不同物候期茶树品种(系)叶绿素荧光特性研究
2020-11-18唐敏翟秀明李解邬秀宏侯渝嘉
唐敏 翟秀明 李解 邬秀宏 侯渝嘉
摘 要 采用PAM-2500便携式调制叶绿素荧光仪测定不同物候期类型的5个茶树品种(系)叶绿素荧光参数,分析不同物候期类型茶树的荧光参数特征,并比较各参数的品种(系)间差异。结果显示,茶树对吸收的光能,有79.4%~83.1%通过调节性或非调节性机制以热量形式耗散掉,20.6%~16.9%用于光化学反应。物候期较早的品种光合中心活性较强,光响应曲线斜率较大,半饱和光强较小,在作用光光强一定时实际光合量子产量高于物候期较晚的品种(系),表现出Y(II)、Alpha、ETRmax等参数显著高于晚生品种(系),Fo显著低于晚生品种(系)的荧光特性,且与茶树物候期的先后具有较高一致性。
关键词 茶树;品种;物候期;叶绿素荧光;光响应曲线
中图分类号:S184 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2020.28.003
茶树是我国重要的经济作物,其经济价值与采收时间具有直接的联系。茶树光量子利用规律是茶树光合作用研究的基础,对早生茶树资源的鉴定与筛选和种质创新具有重要意义。叶绿素荧光技术是以植物体内光系统Ⅱ(PSⅡ)中叶绿素a的荧光为天然探针,研究和探测植物光合生理状况的新型植物活体诊断技术,较表观性的气体交換指标更具反映内在性的特点[1-3],目前,该技术以其方便、快捷、无损的特点,已经在光、温度、干旱等逆境生理研究中得到广泛应用[4-6]。
本研究通过对物候期具有明显差异的5个茶树品种(系)的叶绿素荧光参数进行测试分析,旨在探讨其物候期与主要荧光参数间的关系及品种(系)间差异,了解不同物候期茶树品种(系)的光化学启动特性,为早生茶树资源的早期鉴定和早生茶树种质创新提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为生长健康,无病虫害的福鼎大白茶、渝茶3号、渝茶4号、乌牛早和品系0301的5年生成年茶树,选择当年生成熟叶片作为测定对象。
1.2 试验地点
试验在重庆市农业科学院茶叶研究所试验茶园进行,地理位置为北纬29°75′、东经105°71′,海拔440 m,属亚热带季风性湿润气候。2017—2019年平均年积温4 787.20 ℃,平均年日照时长809.13 h。
1.3 荧光参数测定
采用PAM-2500便携式调制叶绿素荧光仪测定荧光参数。测定前,将叶片进行充分暗适应(20 min以上),待叶片暗适应后,照射检测光,测定初始荧光Fo。然后加饱和脉冲光,测定最大荧光Fm。打开测量光和光化光,每隔10 s照射1次饱和脉冲光,测量光下最大荧光,采用Eilers and Peeters[Ecological Modelling 42(1988) 119-215]的公式拟合光响应曲线。所有参数测定均重复5次取平均值。
1.4 物候期调查
物候期调查所选芽为剪口下第一个芽。各目标品种(系)固定观察10个芽,每隔1~2 d观察1次,以样本数的30%为判断不同时期的标准,参见《茶树种质资源描述规范和数据标准》[7]。
1.5 数据处理
数据整理和统计分析采用Microsoft Excel和SPSS Statistics 22进行。
2 结果与分析
2.1 不同茶树品种的物候期
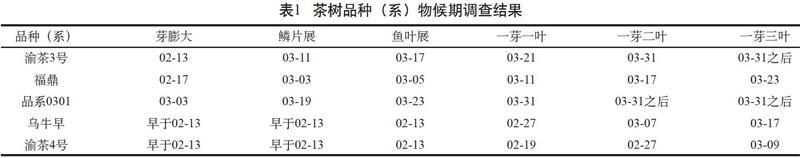
2017—2019年春季,按照《茶树种质资源描述规范和数据标准》分别对渝茶3号、渝茶4号、福鼎大白茶、乌牛早和品系0301等5个品种(系)在重庆永川地区的物候期进行了调查,调查结果列于表1。
结果显示,5个品种(系)按达到一芽一叶期的先后时间顺序为渝茶4号→乌牛早→福鼎→渝茶3号→品系0301,其中乌牛早、渝茶4号一芽一叶开采时间分别早于福鼎大白茶12 d、20 d,属于特早生品种,渝茶3号、品系0301分别晚于福鼎10 d、20 d,属于中生品种和晚生品种。
2.2 不同物候期茶树品种(系)叶片Fo、Fm、Fv和Fv/Fm的差异
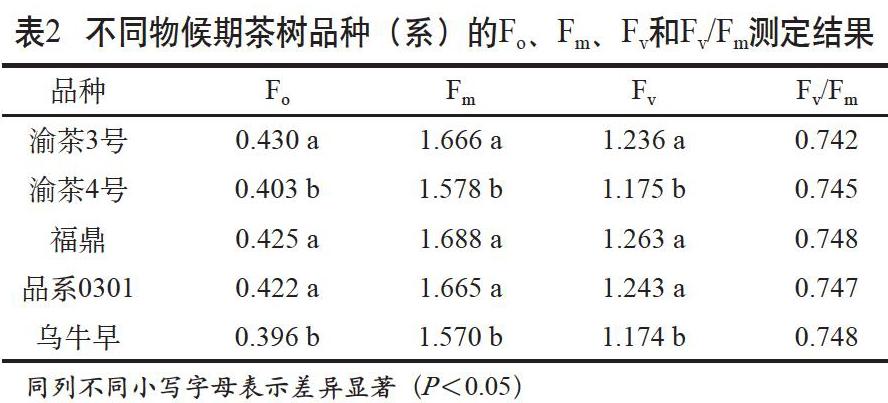
Fm是暗适应后叶片光合机构全部PSII中心都关闭时的荧光强度,这时荧光产量最大,反映了叶片光合中心的全闭合状态。Fo是暗适应后光合机构全部PSII中心都开放时的荧光强度,来自于天线叶绿素Chla,反映了叶片对光的全开放状态,与光合中心活性有关[8-9]。其差值Fv反映了叶片光合中心进行光化学反应的“库”或“能力范围”[10]。Fv/Fm反映的是植物对其生境长期适应的机制,正常生长植物的Fv/Fm相对稳定。当环境变化对植物造成胁迫或伤害后,Fv/Fm会随之降低[11-12]。不同物候期茶树品种(系)的Fo、Fm、Fv和Fv/Fm测定结果见表2。
结果显示,所试5个品种(系)Fv/Fm差异不显著,说明参试材料生长正常或受环境影响程度一致。Fo、Fm和Fv因品种(系)不同而表现出明显差异,其中渝茶4号和乌牛早的Fo、Fm、Fv显著低于其他3个参试品种(系),表明这2个品种光合中心活性高,但其光合中心进行光化学反应的“库”较小。
2.3 不同物候期茶树品种的能量分配差异
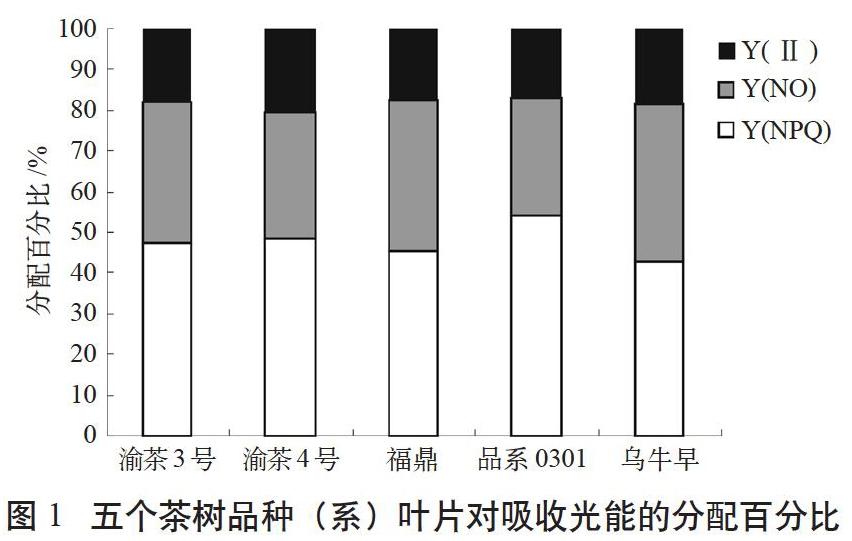
由于叶片接收的光能主要分为光化学能、热耗散和荧光3个部分被转化或耗散。这三者之间具有“光化学能+热耗散+荧光产量=1”的关系,即Y(II)+Y(NO)+Y(NPQ)=1。据此对5个品种的光能利用和分配进行了测定和分析。分别测定了以上5个品种的调节性能量耗散的量子产额Y(NPQ)、非调节性能量耗散的量子产额Y(NO)、实际光量子产量Y(II)(见图1)。
Y(NPQ)是PSII的调节性能量耗散的量子产量。若Y(NPQ)较高,一方面表明植物接受的光强过剩,另一方面则说明植物仍可通过调节过剩光能,将其转化为热而耗散,从而保护自身[13]。Y(NO)是光保护的重要指标,是PSII的非调节性能量耗散的量子产量。Y(NO)较高时,表明光化学能量转换和保护性的调节机制不足以将植物吸收的光能完全消耗掉[14],说明入射光强超过了植物能接受的程度,这时,植物可能已经受到损伤,或者将要受到损伤。
由图1可知,所示5个品种(系)茶树对吸收的光能,有79.4%~83.1%通过调节性或非调节性机制以热量形式耗散掉,20.6%~16.9%的能量用于光化学反应,推动自身的碳氮循环和生理活动,但不同品种对能量的分配比例各有不同。渝茶4号Y(II)最高,品系0301和福鼎最低,且与渝茶4号差异显著,乌牛早、渝茶3号居中,为过渡类型,与Y(II)最高和最低的品种均未达到统计学上的显著差异水平。所试5个品种(系)光合量子产量由大到小的顺序为渝茶4号>乌牛早>渝茶3号>福鼎>品系0301,这与一芽一叶物候期观察结果基本一致。
2.4 不同物候期茶树品种的光响应曲线
通过拟合光响应曲线(RLC)能反应不同光合有效辐射下叶片通过PSII的电子传递速率。本试验连续测定了叶片在0、13、30、55、87、127、178、235、303、390、503和640 μmol·m-2·s-1共12个光合有效辐射下叶片的电子传递速率,并采用Eilers and Peeters(1988)的公式ETR=PAR/(a·PAR2+b·PAR+c),对光响应曲线进行拟合,测定及拟合结果见图2。
结果显示,不同物候期茶树品种(系)在同样环境条件下,各品种(系)通过PSII的电子传递速率(ETR)随着光强的增加而不断增加,当光强增大到400~600μmol·m-2·s-1时,ETR增速减缓直至不再增加。所试5 个茶树品种(系)中渝茶4号的电子传递能力处于较高水平,其余品种(系)相对较弱。据报道,ETR与植物净光合速率呈显著相关,光合速率与ETR的动态变化一致[15]。这說明渝茶4号的光合能力相对较强。
2.5 不同物候期茶树品种的光启动速率差异
通过分析光响应曲线的斜率、最大电子传递速率和半饱和光强可进一步反应不同物候期茶树品种间光启动速率的差异。采用公式(1)(2)(3)对曲线的斜率(Alpha)、最大电子传递速率(ETRmax)和半饱和光强(Ik)进行测算。
计算结果列于表3。
结果显示,乌牛早和渝茶4号光响应曲线斜率最大且半饱和光强最小,说明这两个品种光化学启动较快,且在弱光照条件下就可以启动光化学过程,与这两个品种物候期较早的田间观察结果相吻合。随光照强度增加,乌牛早和渝茶4号通过PSII的电子传递速率迅速上升,直至达到最大电子传递速率时ETR增速减缓直至不再增加。其中ETRmax以福鼎和渝茶4号最大,乌牛早最小,这间接反映了乌牛早较其他参试品种(系)茶树虽能较快启动光化学反应,但也可能更易受到强光胁迫。
3 小结与讨论
叶绿素荧光技术作为植物光合生理的一种方便、快捷的无损检测技术,能准确地反映植物对光能的吸收、利用、分配和耗散过程,从而间接反映不同类型或不同环境下植物的光化学反应速率和最低光能需求的差异。通过分析和比较不同物候期类型茶树品种(系)的叶绿素荧光参数的数量特征及其种间差异可以看出,茶树对吸收的光能,总体上有79.4%~83.1%通过调节性或非调节性机制以热量形式耗散掉,20.6%~16.9%的能量用于光化学反应,但不同物候期类型的茶树的荧光参数存在较大差异,其中物候期较早的品种(系)光合中心活性较强,光响应曲线斜率较大,半饱和光强较小,在作用光光强一定时实际光合量子产量高于物候期较晚的品种(系),表现出早生品种(系)Y(II)、Alpha、ETRmax参数显著高于晚生品种(系),Fo参数显著低于晚生品种(系)的荧光特性,且与茶树物候期的先后具有较高一致性。进一步研究和利用这些特性与差异,对构建以叶绿素荧光技术为基础的茶树早生资源鉴定体系和推动茶树种质创新具有重要意义。
参考文献:
[1] Krause GH, Weis F. Chlorophyll fluorescence and photosynthesis: The Basics[J]. Annual Review of Plant Physiology, 1991(42): 313-349.
[2] 王碧莹,盛连喜,郭静,等.不同施氮处理下短梗五加与花楸叶绿素荧光参数日变化的比较研究[J].东北师大学报(自然科学版),2019,52(3):107-114.
[3] 李晓,冯伟,曾晓春,等.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006,26(10):2186-2196.
[4] Lamont BB, Lamont HC. Utilizable water in leaves of 8 arid species as derived from pressure-volume urves and chlorophyll fluorescence[J]. Physiol Plant, 2000, 110(1): 64-71.
[5] Lu Congming, Zhang Jianhua. Photosynthetic CO2 assimilation, chlorophyll fluorescence and photoinhibition as affected by nitrogen deficiency in maize plants[J]. Plant Science, 2000, 151(2): 135-143.
[6] Ogren E, Evans JR. Photoinhibitionin situin six species of Eucalyptus[J]. Australian Journal of Plant Physiology, 1992, 19: 223-232.
[7] 陈亮,杨亚军,虞富莲,等.茶树种质资源描述规范和数据标准[M].北京:中国农业出版社,2005.
[8] 许大全.光合作用效率[M].上海科学技术出版社,2002:32-33.
[9] 张红娜,苏钻贤,陈厚彬,等.荔枝成花期诱导性低温对其叶片光合特性的影响[J].广东农业科学,2016,10:59-63.
[10] 王兰兰,李琦,宋晓卉,等.环境条件对植物叶绿素荧光参数影响研究进展[J].沈阳师范大学学报(自然科学版),2019(4):362-367.
[11] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[12] 许大全,张玉忠,张荣铣.植物光合作用的光抑制[J].植物生理学通讯,1992,28(4):237-243.
[13] 陈景玲,王静,王谦,等.基于叶绿素荧光的荆条灌丛对栓皮栎幼苗庇荫效应研究[J].西北林学院学报,2014(4):46-53.
[14] 刘爱荣,陈双臣,王淼博,等.高温胁迫对番茄幼苗光合作用和叶绿素荧光参数的影响[J].西北农业学报,2010(5):145-148.
[15] 颜廷武.不同种源美国红枫苗期光合特性研究[J].辽宁林业科技,2014(6):24-26.
(责任编辑:丁志祥)
