In the model group, the expression levels of ALT, AST, TG, TC, SREBP and FAS proteins were significantly higher compared with those in the control group (all P < 0.05). In the curcumin treatment group, the expression levels of ALT, AST, TG, TC, SREBP and FAS proteins were significantly lower compared with those in the model group (all P < 0.05). In the model group, the expression levels of Sirt6 and p-AMPK proteins were significantly lower than those in the control group (both P < 0.05). In the curcumin treatment group, the expression levels of Sirt6 and p-AMPK proteins were significantly higher than those in the model group (both P < 0.05). Conclusion Curcumin can alleviate lipid deposition and hepatic injury in NALFD rats probably by up-regulating the expression levels of Sirt6 and p-AMPK.
【Key words】Non-alcoholic fatty liver disease;Sterol regulatory element-binding protein;
Fatty acid synthetase;Curcumin;Silent information regulator 6;
Phosphorylated adenylate-activated protein kinase
非酒精性脂肪性肝病(NAFLD)是指排除长期大量饮酒和其他明确的肝脏损伤因素所引起的脂质在肝细胞中储积为病理改变的肝脏代谢性疾病。近年来,NAFLD逐渐成为全球最常见的慢性肝病[1]。目前关于NAFLD发病机制尚不十分明确,现被公认的是“二次打击学说”。其可能的机制有氧化应激、内质网应激、胰岛素抵抗、炎症反应以及自噬与DNA 损伤等[2-4]。固醇调节元件结合蛋白(SREBP)和碳水化合物反应原件结合蛋白是调节肝脏脂质合成的關键性基因,进而参与调控肝脏脂质代谢过程[5-6]。因此,通过寻找有效药物抑制SREBP表达,对于改善NAFLD脂质代谢紊乱至关重要。
姜黄素是一种多酚类物质,从植物姜黄中提取。研究发现,姜黄素具备抗炎、抗氧化、抗肿瘤、抗自噬、抗胰岛素抵抗等作用,且对于糖尿病、COPD、动脉粥样硬化、自身免疫性疾病的治疗都有确切的效果。既往研究发现,姜黄素能够通过调节SREBP,进而参与肝脏脂质代谢[7]。在NAFLD大鼠中,降脂颗粒可以通过下调肝X受体α和SREBP-1c的表达,调节脂肪酸代谢[8]。但是有关姜黄素对SREBP调节的分子机制尚不完全清楚。因此,本实验研究通过构建NAFLD型大鼠模型,探究姜黄素对SREBP的分子调控机制。
材料与方法
一、材 料
1.实验动物
实验选取雄性SPF级别,体质量为180 ~ 200 g的SD大鼠共18只(购于锦州医科大学动物中心)并通过动物伦理委员会批准(批号:20190816),实验动物饲养于湿度在30% ~ 70%、温度维持在25 ℃的动物房内并采取昼夜交替光照处理。
2.主要实验药品及试剂
胆固醇(万邦实业有限公司,批号:2017 031625),姜黄素(上海瑞永生物科技有限公司), SREBP一抗和脂肪酸合成酶(FAS)一抗抗体(武汉Proteintech),沉默调节蛋白6(Sirt6)一抗(武汉博士德公司),腺苷酸活化蛋白激酶(AMPK)一抗(美国Cell Signaling Technology),β-actin一抗(江苏碧云天公司) PVDF膜(Millipore公司),SDS-PAGE凝胶(武汉博士德公司),一抗、二抗稀释液(江苏碧云天公司)。
3.主要实验仪器
纯水仪器(湖南湘仪公司),酶标仪(江苏碧云天公司),组织切片机(武汉赛默飞公司),荧光倒置显微镜(日本Olmpus公司),凝胶电泳装置(北京六一生物科技有限公司),显影仪(上海西唐生物科技有限公司),脱色摇床(上海鲁硕实业有限公司),高速离心机(上海卢湘仪离心机仪器有限公司)。
二、方 法
1.造模及分组
将18只实验大鼠随机分为3组,每组各6只,即为对照组、模型组、姜黄素组。模型组和姜黄素组予以高脂高胆固醇食物10 ml/kg灌胃,对照组给予生理盐水10 ml/kg灌胃。其中高脂高胆固醇食物的配制为60%猪油、10%胆固醇及30%玉米面加200 ml食用水,熬制成糊状,放入37℃恒温箱内,对照组给予常规饲料饲养。3个月后随机处死模型组和姜黄素组各1只大鼠,肝脏病理见肝脂肪细胞变性表明建模成功。姜黄素组给予姜黄素100 mg/kg按每日1次给药;对照组和模型组给予等量的生理盐水灌胃;持续给药4周。隔夜禁食水,次日清晨予大鼠腹腔注射1%戊巴比妥钠麻醉,称重、解剖并左心室采血,取部分肝脏组织浸泡于4%的多聚甲醛,经脱水、石蜡包埋后行HE染色和免疫组织化学检测。
2.相关指标检测
按照试剂盒说明书进行大鼠血清中ALT、AST、甘油三酯(TG)、总胆固醇(TC)含量测定。HE染色:4%的多聚甲醛固定大鼠肝脏组织,石蜡包埋后切片,之后行HE染色操作。免疫组织化学染色:①石蜡切片60℃下烘烤1 h;②脱蜡至水,二甲苯处理3次,每次5 min,无水乙醇处理3次,每次10 min,95%乙醇处理2次,每次10 min,dd H2O洗2次,每次5 min,0.01 mmol/L磷酸盐缓冲液(PBS)洗3次,每次10 min;③消除过氧化物酶体,3% H2O2溶液孵育10 min,0.01 mmol/L PBS洗3次,每次5 min;④微波抗原修复,枸橼酸钠缓冲液(pH 6.0)250 ml置于微波炉中450 W预热5 min,将组织放入缓冲液中,微波250 W加热10 min,冷却至室温,约耗时40 min,双蒸水洗3次,每次5 min,PBS洗3次,每次5 min;⑤封闭,利用5%的BSA封闭1 h;⑥打孔透膜,利用0.3%的Triton X-100孵育10 min;⑦一抗孵育,4℃湿盒孵育SREBP和FAS抗体过夜;⑧二抗孵育;⑨二氨基联苯胺(DAB)显色、苏木素复染,梯度乙醇脱水,二甲苯透明;⑩中性树胶封片和显微镜下观察结果。
蛋白免疫印迹法:①组织碾磨,将处理完成的各组大鼠肝脏组织取出,采用PBS清洗后并用手术剪刀剪取少许肝脏组织放置于组织碾磨管内,按照1∶5的比例加入组织裂解液后放置于组织碾磨仪中充分碾磨3 ~ 5 min;②离心,4℃以
12 000 rpm离心15 min,取组织上清液;③蛋白质浓度测定,利用二辛可宁酸(BCA)法进行蛋白质浓度定量测定;④凝胶电泳,按照上样量为40 μg
進行蛋白凝胶电泳120 min;⑤转膜,依据蛋白质marker指示进行切胶,并放置聚偏二氟乙烯(PVDF)膜进行转膜约90 min;⑥封闭,5%的脱脂牛奶封闭1 ~ 2 h;⑦孵育一抗,洗脱完成,孵育对应的一抗过夜;⑧孵育二抗,室温条件下,二抗孵育约2 h;⑨显影,将孵育完成的PVDF膜使用吐温-20的PBS洗涤2 ~ 3次,每次5 min,滴加显影液显影并拍照。
三、统计学处理
采用SPSS 18.0进行数据分析。正态分布计量资料采用表示,多组间比较采用单因素方差分析,两两比较采用LSD-t检验。P < 0.05为差异有统计学意义。
结果
一、姜黄素对大鼠血清生化指标的影响
与对照组相比,模型组ALT、AST、TG和TC增高,差异均有统计学意义(P均< 0.05);与模型组相比,姜黄素组ALT、AST、TG和TC均降低(P均< 0.05)。
二、HE染色观察大鼠肝脏组织形态学变化
与对照组相比,模型组肝小叶内弥漫性分布着空泡细胞,空泡大小不一,肝小叶结构尚存,边界模糊,有淋巴细胞分布于肝脏。相比于模型组,姜黄素组的肝小叶内空泡细胞明显减少,肝小叶结构清晰,肝脏内的空泡细胞数量最少,见图1。
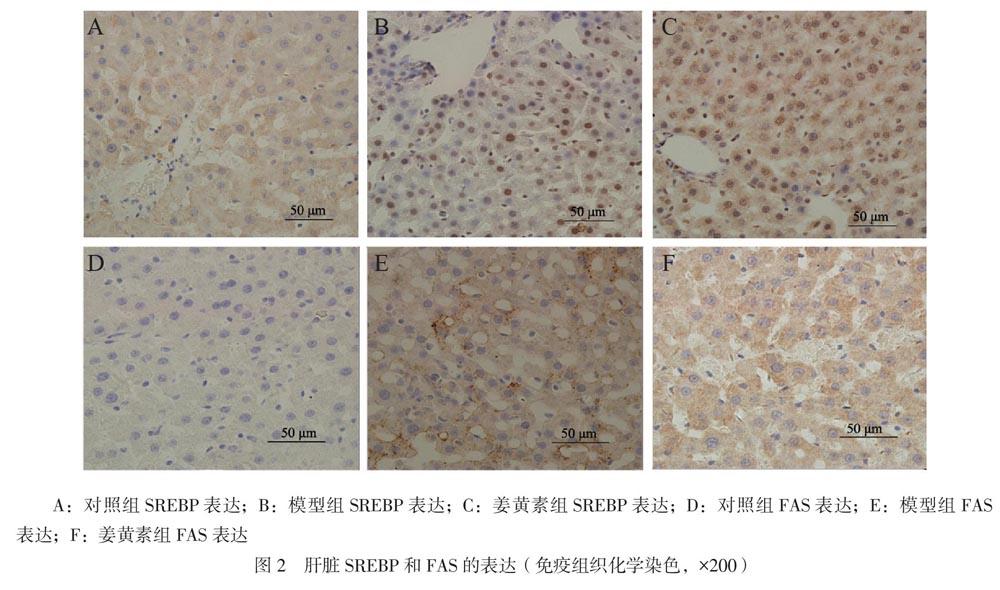
三、免疫组织化学法检测大鼠肝脏组织中的SREBP和FAS表达
SREBP和FAS的定位都位于细胞质内:与对照组相比,模型组SREBP和FAS的表达增加;与模型组相比,姜黄素组SREBP和FAS的表达减少,见图2。
四、姜黄素对大鼠肝脏Sirt6和p-AMPK表达的影响
3组肝脏组织Sirt6和p-AMPK蛋白表达比较差异均有统计学意义(FSirt6 = 12.83,PSirt6 = 0.001;Fp-AMPK = 16.67,Pp-AMPK< 0.001),相比于对照组,模型组Sirt6和p-AMPK蛋白表达降低(P均< 0.05);与模型组相比,姜黄素组Sirt6和p-AMPK蛋白表达升高(P均< 0.05),见图3。
讨论
NAFLD是以弥漫性肝细胞脂肪病变而导致的一种病理性临床病理综合征。NAFLD是一种进展性疾病,其发病机制尚不明确。既往研究发现胰岛素抵抗、脂质代谢异常、炎性细胞因子和氧化应激所致肝细胞凋亡等因素是NAFLD发生和发展的主要原因[9]。现有研究者提出在NAFLD的“二次打击学说”中,初次主要是胰岛素抵抗所致,胰岛素抵抗主要通过促使外周脂解增加和高胰岛素血症引起肝细胞脂肪蓄积,并增加了对内、外源性损害因子敏感性。二次打击主要为反应性氧化代谢产物、脂肪细胞因子、肠应激和肠道菌群内毒素等各种损害因子诱导活动UCP-2、FAS配体等,进而使得脂肪变性的肝细胞出现炎症、坏死等一系列病理生理学改变[10-12]。
NAFLD是指中性脂肪在肝细胞内以脂滴或者脂质囊泡的形式在胞质中过度积累。在诸多研究中发现总胆固醇在NAFLD是非酒精性脂肪性肝炎形成和发展中所起的脂毒性作用尚不完全清楚。因此,通过研究NAFLD中胆固醇合成和代谢的相关基因显得尤为重要。SREBP属于螺旋-环-螺旋-亮氨酸拉链(bHLH-ZIP)结构家族,是一种节脂质合成的转录因子家族,是由SREBP 1a、SREBP 1c 、SREBP 2三种亚型构成[13-14]。多项研究均指出SREBPS在NAFLD的发生发展中起重要作用。近年来诸多研究也发现姜黄素可以通过降低TG、TC改善NAFLD。研究发现,姜黄素可通过下调节SREBP和FAS表达,减少脂质在肝脏内沉积,降低脂毒性对肝脏损害,进而保护肝脏细胞,使得病情得以逆转,从而治疗NAFLD[15]。对此本研究初步研究发现,相比于模型组,通过姜黄素大鼠灌胃处理的NAFLD组SREBP和FAS的表达明显下降,AST、ALT、TG和TC水平均明显降低。上述结果表明,姜黄素可能通过调节SREBP和FAS,从而降低TG、TC等合成,改善NAFLD。那么姜黄素是通过何种分子机制调控SREBP表达?
Sirt6是Sirtuins家族成员之一,具有高度保守的NAD+依赖性的去乙酰化酶,可能通过调节内皮一氧化氮合酶而发挥抗心肌肥大的作用[16]。以往研究发现,Sirt6基因缺失是导致TG累积的主要因素,与脂肪性肝脏疾病密切相关[17]。在高脂喂养的小鼠中Sirt6的表达显著降低,通过过表达Sirt6可以降低小鼠血液脂质含量,减少肝脏内脂肪的积累[18]。亦有研究证实,Sirt6可以通过增加AMP/ATP比例,活化AMPK信号通路,进而调控SREBP蛋白表达[19]。因此,在本实验研究中,我们通过蛋白免疫印迹法检测大鼠肝脏组织中Sirt6和p-AMPK蛋白表达。结果发现,相比于对照组,模型组Sirt6和p-AMPK蛋白表达均明显降低,给予姜黄素处理可以促进Sirt6和p-AMPK蛋白表达,进而改善NAFLD。综上所述,姜黄素可以减轻大鼠NALFD的脂肪沉积和肝损伤,可能与Sirt6、p-AMPK的表达上调相关。以上研究结果为姜黄素治疗NAFLD提供了新的理论依据。
参 考 文 献
[1] Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology,2016,64(1):73-84.
[2] Kim SB, Kang OH, Lee YS, Han SH, Ahn YS, Cha SW, Seo YS, Kong R, Kwon DY. Hepatoprotective effect and synergism of bisdemethoycurcumin against MCD diet-induced nonalcoholic fatty liver disease in mice. PLoS One,2016,11(2):e014 7745.
[3] Macías J, Mancebo M, Merino D, Téllez F, Montes-Ramírez ML, Pulido F, Rivero-Juárez A, Raffo M, Pérez-Pérez M, Merchante N, Cotarelo M, Pineda JA; Spanish AIDS Research Network-HEP09 Study Group. Changes in liver steatosis after switching from efavirenz to raltegravir among human immunodeficiency virus-Infected patients with nonalcoholic fatty liver disease. Clin Infect Dis,2017,65(6):1012-1019.
[4] Sun LY, Yang YS, Qu W, Zhu ZJ, Wei L, Ye ZS, Zhang JR, Sun XY, Zeng ZG. Gut microbiota of liver transplantation recipients. Sci Rep,2017,7(1):3762.
[5] 耿得珍, 卜亚茹, 焦波. 潜在的代谢性疾病治疗靶点——固醇调节元件结合蛋白.中国药学杂志, 2019, 54(15):1205-1210.
[6] Horton JD, Goldstein JL, Brown MS. SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J Clin Invest,2002,109(9):1125-1131.
[7] 韩晓群, 林剑国, 杨婧. 姜黄素对活化肝星状细胞内脂质水平的影响及其机制. 解放军医学杂志, 2018, 43(12):1019-1023.
[8] 杨丽丽, 王淼, 柳涛, 宋海燕,励冬斐,郑培永,刘平,季光. 降脂颗粒对非酒精性脂肪性肝病大鼠肝X受体α和固醇调节元件结合蛋白1c表达的影响. 中西医结合学报, 2011, 9(9):998-1004.
[9] 肖阳, 侯云鹤,尹鑫,康凤,李树德,杨世昆,陶建平.人参皂苷Rg1干预非酒精性脂肪肝模型大鼠肝细胞的凋亡.中国组织工程研究,2019,23(3):384-390.
[10] 张姗姗, 王来友. 非酒精性脂肪性肝病药物治疗学前沿与展望. 世界华人消化杂志,2019,27(2):73-79.
[11] Satapathy SK, Sanyal AJ. Epidemiology and natural history of nonalcoholic fatty liver disease. Semin Liver Dis,2015,35(3):221-235.
[12] Musso G, Cassader M, Rosina F, Gambino R. Impact of current treatments on liver disease, glucose metabolism and cardiovascular risk in non-alcoholic fatty liver disease (NAFLD): a systematic review and meta-analysis of randomised trials. Diabetologia,2012,55(4):885-904.
[13] Brown MS, Goldstein JL. The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell,1997,89(3):331-340.
[14] Goldstein JL, Rawson RB, Brown MS. Mutant mammalian cells as tools to delineate the sterol regulatory element-binding protein pathway for feedback regulation of lipid synthesis. Arch Biochem Biophys,2002,397(2):139-148.
[15] Engelking LJ, Cantoria MJ, Xu Y, Liang G. Developmental and extrahepatic physiological functions of SREBP pathway genes in mice. Semin Cell Dev Biol,2018,81:98-109.
[16] 黄小阳,李卓明,刘志平,刘培庆. SIRT6通过调节eNOS抑制心肌肥大的机制研究.中山大学学报(医学科学版), 2015,36(3):338-345.
[17] Xiong X, Yu J, Fan R, Zhang C, Xu L, Sun X, Huang Y, Wang Q, Ruan HB, Qian X. NAMPT overexpression alleviates alcohol-induced hepatic steatosis in mice. PLoS One,2019,14(2):e0212523.
[18] Kim HS, Xiao C, Wang RH, Lahusen T, Xu X, Vassilopoulos A, Vazquez-Ortiz G, Jeong WI, Park O, Ki SH, Gao B, Deng CX. Hepatic-specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis. Cell Metab,2010,12(3):224-236.
[19] Kanfi Y, Peshti V, Gil R, Naiman S, Nahum L, Levin E, Kronfeld-Schor N, Cohen HY. SIRT6 protects against path-ological damage caused by diet-induced obesity. Aging Cell,2010,9(2):162-173.
(收稿日期:2020-06-11)
(本文編辑:杨江瑜)
