海南岛不同林龄短枝木麻黄凋落物内外真菌多样性分析
2020-11-13左林芝张雅倩
左林芝,黄 蕊,张雅倩,陈 盼,李 蕾
海南师范大学生命科学学院/教育部热带岛屿生态学重点实验室,海口 571158
凋落物是森林系统中重要的组成部分,凋落物的降解是林地土壤养分的重要来源。大量文献已表明,凋落物表面附着的微生物及其内生菌则对凋落物的降解具有重要作用[1- 2]。另外,凋落物表面附着的微生物及其内生菌与植物次生产物代谢密切相关,林地土壤微生物受趋化作用在凋落物表面聚集[3],进而影响凋落物的降解及其产物合成,内生菌则直接以联合代谢的方式参与植物次生代谢产物的合成[4]。
海南岛海防林是由短枝木麻黄(CasuarinaequisetifoliaL.)单一树种构成的纯林,林下凋落物主要是木麻黄衰老死亡的枯枝。关于木麻黄凋落物的研究表明,木麻黄凋落物分解缓慢,对林地养分的贡献较小[5];而凋落物中的次生代谢产物则可通过淋溶等途径进入土壤,并在土壤中积累,通过化感作用,抑制了自身种子或其他植物种子的萌发和幼苗生长[6- 8]。
目前关于木麻黄微生物的研究主要集中于木麻黄林地土壤微生物、植株内生真菌的传统分离、鉴定及其化感作用等方面[9-12],对凋落物微生物的组成及其群落演替特征则较少涉及,而此研究对于探讨木麻黄林地微生物在凋落物的降解、化感物质积累等方面的生态功能具有重要作用。因此,本研究通过Illumina Miseq高通量测序法分析不同林龄木麻黄凋落物内外真菌的多样性变化,比较不同林龄木麻黄凋落物内外真菌群落结构的差异,为探讨微生物在木麻黄凋落物降解过程和次生产物代谢中的作用研究奠定基础。
1 材料与方法
1.1 研究地概况
以海南省海口市桂林洋开发区滨海的短枝木麻黄林为研究对象,该地(20.02°N,110.52°E)属热带海洋季风气候,日照时间长,热量丰富,年平均气温23.8℃。年降水量1500—2000 mm,平均相对湿度约85%,干、湿两季分明,5—10月为多雨季。常年以东风和东北风为主,年平均风速为3.4 m/s,6—10月为台风频发期[13]。
1.2 样品采集与预处理
2016年10月到研究样地的幼龄林(林龄5—8年)、中龄林(林龄15—20年)和成熟林(林龄30年及以上)中,以“S型取样法”选取15个样点,采集5—15 cm厚度的木麻黄凋落物及林地土壤。将样品充分混匀后装入无菌袋,用冰盒带回实验室并置于-80℃冰箱中保存。收集并混匀洗涤凋落物后的无菌水检测其外生真菌;用75%乙醇(1 min)、20% NaClO(10 min)和无菌水表面灭菌后检测凋落物内生真菌[14]。
1.3 凋落物内外真菌测序
取适量预处理后的样品,提取宏基因组DNA,采用真菌ITS区域作为目标DNA序列,以通用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA- 3′)和ITS2R(5′-GCTGCGTTCTTCATCGATGC- 3′)进行PCR扩增并纯化,进行荧光定量及Miseq文库构建后利用Illumina Miseq PE300平台测序,测序工作由上海美吉生物医药科技有限公司完成。
1.4 凋落物和土壤样品理化性质测定
将凋落物和土壤样品风干,研磨后过1mm的筛,混匀备用。参照《土壤理化分析实验指导》、《土壤分析技术规范》分析木麻黄凋落物和林地土壤理化性质[15-16]。采用烘干法测定样品含水率(WC)(GB 7833—1987);电位测定法测定pH值(土液比1∶2.5)(GB 7859—1987);凯氏定氮法测定全氮(TN)(GB 7173—1987);KCl浸提-比色法测定铵态氮(AN);酚二磺酸-比色法测定硝态氮(NN);NaOH熔融-钼锑抗比色法测定全磷(TP)(GB 7852—1987);环刀法测定容重(BD)(GB 7844—1987);重铬酸钾-消化炉加热法测定有机质(OM)(GB 7857—1987)。
1.5 数据处理与分析
根据overlap关系对Miseq测序得到的PE reads进行拼接,同时对序列质量进行质控和过滤,根据不同的相似度水平对所有序列进行OTU划分,对97%相似水平下的OTU进行生物信息统计分析。利用Mothur软件 (version v.1.30.1 http://www.mothur.org/wiki/Schloss_SOP#Alpha_diversity)计算凋落物内外真菌Alpha多样性(Shannon指数、Simpson指数和Chao1指数),分析真菌群落的多样性和丰富度[17- 19]。运用R程序和Mothur进行OTUs的维恩(Venn)分析,统计内外真菌共有和特有OTU数。基于tax_summary_a文件夹中的数据表,利用R语言工具作真菌群落柱形图,统计各样本在各分类水平上含有的真菌种类及比例。使用R v.2.15.2(统计计算的R基础2)和纯素包(2.0- 7版)生成真菌的分类分布热图。利用LEfSe软件(http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=lefse_upload)根据分类学组成进行线性判别分析(LDA),找出对样本差异具有显著影响的菌群。采用QIIME (Quantitative Insights to microecology)软件包1.9.1(Caporaso,J.G.,et al,2010)进行主坐标分析(PCoA)。使用软件Canoco for Windows 4.5作RDA/CCA图,分析凋落物真菌群落组成与环境因子的关系。
2 结果与分析
2.1 凋落物真菌群落多样性与组成分析
2.1.1凋落物内外真菌多样性
3林龄木麻黄凋落物的18个样品经测序得到669,476条有效序列,其中凋落物内生真菌分属于6门23纲56目108科177属255种,外生真菌分属于5门21纲55目114科198属293种。所有样品的文库覆盖率均在99.6%以上,说明测序条数足以覆盖凋落物样品中绝大部分微生物,能够反映绝大多数真菌信息(表1)。由Shannon指数、Simpson指数和Chao1指数看,木麻黄凋落物外生真菌的多样性和丰富度高于内生真菌。由3林龄的Chao1指数可看出,中龄林的丰富度高于幼龄林,成熟林最低。
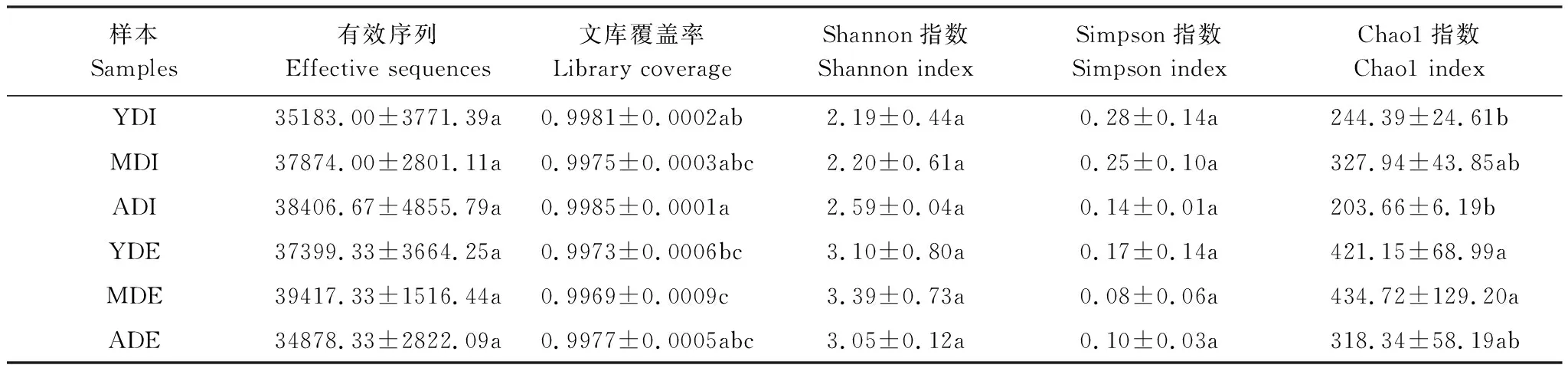
表1 不同林龄木麻黄凋落物内外真菌多样性指数表
YDI:幼龄林凋落物内生真菌,endophytic fungal communities of the litter in the young-aged forest;MDI:中龄林凋落物内生真菌,endophytic fungal communities of the litter in the middle-aged forest;ADI:成熟林凋落物内生真菌,endophytic fungal communities of the litter in the mature-aged forest;YDE:幼龄林凋落物外生真菌,epiphytic fungal communities of the litter in the young-aged forest;MDE:中龄林凋落物外生真菌,epiphytic fungal communities of the litter in the middle-aged forest;ADE:成熟林凋落物外生真菌,epiphytic fungal communities of the litter in the mature-aged forest;同一列中不同字母表示差异显著(P≤0.05)
2.1.2凋落物内外真菌的OTUs分析
对97%相似水平下的OUT进行Venn图分析发现(图1),木麻黄凋落物内生真菌OTUs总数为644,外生真菌为841,其中,555个OTUs为凋落物内、外真菌共有,分别占内、外真菌总数的86.18%、65.99%,表明大多内生真菌可能与外生真菌具有相同来源。从林龄上看,3林龄的共有OTUs数均远多于特有OTUs,表明凋落物内、外真菌群落差异由少数菌群决定,而特有的内生真菌则可能由种子来源[20-21]。
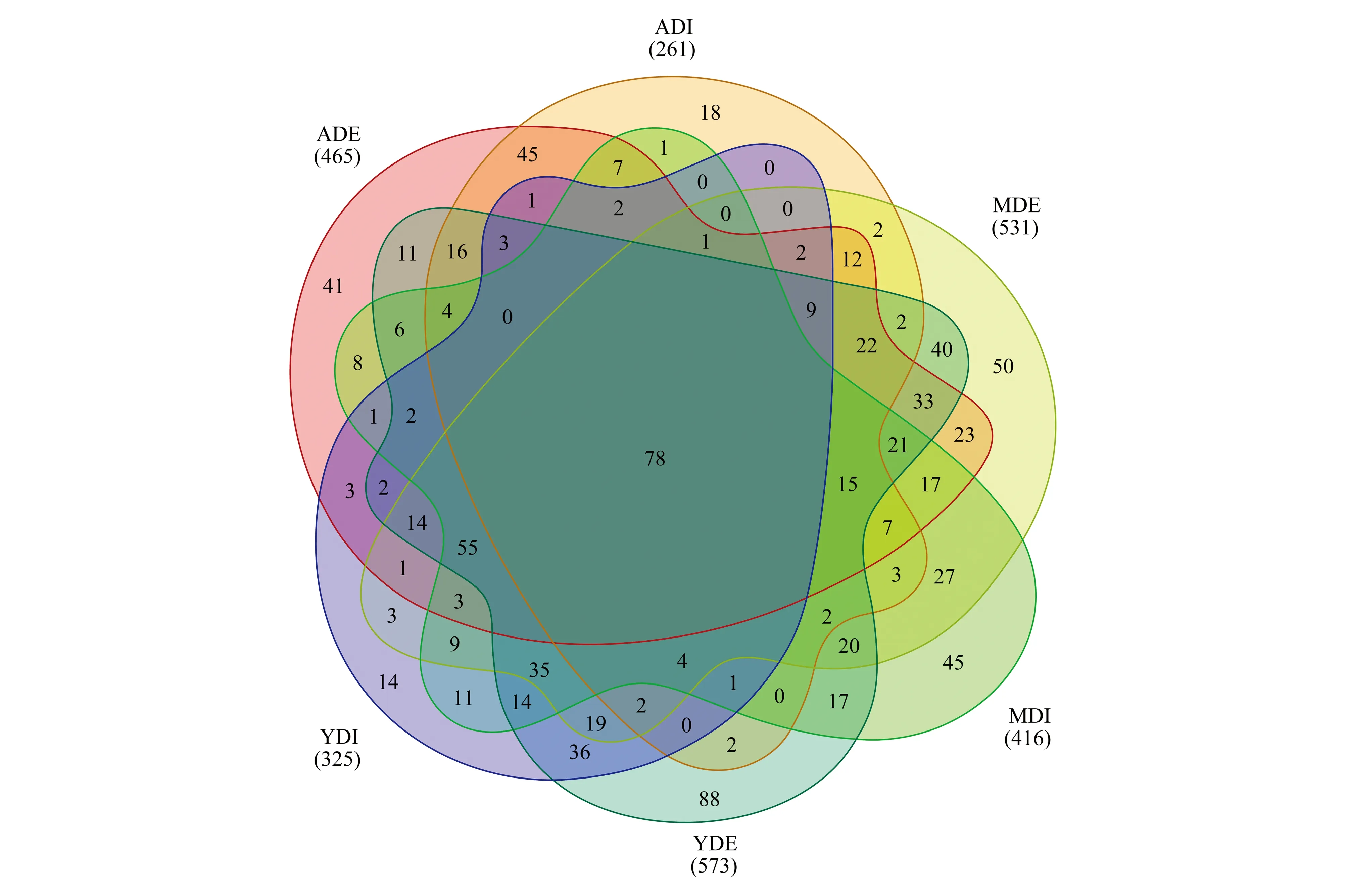
图1 不同林龄木麻黄凋落物内外真菌Venn图Fig.1 Venn diagram of endophytic and epiphytic fungi of C. equisetifolia litter at different ages YDI:幼龄林凋落物内生真菌,endophytic fungal communities of the litter in the young-aged forest;MDI:中龄林凋落物内生真菌,endophytic fungal communities of the litter in the middle-aged forest;ADI:成熟林凋落物内生真菌,endophytic fungal communities of the litter in the mature-aged forest;YDE:幼龄林凋落物外生真菌,epiphytic fungal communities of the litter in the young-aged forest;MDE:中龄林凋落物外生真菌,epiphytic fungal communities of the litter in the middle-aged forest;ADE:成熟林凋落物外生真菌,epiphytic fungal communities of the litter in the mature-aged forest
2.1.3凋落物内外真菌群落组成
不同林龄木麻黄凋落物真菌由6个门组成(图2),担子菌门(Basidiomycota)和子囊菌门(Ascomycota)在内生真菌中比例较高,分别占44.19%、43.78%;子囊菌门在外生真菌中比例最高(占74.63%),尤其在幼林龄外生真菌中占优势。随着林龄的增加,子囊菌门在内外真菌中所占比例呈先减后增的趋势,担子菌门则相反。
在目分类水平上,3林龄凋落物内生真菌共56目,外生真菌共55目,其中7个目为分类不明确目(图2)。煤炱目(Capnodiales)和炭角菌目(Xylariales)为内、外真菌共同优势目,糙孢孔目(Trechisporales)为内生真菌最优势目,格孢腔菌目(Pleosporales)为外生真菌最优势目。另外,随着林龄的增加,内生真菌群落均匀度先降低后升高,外生真菌则相反。
结果还表明,Rozellomycota为内生真菌特有菌门,且仅分布于中林龄凋落物内部。从目水平看,Coniochaetales、Magnaporthales、未分类壶菌门和未分类Rozellomycota等4个目始终定殖于凋落物内部,而蛙粪霉目(Basidiobolales)、丝孢酵母目(Trichosporonales)和未命名伞菌纲始终附生于凋落物外部。
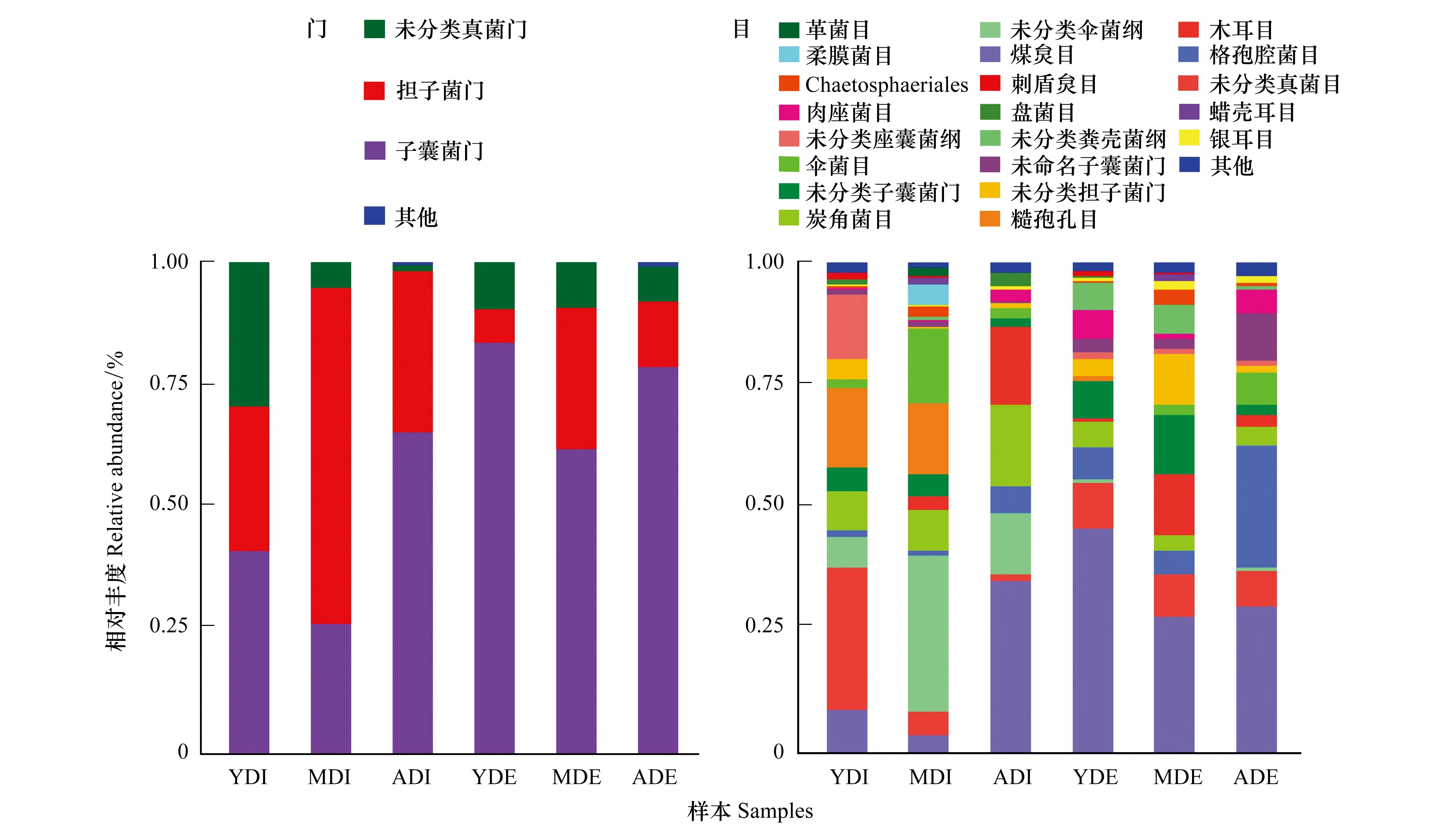
图2 不同林龄木麻黄凋落物内外真菌在不同分类水平中的群落结构Fig.2 Community structure of fungi of C. equisetifolia litter at different classification levels
2.1.4凋落物内外真菌PCoA分析
主坐标(PCoA)分析显示不同样本中97%相似性OTU的组成,反映木麻黄凋落物内外真菌群落组成的相似性和差异性。本研究中PC1、PC2是造成样品差异的两个最大差异性特征,累计解释了35.99%的真菌群落变化(图3)。整体上看,凋落物外生真菌群落分布较内生真菌集中,两者分布区域有部分重叠,表明内、外真菌群落结构存在一定共性。幼、中林龄真菌群落结构的分布区域有较大重叠,表明两者的真菌群落组成较相似。大多数成熟林样本在PC2轴上与幼、中林龄较远,表明该林龄真菌群落组成与其他林龄有较大差异。
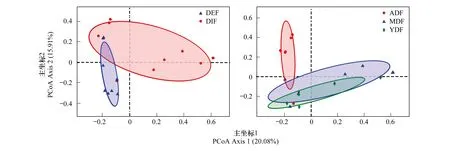
图3 不同林龄木麻黄凋落物内外真菌主坐标分析Fig.3 Principal co-ordinates analysis (PCoA) for endophytic and epiphytic fungal communities of the litter in differently aged C. equisetifolia forestsDEF:凋落物外生真菌,epiphytic fungal communities of the litter;DIF:凋落物内生真菌,endophytic fungal communities of the litter;ADF:成熟林凋落物真菌,fungal communities of the litter in the mature-aged forest;MDF:中龄林凋落物真菌,fungal communities of the litter in the middle-aged forest;YDF:幼龄林凋落物真菌,fungal communities of the litter in the young-aged forest
2.2 真菌群落差异分析
2.2.1凋落物内外真菌物种丰度分析
选取丰度排名前10的3林龄凋落物内外真菌目,对样本间的相对丰度进行聚类分析后得Heatmap图(图4)。糙孢孔目和未分类伞菌纲在内生真菌中的丰度显著高于外生真菌,煤炱目则相反。从林龄上看,未分类伞菌纲在中林龄凋落物内部显著富集,而外部丰度极低。糙孢孔目主要聚集在幼、中林龄凋落物内部。伞菌目(Agaricales)在幼、中林龄凋落物内部的丰度高于外部,成熟林则相反。相反地,木耳目(Auriculariales)在幼、中林龄凋落物内的丰度低于外部,成熟林相反。
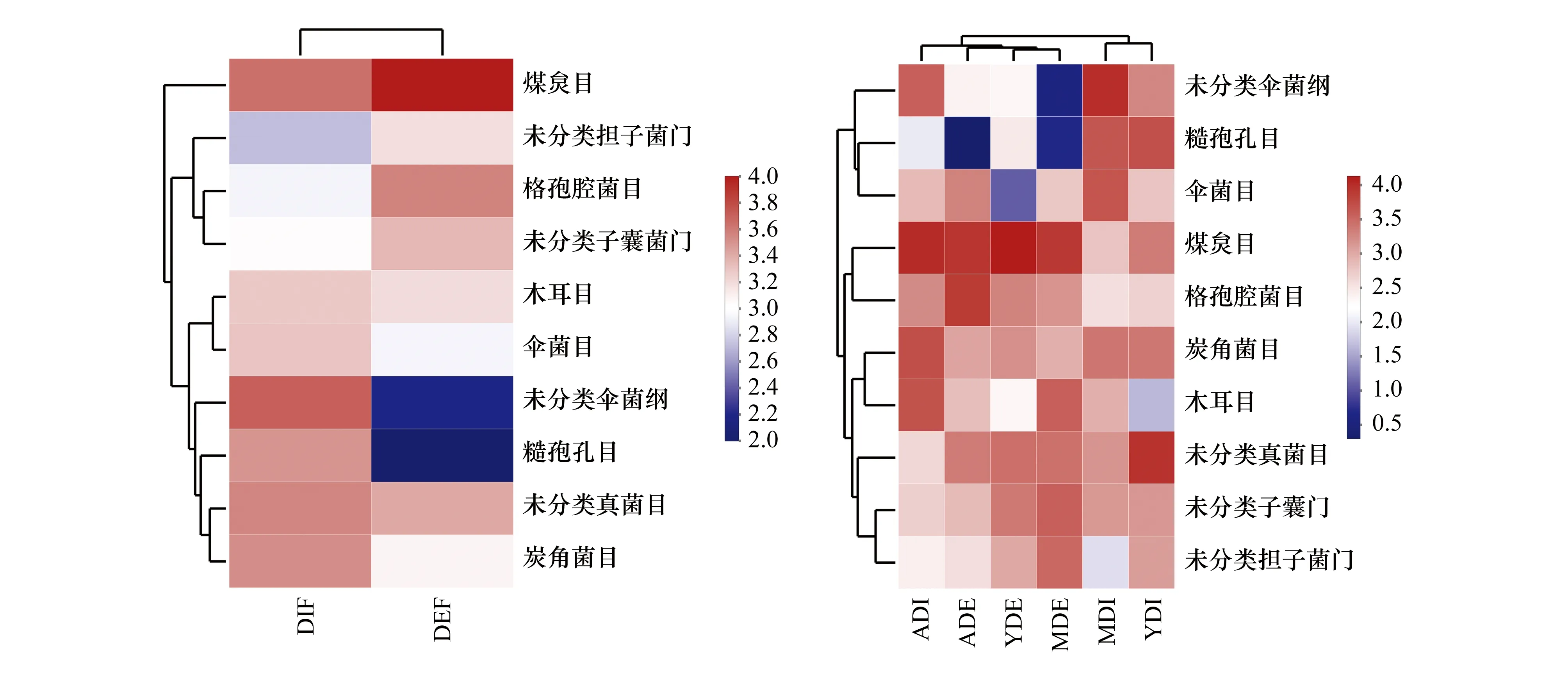
图4 不同林龄木麻黄凋落物真菌在目水平上的物种丰度热图Fig.4 Heat maps of species abundance of endophytic and epiphytic fungi in differently aged C. equisetifolia forests at the order level
2.2.2凋落物内外真菌LEfSe分析
通过LEfSe分析(LDA阈值为4)发现,对3林龄间差异有显著影响的内生真菌有:幼林龄糙孢孔目、中林龄低孔菌科(Hyponectriaceae)(从科到属)和成熟林肉杯菌科(Sarcoscyphaceae)(从科到属)。对3林龄间差异有显著影响的外生真菌有:幼林龄的丛赤壳属(Nectria)、球腔菌科(Mycosphaerellaceae)和成熟林的扁孔腔菌科(Lophiostomataceae)(从科到属)(图5)。由物种丰度分析发现,以上所述糙孢孔目和煤炱目的球腔菌科属于木麻黄凋落物真菌群落优势物种。

图5 不同林龄木麻黄凋落物内外真菌LEfSe分析Fig.5 LDA Effect Size analysis (LEfSe) of endophytic and epiphytic fungal communities of the litter in differently aged C. equisetifolia forests
2.3 凋落物内外真菌与凋落物、土壤理化性质的RDA/CCA分析
研究分析了木麻黄凋落物内外真菌群落结构与凋落物、林地土壤含水率(WC)、pH值、全氮(TN)、铵态氮(AN)、硝态氮(NN)、全磷(TP)、有机碳(OC)、容重(BD)、全钾(TK)、有机质(OM)等理化性质的相关性。结果表明,木麻黄凋落物内生真菌群落结构与凋落物pH值和有机碳有显著相关性,与土壤理化因子关系不显著;而外生真菌群落结构与土壤容重和铵态氮有显著相关性,与凋落物理化性质关系不显著(图6)。
在内外真菌共同的优势目中,煤炱目与凋落物pH、全磷、全氮及土壤含水率相关;炭角菌目与凋落物pH和土壤全磷相关性较大;木耳目与凋落物pH、土壤铵态氮和容重相关性较大。伞菌目、糙孢孔目等内生真菌特有优势目中,前者与凋落物全磷相关性较大,后者与凋落物含水率相关性较大。格孢腔菌目、肉座菌目(Hypocreales)等外生真菌优势目中,格孢腔菌目与土壤pH相关性较大,肉座菌目与凋落物全氮相关性较大(图7)。
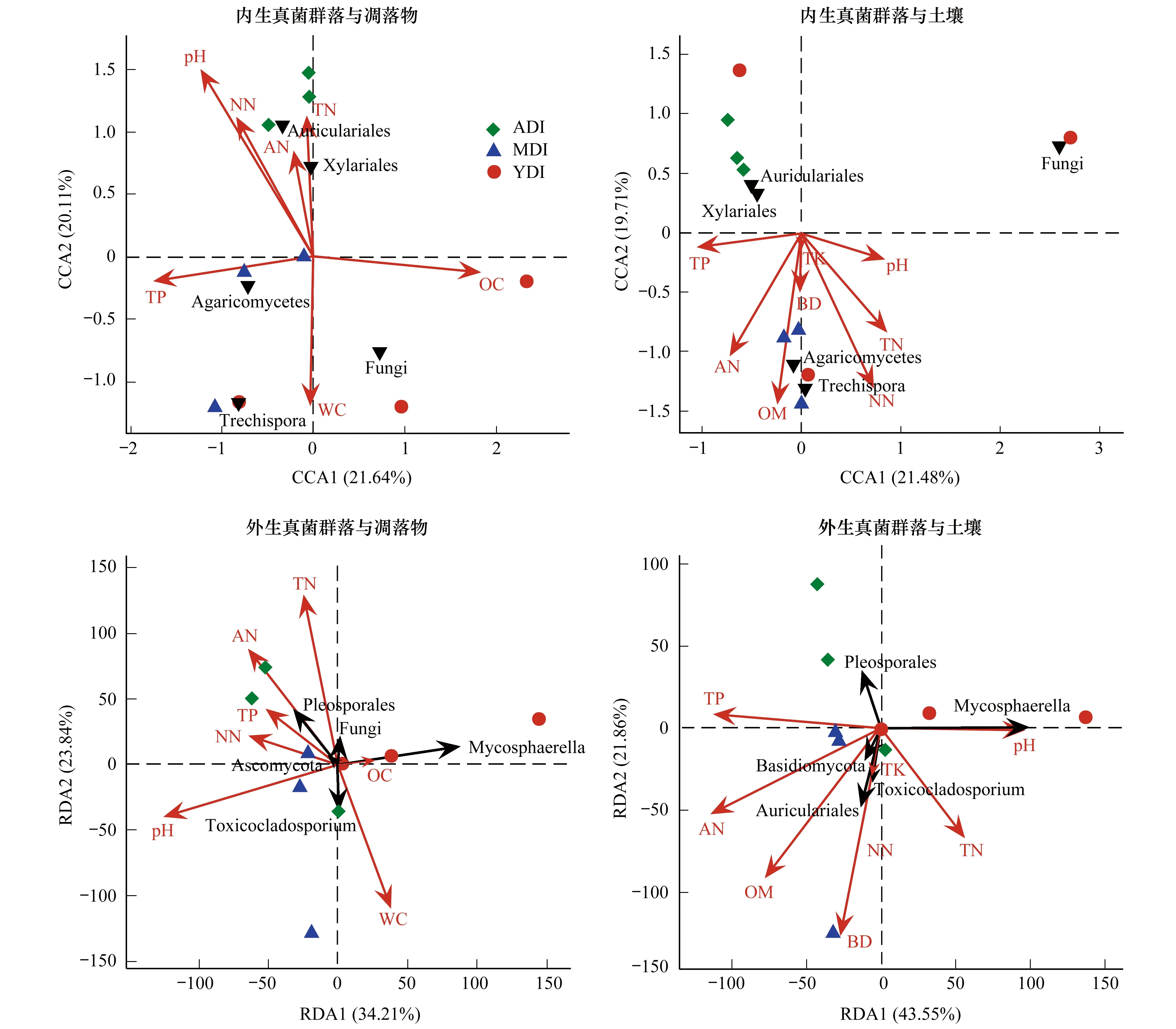
图6 真菌群落与凋落物及土壤理化性质的相关性分析Fig.6 RedundancyAnalysis (RDA)/Canonical Correspondence Analysis (CCA) between fungal communities and the physicochemical property of the forest litter and soil pH:pH值;AN:铵态氮;NN:硝态氮;TN:全氮;TP:全磷;WC:含水率; OC:有机碳;BD:容重;TK:全钾;OM:有机质。黑色倒三角和黑色箭头表示高丰度类群
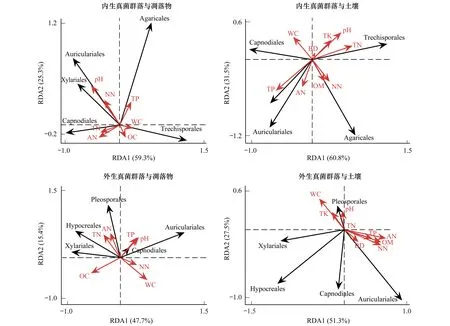
图7 优势真菌群落与凋落物及土壤理化性质的RDA分析Fig.7 RDA analysis between dominant fungal communities and the physicochemical property of the forest litter and soil
3 结论与讨论
3.1 凋落物内外真菌群落组成
凋落物真菌多样性分析表明,不同林龄木麻黄凋落物外生真菌多样性和丰富度高于内生真菌,这与Gang等的研究结果“植物根际真菌群落的多样性高于根内生真菌”相似[22]。凋落物内、外真菌共有物种数目占较大比例,说明木麻黄凋落物内、外真菌群落组成有较大共性,说明外生真菌群落来源部分由于菌群的趋化作用而聚集外[3],大部分可能来源于凋落物的降解。分析结果显示,蛙粪霉目等3个目始终附生于凋落物外部,Coniochaetales 等4个目始终定殖于凋落物内部,这些内生真菌特有种可能来源于木麻黄种子或空气环境[20-21,23],外生真菌特有种则可能是在凋落物降解过程中林地土壤真菌不断向其转移的结果。
不同林龄间,凋落物内外真菌群落组成大多与木麻黄林龄无显著相关性,可能表明木麻黄枝叶作为凋落物脱离母体后,植株对真菌的影响逐渐减弱,而受所处环境的影响逐渐增加。蓝丽英等和徐璇等的研究表明,在凋落物分解过程中,土壤动物以选择性牧食作用来调控微生物种群数量,其排泄物还可刺激微生物生长[24-25]。
3.2 凋落物内外真菌优势物种
分析3林龄木麻黄凋落物内外真菌群落的相似性和差异性发现,子囊菌门和担子菌门为最优势门,在目水平上,煤炱目、炭角菌目和木耳目为凋落物内、外真菌共同优势目。据文献报道,子囊菌门和担子菌门在兰科[26]、铁皮石斛[27]和杨树[28]等多种植物中为主导菌门,煤炱目、炭角菌目是植物中较为普遍的内生真菌[29-30],与我们的研究结果一致。
前人研究还显示,子囊菌门大多菌群可分解新鲜植物残体[31],我们研究中,子囊菌门的格孢腔菌目、肉座菌目等为凋落物外生真菌的优势菌群,表明这些外生真菌优势物种可能在凋落物降解过程中起重要作用。
3.3 凋落物内外真菌群落与环境因子的关系
RDA/CCA分析结果表明,凋落物内生真菌群落结构受凋落物pH值和有机碳显著影响,而凋落物外生真菌群落结构则与土壤容重和铵态氮显著相关。说明宿主植物养分是内生真菌群落组成的主要决定因素,而附着于凋落物表面真菌的生长则主要依赖土壤提供的养分。
前人的研究也表明,pH和有机质对真菌群落组成具有重要影响[32-33],另外,总磷的增加导致真菌群落多样性降低[34]。本研究发现与幼、中林龄相比,成熟林土壤全磷含量最高,而该林龄凋落物内外真菌多样性及丰富度均最低,表明较高的磷含量不利于真菌群落的生长。另外,中林龄凋落物真菌群落较幼、成林高,该林土壤有机质、硝态氮、铵态氮较幼、成林高,课题组前期的研究表明,中林土壤微生态环境适合植株生长[35],真菌群落在其中发挥着重要作用。
